911~ ...
Molecular identification of bacteria and eukarya inhabiting an Antarctic cryoconite hole
2
2003
... 冰尘洞广泛存在于全球的冰川消融区[17], 其被认为是冰冻圈中微生物活动的热点[5].冰尘是散落在冰川表面由矿物质、 有机质和微生物组成的聚合体, 既包括远源输送来的细粉尘和气溶胶组份, 也有局地源的粗冰碛物及来自周围生态系统的土壤和植被碎屑等.冰尘附着在冰川表面, 通过降低反照率和增加局部融化形成的圆形孔洞被称作冰尘洞[18-19](图1), 其通常几十厘米宽和深, 底部含有一层沉积物, 其内水温相对恒定在0.1 ℃[20-22].冰尘洞广泛分布于全球冰川中, 可覆盖1%~10%的冰川表面.冰尘洞生态系统是生命存在的极端环境[15,20], 但是其内拥有丰富的微生物群落, 被病毒、 原核生物和真核生物等多种微生物定殖[13,23], 进行着活跃的生物地球化学循环[19]. ...
... 1983年, Azam首次提出了微生物食物环的概念, 微生物食物环是指异养细菌能够吸收利用生产者生产、 消费者摄食及其他过程中产生的溶解有机物(DOM: Dissolve organic matter), 将其部分转化为自身的颗粒有机物(POM: Particulate organic matter), 并且随着细菌被原生动物摄食, 这些POM又重新回到主食物链中不断循环利用的过程[60].而病毒对于微生物食物环具有重要意义, 使其得到进一步丰富.病毒能够通过裂解宿主, 使得宿主体内的DOM释放出来, 再次被未被病毒感染的异养细菌吸收利用, 从而使得大部分物质和能量在微生物食物环中再循环, 最终被呼吸消耗掉, 产生无机营养物质.这一氧化有机物质、 再生无机营养的环被称为病毒回路[43,61].因而病毒能够将微生物食物环缩短, 催化营养物质从生物体转移到 DOM 和 POM, 将其引导至食物链[5,23], 减少能量向更高营养级的运输[62]. ...
The biodiversity and biogeochemistry of cryoconite holes from McMurdo Dry Valley Glaciers, Antarctica
2
2004
... 在温带冰川地区, 夏季冰川消融时冰尘洞直接暴露于空气中, 并与周围融化的.表面以及气体进行水与大气的交换, 但是在寒冷的时节以及极度寒冷的地点, 冰尘洞会被雪或者冰覆盖, 将冰尘洞与大气及周围表面隔离开来[24].气温和太阳辐射决定了冰尘洞的大小和生长速度[25].而影响冰尘洞形成的另一个重要因素则是这些独特栖息地的微生物组合.藻类、 细菌以及初级生产者的活动会造成深色有机物积累, 增大冰雪的吸光性, 导致冰反照率降低[26-27], 从而加剧冰川的融化与退缩[28]. ...
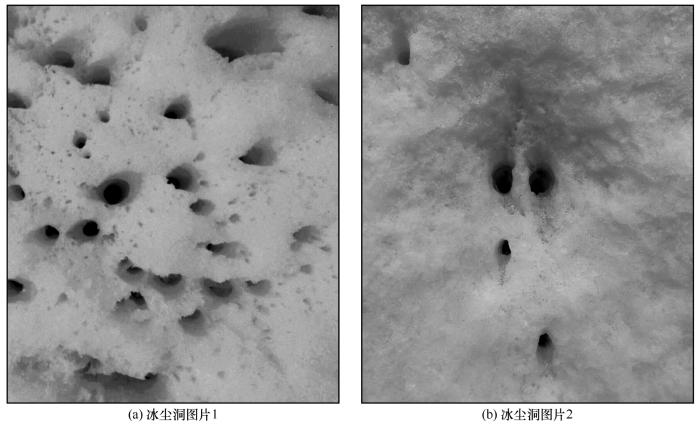
... 冰尘洞中的大部分微生物生物质, 例如藻类、 原核光合自养生物和异养生物, 以及病毒等都存在于冰尘颗粒中[27].这些微生物活性很高, 在冰尘穴中具有极高的活性与代谢活动, 并通过一系列的代谢活动参与碳、 氮、 磷等营养物质循环, 进而促进冰川系统内有机物质形成[6,24,29](图2).Anesio[27], Edwards等[30]在测量Svalbard群岛, Greenland和Alps山冰川冰尘洞光合作用和呼吸速率的基础上, 估计出冰尘洞中的净初级生产量可能能够达到每年64 000 t碳, 极大的挑战了之前对于冰川生态系统是异养生态系统的认知. ...
Cryoconite hole thermodynamics
1
... 在温带冰川地区, 夏季冰川消融时冰尘洞直接暴露于空气中, 并与周围融化的.表面以及气体进行水与大气的交换, 但是在寒冷的时节以及极度寒冷的地点, 冰尘洞会被雪或者冰覆盖, 将冰尘洞与大气及周围表面隔离开来[24].气温和太阳辐射决定了冰尘洞的大小和生长速度[25].而影响冰尘洞形成的另一个重要因素则是这些独特栖息地的微生物组合.藻类、 细菌以及初级生产者的活动会造成深色有机物积累, 增大冰雪的吸光性, 导致冰反照率降低[26-27], 从而加剧冰川的融化与退缩[28]. ...
The microorganisms of cryoconite holes (algae, Archaea, bacteria, cyanobacteria, fungi, and Protista): a review
1
2016
... 在温带冰川地区, 夏季冰川消融时冰尘洞直接暴露于空气中, 并与周围融化的.表面以及气体进行水与大气的交换, 但是在寒冷的时节以及极度寒冷的地点, 冰尘洞会被雪或者冰覆盖, 将冰尘洞与大气及周围表面隔离开来[24].气温和太阳辐射决定了冰尘洞的大小和生长速度[25].而影响冰尘洞形成的另一个重要因素则是这些独特栖息地的微生物组合.藻类、 细菌以及初级生产者的活动会造成深色有机物积累, 增大冰雪的吸光性, 导致冰反照率降低[26-27], 从而加剧冰川的融化与退缩[28]. ...
High microbial activity on glaciers: importance to the global carbon cycle
3
2010
... 在温带冰川地区, 夏季冰川消融时冰尘洞直接暴露于空气中, 并与周围融化的.表面以及气体进行水与大气的交换, 但是在寒冷的时节以及极度寒冷的地点, 冰尘洞会被雪或者冰覆盖, 将冰尘洞与大气及周围表面隔离开来[24].气温和太阳辐射决定了冰尘洞的大小和生长速度[25].而影响冰尘洞形成的另一个重要因素则是这些独特栖息地的微生物组合.藻类、 细菌以及初级生产者的活动会造成深色有机物积累, 增大冰雪的吸光性, 导致冰反照率降低[26-27], 从而加剧冰川的融化与退缩[28]. ...
... 冰尘洞中的大部分微生物生物质, 例如藻类、 原核光合自养生物和异养生物, 以及病毒等都存在于冰尘颗粒中[27].这些微生物活性很高, 在冰尘穴中具有极高的活性与代谢活动, 并通过一系列的代谢活动参与碳、 氮、 磷等营养物质循环, 进而促进冰川系统内有机物质形成[6,24,29](图2).Anesio[27], Edwards等[30]在测量Svalbard群岛, Greenland和Alps山冰川冰尘洞光合作用和呼吸速率的基础上, 估计出冰尘洞中的净初级生产量可能能够达到每年64 000 t碳, 极大的挑战了之前对于冰川生态系统是异养生态系统的认知. ...
... [27], Edwards等[30]在测量Svalbard群岛, Greenland和Alps山冰川冰尘洞光合作用和呼吸速率的基础上, 估计出冰尘洞中的净初级生产量可能能够达到每年64 000 t碳, 极大的挑战了之前对于冰川生态系统是异养生态系统的认知. ...
Biogeochemically diverse organic matter in Alpine glaciers and its downstream fate
1
2012
... 在温带冰川地区, 夏季冰川消融时冰尘洞直接暴露于空气中, 并与周围融化的.表面以及气体进行水与大气的交换, 但是在寒冷的时节以及极度寒冷的地点, 冰尘洞会被雪或者冰覆盖, 将冰尘洞与大气及周围表面隔离开来[24].气温和太阳辐射决定了冰尘洞的大小和生长速度[25].而影响冰尘洞形成的另一个重要因素则是这些独特栖息地的微生物组合.藻类、 细菌以及初级生产者的活动会造成深色有机物积累, 增大冰雪的吸光性, 导致冰反照率降低[26-27], 从而加剧冰川的融化与退缩[28]. ...
Biogeography of cryoconite forming cyanobacteria on polar and Asian glaciers
1
2017
... 冰尘洞中的大部分微生物生物质, 例如藻类、 原核光合自养生物和异养生物, 以及病毒等都存在于冰尘颗粒中[27].这些微生物活性很高, 在冰尘穴中具有极高的活性与代谢活动, 并通过一系列的代谢活动参与碳、 氮、 磷等营养物质循环, 进而促进冰川系统内有机物质形成[6,24,29](图2).Anesio[27], Edwards等[30]在测量Svalbard群岛, Greenland和Alps山冰川冰尘洞光合作用和呼吸速率的基础上, 估计出冰尘洞中的净初级生产量可能能够达到每年64 000 t碳, 极大的挑战了之前对于冰川生态系统是异养生态系统的认知. ...
Possible interactions between bacterial diversity, microbial activity and supraglacial hydrology of cryoconite holes in Svalbard
1
2010
... 冰尘洞中的大部分微生物生物质, 例如藻类、 原核光合自养生物和异养生物, 以及病毒等都存在于冰尘颗粒中[27].这些微生物活性很高, 在冰尘穴中具有极高的活性与代谢活动, 并通过一系列的代谢活动参与碳、 氮、 磷等营养物质循环, 进而促进冰川系统内有机物质形成[6,24,29](图2).Anesio[27], Edwards等[30]在测量Svalbard群岛, Greenland和Alps山冰川冰尘洞光合作用和呼吸速率的基础上, 估计出冰尘洞中的净初级生产量可能能够达到每年64 000 t碳, 极大的挑战了之前对于冰川生态系统是异养生态系统的认知. ...
Virioplankton: viruses in aquatic ecosystems
5
2000
... 目前关于冰尘洞内病毒丰度的相关研究较少, 仅有的少量研究集中于北极[13,21-22](表1).北极冰尘洞内病毒丰度跨越5个数量级, 且冰尘洞不同组分内病毒丰度差别较大.一般而言, 底层沉积物的病毒丰度要高于水或者冰中的病毒丰度, 其范围为8.79×106~2.62×109 VLP·g-1(VLP: Virus like particles, 病毒类似颗粒).例如, Bellas等[21]对北极Svalbard地区的两个山谷冰川以及Greenland冰盖消融区冰尘洞沉积物内的病毒丰度进行了研究, 结果表明上述地区冰尘洞中病毒丰度达到5.62~24.5×108 VLP·g-1, 平均值为(14.8±7.3)×108 VLP·g-1.水中和雪中病毒丰度较少, 一般在105 VLP·mL-1左右.Anesio等发现[13], 北极Midre Lovénbreen和Austre Brøggerbreen两个山谷冰川冰尘洞水中的病毒丰度在2.4~11.9×105 VLP·mL-1之间, 而冰中病毒丰度只有1~56×104 VLP·mL-1.Säwström等[22]的研究结果表示, Midre Lovénbreen冰尘洞水中病毒丰度为3.97~7.04×104 VLP·mL-1.全球海洋和湖泊中病毒丰度的相关研究表明, 湖泊中浮游病毒丰度一般在105~108 VLP·mL-1之间[31-32], 而海洋中病毒丰度一般为106~107 VLP·mL-1[10].由此可知, 与温带地区淡水生态系统相比, 冰和水中的病毒丰度通常低2个数量级[33]. ...
... 此外, 从表1, 可以看出, 北极冰尘洞中病毒丰度与细菌丰度比值(VBR: Virus to bacteria ratio)变化范围较大, 为0.3~74.一般而言, 较高的VBR表明病毒对细菌群落的控制作用越强[13].比如: 南极洲盐湖的VBR在30.5~126.7之间[34], 极地寡营养湖泊中VBR在1.15~96.7[14,35-36].一般情况下, 营养状况和生产力越大, VBR越大[31,37], 所以极地地区VBR会略低于低纬度地区.冰尘洞中较低的 VBR 并不一定意味着病毒对细菌群落动态的控制作用低[13], Mei和Danovaro认为[38], 尽管VBR较低, 但病毒仍占地中海沉积物中细菌死亡率的40%以上.Danovaro研究表明[39], 沉积物中较低的VBR可能是由腐殖质和富含外酶的复杂有机基质造成的, 它们的存在会引起病毒在感染宿主并被释放之后迅速衰变, 进而导致VBR较低. ...
... 病毒是一种寄生生物, 必须通过侵染特定的宿主, 借助于宿主的营养物质及遗传信息才能够完成自己的生命周期.根据病毒对宿主细胞裂解能力的大小, 可以将病毒的增殖方式分为三种基本感染周期类型: 裂解性感染、 溶源性感染和假溶源性感染[42].裂解性感染能够在短时间内引起宿主细胞的裂解, 进而释放出大量的子代病毒, 因而是病毒繁殖的主要方式[43-44].然而, 溶源性感染通常被看作是病毒用于应对增殖不利条件的一种生活策略[9,31,45].该策略在低温生境尤其是超寡营养湖泊和冰川中发挥着比在其他生境中更大的作用[9].溶源循环或裂解循环是否发生取决于宿主细胞和环境条件, 如水体的营养状态[46].Laybourn-Parry[47]和Säwström等[48]分别在南极湖泊开展了研究, 结果均表明南极湖泊中冬季和春季溶源性发生频率较高, 而夏季则未发现溶源性.Säwström等[49]对北极湖泊的研究结果也表明夏季北极淡水环境中没有溶源性.Bellas等[21]对北极冰尘洞内的沉积物进行了研究, 同样在夏季冰尘洞沉积物中未检测到溶源性.以上结果均表明溶源性感染策略在低温等不利条件下发挥更大的作用.假溶源性是指表面上类似于溶源性, 但是病毒在宿主体内不会立即表达复制, 而是暂时处于休眠状态.该策略被认为能够使病毒在宿主丰度较低且营养限制的情况下在细菌群体中维持自身[31].Säwström等[49]研究发现, 北极和南极洲的超寡营养低温生境中受病毒感染的细胞大多具有非常低的爆发大小.因而在营养限制的条件下, 上述假溶源性策略可能会帮助病毒快速地对外界环境的变化做出反应[31].溶源性策略和假溶源性策略能够帮助部分病毒在极端条件下延长寿命, 从而维持更多的病毒数量和保证多样性[9,50]. ...
... [31].Säwström等[49]研究发现, 北极和南极洲的超寡营养低温生境中受病毒感染的细胞大多具有非常低的爆发大小.因而在营养限制的条件下, 上述假溶源性策略可能会帮助病毒快速地对外界环境的变化做出反应[31].溶源性策略和假溶源性策略能够帮助部分病毒在极端条件下延长寿命, 从而维持更多的病毒数量和保证多样性[9,50]. ...
... [31].溶源性策略和假溶源性策略能够帮助部分病毒在极端条件下延长寿命, 从而维持更多的病毒数量和保证多样性[9,50]. ...
Current insights into phage biodiversity and biogeography
1
2009
... 目前关于冰尘洞内病毒丰度的相关研究较少, 仅有的少量研究集中于北极[13,21-22](表1).北极冰尘洞内病毒丰度跨越5个数量级, 且冰尘洞不同组分内病毒丰度差别较大.一般而言, 底层沉积物的病毒丰度要高于水或者冰中的病毒丰度, 其范围为8.79×106~2.62×109 VLP·g-1(VLP: Virus like particles, 病毒类似颗粒).例如, Bellas等[21]对北极Svalbard地区的两个山谷冰川以及Greenland冰盖消融区冰尘洞沉积物内的病毒丰度进行了研究, 结果表明上述地区冰尘洞中病毒丰度达到5.62~24.5×108 VLP·g-1, 平均值为(14.8±7.3)×108 VLP·g-1.水中和雪中病毒丰度较少, 一般在105 VLP·mL-1左右.Anesio等发现[13], 北极Midre Lovénbreen和Austre Brøggerbreen两个山谷冰川冰尘洞水中的病毒丰度在2.4~11.9×105 VLP·mL-1之间, 而冰中病毒丰度只有1~56×104 VLP·mL-1.Säwström等[22]的研究结果表示, Midre Lovénbreen冰尘洞水中病毒丰度为3.97~7.04×104 VLP·mL-1.全球海洋和湖泊中病毒丰度的相关研究表明, 湖泊中浮游病毒丰度一般在105~108 VLP·mL-1之间[31-32], 而海洋中病毒丰度一般为106~107 VLP·mL-1[10].由此可知, 与温带地区淡水生态系统相比, 冰和水中的病毒丰度通常低2个数量级[33]. ...
Influence of humic substances on bacterial and viral dynamics in freshwaters
1
2004
... 目前关于冰尘洞内病毒丰度的相关研究较少, 仅有的少量研究集中于北极[13,21-22](表1).北极冰尘洞内病毒丰度跨越5个数量级, 且冰尘洞不同组分内病毒丰度差别较大.一般而言, 底层沉积物的病毒丰度要高于水或者冰中的病毒丰度, 其范围为8.79×106~2.62×109 VLP·g-1(VLP: Virus like particles, 病毒类似颗粒).例如, Bellas等[21]对北极Svalbard地区的两个山谷冰川以及Greenland冰盖消融区冰尘洞沉积物内的病毒丰度进行了研究, 结果表明上述地区冰尘洞中病毒丰度达到5.62~24.5×108 VLP·g-1, 平均值为(14.8±7.3)×108 VLP·g-1.水中和雪中病毒丰度较少, 一般在105 VLP·mL-1左右.Anesio等发现[13], 北极Midre Lovénbreen和Austre Brøggerbreen两个山谷冰川冰尘洞水中的病毒丰度在2.4~11.9×105 VLP·mL-1之间, 而冰中病毒丰度只有1~56×104 VLP·mL-1.Säwström等[22]的研究结果表示, Midre Lovénbreen冰尘洞水中病毒丰度为3.97~7.04×104 VLP·mL-1.全球海洋和湖泊中病毒丰度的相关研究表明, 湖泊中浮游病毒丰度一般在105~108 VLP·mL-1之间[31-32], 而海洋中病毒丰度一般为106~107 VLP·mL-1[10].由此可知, 与温带地区淡水生态系统相比, 冰和水中的病毒丰度通常低2个数量级[33]. ...
Virus and microbial loop dynamics over an annual cycle in three contrasting Antarctic lakes
1
2005
... 此外, 从表1, 可以看出, 北极冰尘洞中病毒丰度与细菌丰度比值(VBR: Virus to bacteria ratio)变化范围较大, 为0.3~74.一般而言, 较高的VBR表明病毒对细菌群落的控制作用越强[13].比如: 南极洲盐湖的VBR在30.5~126.7之间[34], 极地寡营养湖泊中VBR在1.15~96.7[14,35-36].一般情况下, 营养状况和生产力越大, VBR越大[31,37], 所以极地地区VBR会略低于低纬度地区.冰尘洞中较低的 VBR 并不一定意味着病毒对细菌群落动态的控制作用低[13], Mei和Danovaro认为[38], 尽管VBR较低, 但病毒仍占地中海沉积物中细菌死亡率的40%以上.Danovaro研究表明[39], 沉积物中较低的VBR可能是由腐殖质和富含外酶的复杂有机基质造成的, 它们的存在会引起病毒在感染宿主并被释放之后迅速衰变, 进而导致VBR较低. ...
Carbon dynamics in an ultra-oligotrophic epishelf lake (Beaver Lake, Antarctica) in summer
1
2010
... 此外, 从表1, 可以看出, 北极冰尘洞中病毒丰度与细菌丰度比值(VBR: Virus to bacteria ratio)变化范围较大, 为0.3~74.一般而言, 较高的VBR表明病毒对细菌群落的控制作用越强[13].比如: 南极洲盐湖的VBR在30.5~126.7之间[34], 极地寡营养湖泊中VBR在1.15~96.7[14,35-36].一般情况下, 营养状况和生产力越大, VBR越大[31,37], 所以极地地区VBR会略低于低纬度地区.冰尘洞中较低的 VBR 并不一定意味着病毒对细菌群落动态的控制作用低[13], Mei和Danovaro认为[38], 尽管VBR较低, 但病毒仍占地中海沉积物中细菌死亡率的40%以上.Danovaro研究表明[39], 沉积物中较低的VBR可能是由腐殖质和富含外酶的复杂有机基质造成的, 它们的存在会引起病毒在感染宿主并被释放之后迅速衰变, 进而导致VBR较低. ...
Spatial and temporal distribution of virioplankton in arctic-alpine lake
1
2018
... 此外, 从表1, 可以看出, 北极冰尘洞中病毒丰度与细菌丰度比值(VBR: Virus to bacteria ratio)变化范围较大, 为0.3~74.一般而言, 较高的VBR表明病毒对细菌群落的控制作用越强[13].比如: 南极洲盐湖的VBR在30.5~126.7之间[34], 极地寡营养湖泊中VBR在1.15~96.7[14,35-36].一般情况下, 营养状况和生产力越大, VBR越大[31,37], 所以极地地区VBR会略低于低纬度地区.冰尘洞中较低的 VBR 并不一定意味着病毒对细菌群落动态的控制作用低[13], Mei和Danovaro认为[38], 尽管VBR较低, 但病毒仍占地中海沉积物中细菌死亡率的40%以上.Danovaro研究表明[39], 沉积物中较低的VBR可能是由腐殖质和富含外酶的复杂有机基质造成的, 它们的存在会引起病毒在感染宿主并被释放之后迅速衰变, 进而导致VBR较低. ...
高寒地区湖泊浮游病毒时空分布特征
1
2018
... 此外, 从表1, 可以看出, 北极冰尘洞中病毒丰度与细菌丰度比值(VBR: Virus to bacteria ratio)变化范围较大, 为0.3~74.一般而言, 较高的VBR表明病毒对细菌群落的控制作用越强[13].比如: 南极洲盐湖的VBR在30.5~126.7之间[34], 极地寡营养湖泊中VBR在1.15~96.7[14,35-36].一般情况下, 营养状况和生产力越大, VBR越大[31,37], 所以极地地区VBR会略低于低纬度地区.冰尘洞中较低的 VBR 并不一定意味着病毒对细菌群落动态的控制作用低[13], Mei和Danovaro认为[38], 尽管VBR较低, 但病毒仍占地中海沉积物中细菌死亡率的40%以上.Danovaro研究表明[39], 沉积物中较低的VBR可能是由腐殖质和富含外酶的复杂有机基质造成的, 它们的存在会引起病毒在感染宿主并被释放之后迅速衰变, 进而导致VBR较低. ...
Temporal variation in freshwater viral and bacterial community composition
1
2008
... 此外, 从表1, 可以看出, 北极冰尘洞中病毒丰度与细菌丰度比值(VBR: Virus to bacteria ratio)变化范围较大, 为0.3~74.一般而言, 较高的VBR表明病毒对细菌群落的控制作用越强[13].比如: 南极洲盐湖的VBR在30.5~126.7之间[34], 极地寡营养湖泊中VBR在1.15~96.7[14,35-36].一般情况下, 营养状况和生产力越大, VBR越大[31,37], 所以极地地区VBR会略低于低纬度地区.冰尘洞中较低的 VBR 并不一定意味着病毒对细菌群落动态的控制作用低[13], Mei和Danovaro认为[38], 尽管VBR较低, 但病毒仍占地中海沉积物中细菌死亡率的40%以上.Danovaro研究表明[39], 沉积物中较低的VBR可能是由腐殖质和富含外酶的复杂有机基质造成的, 它们的存在会引起病毒在感染宿主并被释放之后迅速衰变, 进而导致VBR较低. ...
Virus production and life strategies in aquatic sediments
1
2004
... 此外, 从表1, 可以看出, 北极冰尘洞中病毒丰度与细菌丰度比值(VBR: Virus to bacteria ratio)变化范围较大, 为0.3~74.一般而言, 较高的VBR表明病毒对细菌群落的控制作用越强[13].比如: 南极洲盐湖的VBR在30.5~126.7之间[34], 极地寡营养湖泊中VBR在1.15~96.7[14,35-36].一般情况下, 营养状况和生产力越大, VBR越大[31,37], 所以极地地区VBR会略低于低纬度地区.冰尘洞中较低的 VBR 并不一定意味着病毒对细菌群落动态的控制作用低[13], Mei和Danovaro认为[38], 尽管VBR较低, 但病毒仍占地中海沉积物中细菌死亡率的40%以上.Danovaro研究表明[39], 沉积物中较低的VBR可能是由腐殖质和富含外酶的复杂有机基质造成的, 它们的存在会引起病毒在感染宿主并被释放之后迅速衰变, 进而导致VBR较低. ...
Higher abundance of bacteria than of viruses in deep Mediterranean sediments
1
2002
... 此外, 从表1, 可以看出, 北极冰尘洞中病毒丰度与细菌丰度比值(VBR: Virus to bacteria ratio)变化范围较大, 为0.3~74.一般而言, 较高的VBR表明病毒对细菌群落的控制作用越强[13].比如: 南极洲盐湖的VBR在30.5~126.7之间[34], 极地寡营养湖泊中VBR在1.15~96.7[14,35-36].一般情况下, 营养状况和生产力越大, VBR越大[31,37], 所以极地地区VBR会略低于低纬度地区.冰尘洞中较低的 VBR 并不一定意味着病毒对细菌群落动态的控制作用低[13], Mei和Danovaro认为[38], 尽管VBR较低, 但病毒仍占地中海沉积物中细菌死亡率的40%以上.Danovaro研究表明[39], 沉积物中较低的VBR可能是由腐殖质和富含外酶的复杂有机基质造成的, 它们的存在会引起病毒在感染宿主并被释放之后迅速衰变, 进而导致VBR较低. ...
Viral decay and viral production rates in continental-shelf and deep-sea sediments of the Mediterranean Sea
1
2010
... 病毒在生态系统中时刻变化, 而病毒生产力(VP: Virus production)则是表示病毒活性及病毒侵染状况的一个指标, 其指单位体积单位时间内新的病毒颗粒的释放量.Bellas等[21]的研究表明, 北极Midre Lovénbreen 、 Austre Brøggerbreen 以及 Russell三个冰川的冰尘洞中, VP值介于4.41×107~ 8.98×107 VLP·g-1·h-1之间, 平均值为(7.06±1.8)×107 VLP·g-1·h-1, 细菌碳产生(BCP: Bacterial carbon production)范围为35.4 ~ 73.3 ng C·g-1·h-1, 平均值为(57.8±12.9) ng C·g-1·h-1.Corinaldesi等[40]对地中海沉积物中的病毒生产力进行了研究, 结果表明, 地中海沉积物VP值1.1×107~61.2×107 VLP·g-1·h-1.通过上述研究, 可以看出, 冰尘洞沉积物中VP值大致与海洋沉积物相同.此外, Bellas等[21]的研究结果还表明, VP值与细菌丰度显著正相关, 随细菌丰度的增加, VP值逐渐增大. ...
Analysis of virus genomes from glacial environments reveals novel virus groups with unusual host interactions
2
2015
... 冰川和冰盖等高寒地区拥有惊人的微生物多样性[41].冰尘洞等生态系统的一些极端性质导致他们成为探测新噬菌体序列的理想位置, 这些噬菌体序列可能是这些栖息地所特有的, 也可能是全球普遍存在的噬菌体多样性的一部分[4].随着测序水平的不断提高以及宏基因组学的应用, 关于高寒地区病毒遗传多样性的研究也逐渐增多. ...
... Bellas等[4]对北极 Austre Brøggerbreen和Västre Brøggerbreen两个山谷冰川表面T4噬菌体的遗传多样性进行了研究, 结果表明, 冰尘洞中, T4噬菌体多样性非常丰富, 系统发育分析表明, 冰尘洞沉积物中含有跨越多个T4型噬菌体亚群的混合噬菌体群落, 大多数(71%)的噬菌体序列聚集成三个新的系统发育上不同的组, 而其余的聚类有已知的海洋和土壤衍生的噬菌体序列, 冰尘洞融水中还含有与先前检测到的海洋噬菌体变体相关的另外的不同噬菌体群落, 冰尘洞沉积物与水中的噬菌体组成也有不同.通过对上述两个冰川冰尘洞中的三个双链DNA病毒体组装的假定病毒基因组进行分析, Bellas等[41]进一步发现了其中一种组装噬菌体不仅编码质粒分区基因, 而且聚集有规律的间隔短回文重复(CRISPR)/Cas自适应细菌免疫系统, 其中一个间隔区与另一个噬菌体完全匹配, 表明溶原菌可能赋予其细菌宿主免于其他噬菌体的免疫力.因此冰川环境中存在高度新颖和多样化的病毒群, 其中一些病毒利用非常不寻常的生命策略和基因来控制其复制并与宿主保持长期关系. ...
Importance of prophages to evolution and virulence of bacterial pathogens
1
2013
... 病毒是一种寄生生物, 必须通过侵染特定的宿主, 借助于宿主的营养物质及遗传信息才能够完成自己的生命周期.根据病毒对宿主细胞裂解能力的大小, 可以将病毒的增殖方式分为三种基本感染周期类型: 裂解性感染、 溶源性感染和假溶源性感染[42].裂解性感染能够在短时间内引起宿主细胞的裂解, 进而释放出大量的子代病毒, 因而是病毒繁殖的主要方式[43-44].然而, 溶源性感染通常被看作是病毒用于应对增殖不利条件的一种生活策略[9,31,45].该策略在低温生境尤其是超寡营养湖泊和冰川中发挥着比在其他生境中更大的作用[9].溶源循环或裂解循环是否发生取决于宿主细胞和环境条件, 如水体的营养状态[46].Laybourn-Parry[47]和Säwström等[48]分别在南极湖泊开展了研究, 结果均表明南极湖泊中冬季和春季溶源性发生频率较高, 而夏季则未发现溶源性.Säwström等[49]对北极湖泊的研究结果也表明夏季北极淡水环境中没有溶源性.Bellas等[21]对北极冰尘洞内的沉积物进行了研究, 同样在夏季冰尘洞沉积物中未检测到溶源性.以上结果均表明溶源性感染策略在低温等不利条件下发挥更大的作用.假溶源性是指表面上类似于溶源性, 但是病毒在宿主体内不会立即表达复制, 而是暂时处于休眠状态.该策略被认为能够使病毒在宿主丰度较低且营养限制的情况下在细菌群体中维持自身[31].Säwström等[49]研究发现, 北极和南极洲的超寡营养低温生境中受病毒感染的细胞大多具有非常低的爆发大小.因而在营养限制的条件下, 上述假溶源性策略可能会帮助病毒快速地对外界环境的变化做出反应[31].溶源性策略和假溶源性策略能够帮助部分病毒在极端条件下延长寿命, 从而维持更多的病毒数量和保证多样性[9,50]. ...
Marine viruses and their biogeochemical and ecological effects
3
1999
... 病毒是一种寄生生物, 必须通过侵染特定的宿主, 借助于宿主的营养物质及遗传信息才能够完成自己的生命周期.根据病毒对宿主细胞裂解能力的大小, 可以将病毒的增殖方式分为三种基本感染周期类型: 裂解性感染、 溶源性感染和假溶源性感染[42].裂解性感染能够在短时间内引起宿主细胞的裂解, 进而释放出大量的子代病毒, 因而是病毒繁殖的主要方式[43-44].然而, 溶源性感染通常被看作是病毒用于应对增殖不利条件的一种生活策略[9,31,45].该策略在低温生境尤其是超寡营养湖泊和冰川中发挥着比在其他生境中更大的作用[9].溶源循环或裂解循环是否发生取决于宿主细胞和环境条件, 如水体的营养状态[46].Laybourn-Parry[47]和Säwström等[48]分别在南极湖泊开展了研究, 结果均表明南极湖泊中冬季和春季溶源性发生频率较高, 而夏季则未发现溶源性.Säwström等[49]对北极湖泊的研究结果也表明夏季北极淡水环境中没有溶源性.Bellas等[21]对北极冰尘洞内的沉积物进行了研究, 同样在夏季冰尘洞沉积物中未检测到溶源性.以上结果均表明溶源性感染策略在低温等不利条件下发挥更大的作用.假溶源性是指表面上类似于溶源性, 但是病毒在宿主体内不会立即表达复制, 而是暂时处于休眠状态.该策略被认为能够使病毒在宿主丰度较低且营养限制的情况下在细菌群体中维持自身[31].Säwström等[49]研究发现, 北极和南极洲的超寡营养低温生境中受病毒感染的细胞大多具有非常低的爆发大小.因而在营养限制的条件下, 上述假溶源性策略可能会帮助病毒快速地对外界环境的变化做出反应[31].溶源性策略和假溶源性策略能够帮助部分病毒在极端条件下延长寿命, 从而维持更多的病毒数量和保证多样性[9,50]. ...
... 由于病毒无法控制自己的运动, 只能通过随机接触到达宿主, 因此, 病毒感染宿主的过程是密度依赖性的[43].异养细菌在冰川生态系统中占有绝对的数量优势, 因而它是病毒的主要宿主[21].研究表明病毒可能在冰川等极端环境中对细菌群落施加重要的控制作用[13].在温带淡水环境中, 可见噬菌体感染的细菌细胞的频率(FVIB: Frequency of visibly infected bacteria)介于0.6%~5%之间[51-54].Säwström等[15]对南极湖泊和北极冰尘洞中的FVIB进行了研究, 结果显示, 在南极Cooked湖中, 高FVIB与细菌丰度的峰值一致, 秋季接近一半的细菌明显感染, 而北极冰尘洞与南极湖泊相比, FVIB值较低, 平均值为11.3%.可以看出, 两极湖泊与冰尘洞中的FVIB值明显高于温带淡水湖泊.因此, 可以推断, 病毒感染对于冰尘洞以及极地寡营养淡水湖泊非常重要, 在控制细菌群落动态的过程中发挥着至关重要的作用. ...
... 1983年, Azam首次提出了微生物食物环的概念, 微生物食物环是指异养细菌能够吸收利用生产者生产、 消费者摄食及其他过程中产生的溶解有机物(DOM: Dissolve organic matter), 将其部分转化为自身的颗粒有机物(POM: Particulate organic matter), 并且随着细菌被原生动物摄食, 这些POM又重新回到主食物链中不断循环利用的过程[60].而病毒对于微生物食物环具有重要意义, 使其得到进一步丰富.病毒能够通过裂解宿主, 使得宿主体内的DOM释放出来, 再次被未被病毒感染的异养细菌吸收利用, 从而使得大部分物质和能量在微生物食物环中再循环, 最终被呼吸消耗掉, 产生无机营养物质.这一氧化有机物质、 再生无机营养的环被称为病毒回路[43,61].因而病毒能够将微生物食物环缩短, 催化营养物质从生物体转移到 DOM 和 POM, 将其引导至食物链[5,23], 减少能量向更高营养级的运输[62]. ...
Marine viruses: major players in the global ecosystem
1
2007
... 病毒是一种寄生生物, 必须通过侵染特定的宿主, 借助于宿主的营养物质及遗传信息才能够完成自己的生命周期.根据病毒对宿主细胞裂解能力的大小, 可以将病毒的增殖方式分为三种基本感染周期类型: 裂解性感染、 溶源性感染和假溶源性感染[42].裂解性感染能够在短时间内引起宿主细胞的裂解, 进而释放出大量的子代病毒, 因而是病毒繁殖的主要方式[43-44].然而, 溶源性感染通常被看作是病毒用于应对增殖不利条件的一种生活策略[9,31,45].该策略在低温生境尤其是超寡营养湖泊和冰川中发挥着比在其他生境中更大的作用[9].溶源循环或裂解循环是否发生取决于宿主细胞和环境条件, 如水体的营养状态[46].Laybourn-Parry[47]和Säwström等[48]分别在南极湖泊开展了研究, 结果均表明南极湖泊中冬季和春季溶源性发生频率较高, 而夏季则未发现溶源性.Säwström等[49]对北极湖泊的研究结果也表明夏季北极淡水环境中没有溶源性.Bellas等[21]对北极冰尘洞内的沉积物进行了研究, 同样在夏季冰尘洞沉积物中未检测到溶源性.以上结果均表明溶源性感染策略在低温等不利条件下发挥更大的作用.假溶源性是指表面上类似于溶源性, 但是病毒在宿主体内不会立即表达复制, 而是暂时处于休眠状态.该策略被认为能够使病毒在宿主丰度较低且营养限制的情况下在细菌群体中维持自身[31].Säwström等[49]研究发现, 北极和南极洲的超寡营养低温生境中受病毒感染的细胞大多具有非常低的爆发大小.因而在营养限制的条件下, 上述假溶源性策略可能会帮助病毒快速地对外界环境的变化做出反应[31].溶源性策略和假溶源性策略能够帮助部分病毒在极端条件下延长寿命, 从而维持更多的病毒数量和保证多样性[9,50]. ...
Prophages in marine bacteria: dangerous molecular time bombs or the key to survival in the seas?
1
2008
... 病毒是一种寄生生物, 必须通过侵染特定的宿主, 借助于宿主的营养物质及遗传信息才能够完成自己的生命周期.根据病毒对宿主细胞裂解能力的大小, 可以将病毒的增殖方式分为三种基本感染周期类型: 裂解性感染、 溶源性感染和假溶源性感染[42].裂解性感染能够在短时间内引起宿主细胞的裂解, 进而释放出大量的子代病毒, 因而是病毒繁殖的主要方式[43-44].然而, 溶源性感染通常被看作是病毒用于应对增殖不利条件的一种生活策略[9,31,45].该策略在低温生境尤其是超寡营养湖泊和冰川中发挥着比在其他生境中更大的作用[9].溶源循环或裂解循环是否发生取决于宿主细胞和环境条件, 如水体的营养状态[46].Laybourn-Parry[47]和Säwström等[48]分别在南极湖泊开展了研究, 结果均表明南极湖泊中冬季和春季溶源性发生频率较高, 而夏季则未发现溶源性.Säwström等[49]对北极湖泊的研究结果也表明夏季北极淡水环境中没有溶源性.Bellas等[21]对北极冰尘洞内的沉积物进行了研究, 同样在夏季冰尘洞沉积物中未检测到溶源性.以上结果均表明溶源性感染策略在低温等不利条件下发挥更大的作用.假溶源性是指表面上类似于溶源性, 但是病毒在宿主体内不会立即表达复制, 而是暂时处于休眠状态.该策略被认为能够使病毒在宿主丰度较低且营养限制的情况下在细菌群体中维持自身[31].Säwström等[49]研究发现, 北极和南极洲的超寡营养低温生境中受病毒感染的细胞大多具有非常低的爆发大小.因而在营养限制的条件下, 上述假溶源性策略可能会帮助病毒快速地对外界环境的变化做出反应[31].溶源性策略和假溶源性策略能够帮助部分病毒在极端条件下延长寿命, 从而维持更多的病毒数量和保证多样性[9,50]. ...
Occurrence of lysogenic bacteria in marine microbial communities as determined by prophage induction
1
1996
... 病毒是一种寄生生物, 必须通过侵染特定的宿主, 借助于宿主的营养物质及遗传信息才能够完成自己的生命周期.根据病毒对宿主细胞裂解能力的大小, 可以将病毒的增殖方式分为三种基本感染周期类型: 裂解性感染、 溶源性感染和假溶源性感染[42].裂解性感染能够在短时间内引起宿主细胞的裂解, 进而释放出大量的子代病毒, 因而是病毒繁殖的主要方式[43-44].然而, 溶源性感染通常被看作是病毒用于应对增殖不利条件的一种生活策略[9,31,45].该策略在低温生境尤其是超寡营养湖泊和冰川中发挥着比在其他生境中更大的作用[9].溶源循环或裂解循环是否发生取决于宿主细胞和环境条件, 如水体的营养状态[46].Laybourn-Parry[47]和Säwström等[48]分别在南极湖泊开展了研究, 结果均表明南极湖泊中冬季和春季溶源性发生频率较高, 而夏季则未发现溶源性.Säwström等[49]对北极湖泊的研究结果也表明夏季北极淡水环境中没有溶源性.Bellas等[21]对北极冰尘洞内的沉积物进行了研究, 同样在夏季冰尘洞沉积物中未检测到溶源性.以上结果均表明溶源性感染策略在低温等不利条件下发挥更大的作用.假溶源性是指表面上类似于溶源性, 但是病毒在宿主体内不会立即表达复制, 而是暂时处于休眠状态.该策略被认为能够使病毒在宿主丰度较低且营养限制的情况下在细菌群体中维持自身[31].Säwström等[49]研究发现, 北极和南极洲的超寡营养低温生境中受病毒感染的细胞大多具有非常低的爆发大小.因而在营养限制的条件下, 上述假溶源性策略可能会帮助病毒快速地对外界环境的变化做出反应[31].溶源性策略和假溶源性策略能够帮助部分病毒在极端条件下延长寿命, 从而维持更多的病毒数量和保证多样性[9,50]. ...
Viral dynamics and patterns of lysogeny in saline Antarctic lakes
1
2007
... 病毒是一种寄生生物, 必须通过侵染特定的宿主, 借助于宿主的营养物质及遗传信息才能够完成自己的生命周期.根据病毒对宿主细胞裂解能力的大小, 可以将病毒的增殖方式分为三种基本感染周期类型: 裂解性感染、 溶源性感染和假溶源性感染[42].裂解性感染能够在短时间内引起宿主细胞的裂解, 进而释放出大量的子代病毒, 因而是病毒繁殖的主要方式[43-44].然而, 溶源性感染通常被看作是病毒用于应对增殖不利条件的一种生活策略[9,31,45].该策略在低温生境尤其是超寡营养湖泊和冰川中发挥着比在其他生境中更大的作用[9].溶源循环或裂解循环是否发生取决于宿主细胞和环境条件, 如水体的营养状态[46].Laybourn-Parry[47]和Säwström等[48]分别在南极湖泊开展了研究, 结果均表明南极湖泊中冬季和春季溶源性发生频率较高, 而夏季则未发现溶源性.Säwström等[49]对北极湖泊的研究结果也表明夏季北极淡水环境中没有溶源性.Bellas等[21]对北极冰尘洞内的沉积物进行了研究, 同样在夏季冰尘洞沉积物中未检测到溶源性.以上结果均表明溶源性感染策略在低温等不利条件下发挥更大的作用.假溶源性是指表面上类似于溶源性, 但是病毒在宿主体内不会立即表达复制, 而是暂时处于休眠状态.该策略被认为能够使病毒在宿主丰度较低且营养限制的情况下在细菌群体中维持自身[31].Säwström等[49]研究发现, 北极和南极洲的超寡营养低温生境中受病毒感染的细胞大多具有非常低的爆发大小.因而在营养限制的条件下, 上述假溶源性策略可能会帮助病毒快速地对外界环境的变化做出反应[31].溶源性策略和假溶源性策略能够帮助部分病毒在极端条件下延长寿命, 从而维持更多的病毒数量和保证多样性[9,50]. ...
Seasonal viral loop dynamics in two large ultraoligotrophic Antarctic freshwater lakes
1
2007
... 病毒是一种寄生生物, 必须通过侵染特定的宿主, 借助于宿主的营养物质及遗传信息才能够完成自己的生命周期.根据病毒对宿主细胞裂解能力的大小, 可以将病毒的增殖方式分为三种基本感染周期类型: 裂解性感染、 溶源性感染和假溶源性感染[42].裂解性感染能够在短时间内引起宿主细胞的裂解, 进而释放出大量的子代病毒, 因而是病毒繁殖的主要方式[43-44].然而, 溶源性感染通常被看作是病毒用于应对增殖不利条件的一种生活策略[9,31,45].该策略在低温生境尤其是超寡营养湖泊和冰川中发挥着比在其他生境中更大的作用[9].溶源循环或裂解循环是否发生取决于宿主细胞和环境条件, 如水体的营养状态[46].Laybourn-Parry[47]和Säwström等[48]分别在南极湖泊开展了研究, 结果均表明南极湖泊中冬季和春季溶源性发生频率较高, 而夏季则未发现溶源性.Säwström等[49]对北极湖泊的研究结果也表明夏季北极淡水环境中没有溶源性.Bellas等[21]对北极冰尘洞内的沉积物进行了研究, 同样在夏季冰尘洞沉积物中未检测到溶源性.以上结果均表明溶源性感染策略在低温等不利条件下发挥更大的作用.假溶源性是指表面上类似于溶源性, 但是病毒在宿主体内不会立即表达复制, 而是暂时处于休眠状态.该策略被认为能够使病毒在宿主丰度较低且营养限制的情况下在细菌群体中维持自身[31].Säwström等[49]研究发现, 北极和南极洲的超寡营养低温生境中受病毒感染的细胞大多具有非常低的爆发大小.因而在营养限制的条件下, 上述假溶源性策略可能会帮助病毒快速地对外界环境的变化做出反应[31].溶源性策略和假溶源性策略能够帮助部分病毒在极端条件下延长寿命, 从而维持更多的病毒数量和保证多样性[9,50]. ...
Heterotrophic bacterial and viral dynamics in Arctic freshwaters: results from a field study and nutrient-temperature manipulation experiments
2
2007
... 病毒是一种寄生生物, 必须通过侵染特定的宿主, 借助于宿主的营养物质及遗传信息才能够完成自己的生命周期.根据病毒对宿主细胞裂解能力的大小, 可以将病毒的增殖方式分为三种基本感染周期类型: 裂解性感染、 溶源性感染和假溶源性感染[42].裂解性感染能够在短时间内引起宿主细胞的裂解, 进而释放出大量的子代病毒, 因而是病毒繁殖的主要方式[43-44].然而, 溶源性感染通常被看作是病毒用于应对增殖不利条件的一种生活策略[9,31,45].该策略在低温生境尤其是超寡营养湖泊和冰川中发挥着比在其他生境中更大的作用[9].溶源循环或裂解循环是否发生取决于宿主细胞和环境条件, 如水体的营养状态[46].Laybourn-Parry[47]和Säwström等[48]分别在南极湖泊开展了研究, 结果均表明南极湖泊中冬季和春季溶源性发生频率较高, 而夏季则未发现溶源性.Säwström等[49]对北极湖泊的研究结果也表明夏季北极淡水环境中没有溶源性.Bellas等[21]对北极冰尘洞内的沉积物进行了研究, 同样在夏季冰尘洞沉积物中未检测到溶源性.以上结果均表明溶源性感染策略在低温等不利条件下发挥更大的作用.假溶源性是指表面上类似于溶源性, 但是病毒在宿主体内不会立即表达复制, 而是暂时处于休眠状态.该策略被认为能够使病毒在宿主丰度较低且营养限制的情况下在细菌群体中维持自身[31].Säwström等[49]研究发现, 北极和南极洲的超寡营养低温生境中受病毒感染的细胞大多具有非常低的爆发大小.因而在营养限制的条件下, 上述假溶源性策略可能会帮助病毒快速地对外界环境的变化做出反应[31].溶源性策略和假溶源性策略能够帮助部分病毒在极端条件下延长寿命, 从而维持更多的病毒数量和保证多样性[9,50]. ...
... [49]研究发现, 北极和南极洲的超寡营养低温生境中受病毒感染的细胞大多具有非常低的爆发大小.因而在营养限制的条件下, 上述假溶源性策略可能会帮助病毒快速地对外界环境的变化做出反应[31].溶源性策略和假溶源性策略能够帮助部分病毒在极端条件下延长寿命, 从而维持更多的病毒数量和保证多样性[9,50]. ...
The role of pseudolysogeny in bacteriophage-host interactions in a natural freshwater environment
1
1997
... 病毒是一种寄生生物, 必须通过侵染特定的宿主, 借助于宿主的营养物质及遗传信息才能够完成自己的生命周期.根据病毒对宿主细胞裂解能力的大小, 可以将病毒的增殖方式分为三种基本感染周期类型: 裂解性感染、 溶源性感染和假溶源性感染[42].裂解性感染能够在短时间内引起宿主细胞的裂解, 进而释放出大量的子代病毒, 因而是病毒繁殖的主要方式[43-44].然而, 溶源性感染通常被看作是病毒用于应对增殖不利条件的一种生活策略[9,31,45].该策略在低温生境尤其是超寡营养湖泊和冰川中发挥着比在其他生境中更大的作用[9].溶源循环或裂解循环是否发生取决于宿主细胞和环境条件, 如水体的营养状态[46].Laybourn-Parry[47]和Säwström等[48]分别在南极湖泊开展了研究, 结果均表明南极湖泊中冬季和春季溶源性发生频率较高, 而夏季则未发现溶源性.Säwström等[49]对北极湖泊的研究结果也表明夏季北极淡水环境中没有溶源性.Bellas等[21]对北极冰尘洞内的沉积物进行了研究, 同样在夏季冰尘洞沉积物中未检测到溶源性.以上结果均表明溶源性感染策略在低温等不利条件下发挥更大的作用.假溶源性是指表面上类似于溶源性, 但是病毒在宿主体内不会立即表达复制, 而是暂时处于休眠状态.该策略被认为能够使病毒在宿主丰度较低且营养限制的情况下在细菌群体中维持自身[31].Säwström等[49]研究发现, 北极和南极洲的超寡营养低温生境中受病毒感染的细胞大多具有非常低的爆发大小.因而在营养限制的条件下, 上述假溶源性策略可能会帮助病毒快速地对外界环境的变化做出反应[31].溶源性策略和假溶源性策略能够帮助部分病毒在极端条件下延长寿命, 从而维持更多的病毒数量和保证多样性[9,50]. ...
Significance of bacteriophages for controlling bacterioplankton growth in a Mesotrophic Lake
1
1995
... 由于病毒无法控制自己的运动, 只能通过随机接触到达宿主, 因此, 病毒感染宿主的过程是密度依赖性的[43].异养细菌在冰川生态系统中占有绝对的数量优势, 因而它是病毒的主要宿主[21].研究表明病毒可能在冰川等极端环境中对细菌群落施加重要的控制作用[13].在温带淡水环境中, 可见噬菌体感染的细菌细胞的频率(FVIB: Frequency of visibly infected bacteria)介于0.6%~5%之间[51-54].Säwström等[15]对南极湖泊和北极冰尘洞中的FVIB进行了研究, 结果显示, 在南极Cooked湖中, 高FVIB与细菌丰度的峰值一致, 秋季接近一半的细菌明显感染, 而北极冰尘洞与南极湖泊相比, FVIB值较低, 平均值为11.3%.可以看出, 两极湖泊与冰尘洞中的FVIB值明显高于温带淡水湖泊.因此, 可以推断, 病毒感染对于冰尘洞以及极地寡营养淡水湖泊非常重要, 在控制细菌群落动态的过程中发挥着至关重要的作用. ...
Bacterial carbon production in Lake Erie is influenced by viruses and solar radiation
0
2000
Significance of viral lysis and flagellate grazing as factors controlling bacterioplankton production in a eutrophic lake
0
1998
Estimates of protozoan- and viral-mediated mortality of bacterioplankton in Lake Bourget (France)
1
2005
... 由于病毒无法控制自己的运动, 只能通过随机接触到达宿主, 因此, 病毒感染宿主的过程是密度依赖性的[43].异养细菌在冰川生态系统中占有绝对的数量优势, 因而它是病毒的主要宿主[21].研究表明病毒可能在冰川等极端环境中对细菌群落施加重要的控制作用[13].在温带淡水环境中, 可见噬菌体感染的细菌细胞的频率(FVIB: Frequency of visibly infected bacteria)介于0.6%~5%之间[51-54].Säwström等[15]对南极湖泊和北极冰尘洞中的FVIB进行了研究, 结果显示, 在南极Cooked湖中, 高FVIB与细菌丰度的峰值一致, 秋季接近一半的细菌明显感染, 而北极冰尘洞与南极湖泊相比, FVIB值较低, 平均值为11.3%.可以看出, 两极湖泊与冰尘洞中的FVIB值明显高于温带淡水湖泊.因此, 可以推断, 病毒感染对于冰尘洞以及极地寡营养淡水湖泊非常重要, 在控制细菌群落动态的过程中发挥着至关重要的作用. ...
Global distribution of nearly identical phage-encoded DNA sequences
1
2004
... Bellas等[4]对北极冰川冰尘洞内的病毒进行了移植实验, 将来自冰尘洞和Kongsfjorden海洋峡湾中的病毒分别加入到Kongsfjorden海洋峡湾和冰尘洞的细菌群落中, 观察是否能够发生感染, 结果表明, 来自海洋峡湾的噬菌体能够引发冰尘洞细菌的感染, 由此得出, 海洋以及其他具有全球分布潜力的噬菌体的出现能够为冰尘洞生态系统中不同噬菌体的重组提供机会, 并且能够增加分布广泛的细菌与冰川生态系统中嗜冷宿主之间进行转导的机会.Anesio等[13]也利用Kongsfjord两个山谷冰川中的冰尘洞和冰川前端冰前湖中的病毒与细菌进行了相似的实验研究, 结果发现来自冰尘洞的病毒能够感染冰前湖中的细菌.上述研究结果表明, 病毒能够在生物圈周围移动, 感染类似的宿主, 或者说病毒本质上更具机会性, 具有广泛的宿主感染范围[13,55-56]. ...
Nearly identical bacteriophage structural gene sequences are widely distributed in both marine and freshwater environments
1
2005
... Bellas等[4]对北极冰川冰尘洞内的病毒进行了移植实验, 将来自冰尘洞和Kongsfjorden海洋峡湾中的病毒分别加入到Kongsfjorden海洋峡湾和冰尘洞的细菌群落中, 观察是否能够发生感染, 结果表明, 来自海洋峡湾的噬菌体能够引发冰尘洞细菌的感染, 由此得出, 海洋以及其他具有全球分布潜力的噬菌体的出现能够为冰尘洞生态系统中不同噬菌体的重组提供机会, 并且能够增加分布广泛的细菌与冰川生态系统中嗜冷宿主之间进行转导的机会.Anesio等[13]也利用Kongsfjord两个山谷冰川中的冰尘洞和冰川前端冰前湖中的病毒与细菌进行了相似的实验研究, 结果发现来自冰尘洞的病毒能够感染冰前湖中的细菌.上述研究结果表明, 病毒能够在生物圈周围移动, 感染类似的宿主, 或者说病毒本质上更具机会性, 具有广泛的宿主感染范围[13,55-56]. ...
Are viruses driving microbial diversification and diversity?
1
2003
... 此外, 病毒不仅对宿主的丰度产生影响, 它还能够导致丰富的微生物多样性.病毒通过“杀死胜利者”假说选择宿主[9], 即在生态系统中占主导地位的细菌遇到病毒并被感染和破坏的几率最高.通过这种方式, 病毒的捕食压力能够控制物种数量, 调节微生物群落的多样性[57].同时, “红皇后效应”在一定程度上也会驱动病毒——宿主之间的进化[9].病毒与宿主双方在竞争中均不断进化, 病毒不断地增强毒力以产生更多的感染, 而宿主则不断增加对病毒的抗性, 最终达到相对平衡[58].这种拮抗作用使得病毒和宿主产生更大的遗传和表型多样性[8-9,59]. ...
Antagonistic coevolution between a bacterium and a bacteriophage
1
2002
... 此外, 病毒不仅对宿主的丰度产生影响, 它还能够导致丰富的微生物多样性.病毒通过“杀死胜利者”假说选择宿主[9], 即在生态系统中占主导地位的细菌遇到病毒并被感染和破坏的几率最高.通过这种方式, 病毒的捕食压力能够控制物种数量, 调节微生物群落的多样性[57].同时, “红皇后效应”在一定程度上也会驱动病毒——宿主之间的进化[9].病毒与宿主双方在竞争中均不断进化, 病毒不断地增强毒力以产生更多的感染, 而宿主则不断增加对病毒的抗性, 最终达到相对平衡[58].这种拮抗作用使得病毒和宿主产生更大的遗传和表型多样性[8-9,59]. ...
Antagonistic coevolution accelerates molecular evolution
1
2010
... 此外, 病毒不仅对宿主的丰度产生影响, 它还能够导致丰富的微生物多样性.病毒通过“杀死胜利者”假说选择宿主[9], 即在生态系统中占主导地位的细菌遇到病毒并被感染和破坏的几率最高.通过这种方式, 病毒的捕食压力能够控制物种数量, 调节微生物群落的多样性[57].同时, “红皇后效应”在一定程度上也会驱动病毒——宿主之间的进化[9].病毒与宿主双方在竞争中均不断进化, 病毒不断地增强毒力以产生更多的感染, 而宿主则不断增加对病毒的抗性, 最终达到相对平衡[58].这种拮抗作用使得病毒和宿主产生更大的遗传和表型多样性[8-9,59]. ...
The ecological role of water-column microbes in the sea
1
1983
... 1983年, Azam首次提出了微生物食物环的概念, 微生物食物环是指异养细菌能够吸收利用生产者生产、 消费者摄食及其他过程中产生的溶解有机物(DOM: Dissolve organic matter), 将其部分转化为自身的颗粒有机物(POM: Particulate organic matter), 并且随着细菌被原生动物摄食, 这些POM又重新回到主食物链中不断循环利用的过程[60].而病毒对于微生物食物环具有重要意义, 使其得到进一步丰富.病毒能够通过裂解宿主, 使得宿主体内的DOM释放出来, 再次被未被病毒感染的异养细菌吸收利用, 从而使得大部分物质和能量在微生物食物环中再循环, 最终被呼吸消耗掉, 产生无机营养物质.这一氧化有机物质、 再生无机营养的环被称为病毒回路[43,61].因而病毒能够将微生物食物环缩短, 催化营养物质从生物体转移到 DOM 和 POM, 将其引导至食物链[5,23], 减少能量向更高营养级的运输[62]. ...
Viruses and Nutrient Cycles in the SeaViruses play critical roles in the structure and function of aquatic food webs
1
1999
... 1983年, Azam首次提出了微生物食物环的概念, 微生物食物环是指异养细菌能够吸收利用生产者生产、 消费者摄食及其他过程中产生的溶解有机物(DOM: Dissolve organic matter), 将其部分转化为自身的颗粒有机物(POM: Particulate organic matter), 并且随着细菌被原生动物摄食, 这些POM又重新回到主食物链中不断循环利用的过程[60].而病毒对于微生物食物环具有重要意义, 使其得到进一步丰富.病毒能够通过裂解宿主, 使得宿主体内的DOM释放出来, 再次被未被病毒感染的异养细菌吸收利用, 从而使得大部分物质和能量在微生物食物环中再循环, 最终被呼吸消耗掉, 产生无机营养物质.这一氧化有机物质、 再生无机营养的环被称为病毒回路[43,61].因而病毒能够将微生物食物环缩短, 催化营养物质从生物体转移到 DOM 和 POM, 将其引导至食物链[5,23], 减少能量向更高营养级的运输[62]. ...
Ecological characteristics of marine virioplankton and viruses in the deep biosphere
2
2016
... 1983年, Azam首次提出了微生物食物环的概念, 微生物食物环是指异养细菌能够吸收利用生产者生产、 消费者摄食及其他过程中产生的溶解有机物(DOM: Dissolve organic matter), 将其部分转化为自身的颗粒有机物(POM: Particulate organic matter), 并且随着细菌被原生动物摄食, 这些POM又重新回到主食物链中不断循环利用的过程[60].而病毒对于微生物食物环具有重要意义, 使其得到进一步丰富.病毒能够通过裂解宿主, 使得宿主体内的DOM释放出来, 再次被未被病毒感染的异养细菌吸收利用, 从而使得大部分物质和能量在微生物食物环中再循环, 最终被呼吸消耗掉, 产生无机营养物质.这一氧化有机物质、 再生无机营养的环被称为病毒回路[43,61].因而病毒能够将微生物食物环缩短, 催化营养物质从生物体转移到 DOM 和 POM, 将其引导至食物链[5,23], 减少能量向更高营养级的运输[62]. ...
... (3)病毒的分离培养: 由于病毒的生存必须依赖于宿主, 而环境中绝大多数的宿主微生物是不可培养的, 因为病毒的分离培养受到了极大限制[62].目前, 只有极少的环境病毒分离培养成功, 而冰尘病毒分离培养至今尚未实现. ...
海洋浮游病毒和深部生物圈病毒的生态特性
2
2016
... 1983年, Azam首次提出了微生物食物环的概念, 微生物食物环是指异养细菌能够吸收利用生产者生产、 消费者摄食及其他过程中产生的溶解有机物(DOM: Dissolve organic matter), 将其部分转化为自身的颗粒有机物(POM: Particulate organic matter), 并且随着细菌被原生动物摄食, 这些POM又重新回到主食物链中不断循环利用的过程[60].而病毒对于微生物食物环具有重要意义, 使其得到进一步丰富.病毒能够通过裂解宿主, 使得宿主体内的DOM释放出来, 再次被未被病毒感染的异养细菌吸收利用, 从而使得大部分物质和能量在微生物食物环中再循环, 最终被呼吸消耗掉, 产生无机营养物质.这一氧化有机物质、 再生无机营养的环被称为病毒回路[43,61].因而病毒能够将微生物食物环缩短, 催化营养物质从生物体转移到 DOM 和 POM, 将其引导至食物链[5,23], 减少能量向更高营养级的运输[62]. ...
... (3)病毒的分离培养: 由于病毒的生存必须依赖于宿主, 而环境中绝大多数的宿主微生物是不可培养的, 因为病毒的分离培养受到了极大限制[62].目前, 只有极少的环境病毒分离培养成功, 而冰尘病毒分离培养至今尚未实现. ...
Major viral impact on the functioning of benthic deep-sea ecosystems
2
2008
... 冰尘洞是微生物主导的生态系统, 微生物食物环可能是能量和碳流动的主要途径[13].Danovaro等[63]研究表明病毒诱导的细菌死亡率(VIBM: Virus induced bacterial mortality)可以解释全球深海底栖生态系统中约80%的细菌产量减少量, 使细菌产量与二级消费者分离.Danovaro发现淡水沉积物中VIBM约为10%[64], 海洋沉积物中VIBM为31%~89%[63-64].而Bellas等[21]的研究结果表明, 北极冰尘洞沉积物中VIBM值大于100%, 并且计算出病毒是造成冰尘洞中598%~1710%细菌产量减少的原因.由此得知, 冰尘洞中的VIBM远高于湖泊和海洋.这里计算的高VIBM意味着冰尘洞生态系统中的大部分碳流经病毒介导的过程, 因此, 病毒很有可能在冰尘洞等寒冷、 寡营养生态系统的碳和营养物质的循环中发挥着重要的作用[13,21].Säwström等[65]的研究结果也证明了病毒在溶解有机碳的循环和控制冰川中的细菌死亡率方面发挥着核心作用.因此, 病毒诱导细菌死亡的过程是冰上生态系统中碳和营养物质释放和再循环的主导过程. ...
... [63-64].而Bellas等[21]的研究结果表明, 北极冰尘洞沉积物中VIBM值大于100%, 并且计算出病毒是造成冰尘洞中598%~1710%细菌产量减少的原因.由此得知, 冰尘洞中的VIBM远高于湖泊和海洋.这里计算的高VIBM意味着冰尘洞生态系统中的大部分碳流经病毒介导的过程, 因此, 病毒很有可能在冰尘洞等寒冷、 寡营养生态系统的碳和营养物质的循环中发挥着重要的作用[13,21].Säwström等[65]的研究结果也证明了病毒在溶解有机碳的循环和控制冰川中的细菌死亡率方面发挥着核心作用.因此, 病毒诱导细菌死亡的过程是冰上生态系统中碳和营养物质释放和再循环的主导过程. ...
Viriobenthos in freshwater and marine sediments: a review
2
2008
... 冰尘洞是微生物主导的生态系统, 微生物食物环可能是能量和碳流动的主要途径[13].Danovaro等[63]研究表明病毒诱导的细菌死亡率(VIBM: Virus induced bacterial mortality)可以解释全球深海底栖生态系统中约80%的细菌产量减少量, 使细菌产量与二级消费者分离.Danovaro发现淡水沉积物中VIBM约为10%[64], 海洋沉积物中VIBM为31%~89%[63-64].而Bellas等[21]的研究结果表明, 北极冰尘洞沉积物中VIBM值大于100%, 并且计算出病毒是造成冰尘洞中598%~1710%细菌产量减少的原因.由此得知, 冰尘洞中的VIBM远高于湖泊和海洋.这里计算的高VIBM意味着冰尘洞生态系统中的大部分碳流经病毒介导的过程, 因此, 病毒很有可能在冰尘洞等寒冷、 寡营养生态系统的碳和营养物质的循环中发挥着重要的作用[13,21].Säwström等[65]的研究结果也证明了病毒在溶解有机碳的循环和控制冰川中的细菌死亡率方面发挥着核心作用.因此, 病毒诱导细菌死亡的过程是冰上生态系统中碳和营养物质释放和再循环的主导过程. ...
... -64].而Bellas等[21]的研究结果表明, 北极冰尘洞沉积物中VIBM值大于100%, 并且计算出病毒是造成冰尘洞中598%~1710%细菌产量减少的原因.由此得知, 冰尘洞中的VIBM远高于湖泊和海洋.这里计算的高VIBM意味着冰尘洞生态系统中的大部分碳流经病毒介导的过程, 因此, 病毒很有可能在冰尘洞等寒冷、 寡营养生态系统的碳和营养物质的循环中发挥着重要的作用[13,21].Säwström等[65]的研究结果也证明了病毒在溶解有机碳的循环和控制冰川中的细菌死亡率方面发挥着核心作用.因此, 病毒诱导细菌死亡的过程是冰上生态系统中碳和营养物质释放和再循环的主导过程. ...
High viral infection rates in Antarctic and Arctic bacterioplankton
1
2007
... 冰尘洞是微生物主导的生态系统, 微生物食物环可能是能量和碳流动的主要途径[13].Danovaro等[63]研究表明病毒诱导的细菌死亡率(VIBM: Virus induced bacterial mortality)可以解释全球深海底栖生态系统中约80%的细菌产量减少量, 使细菌产量与二级消费者分离.Danovaro发现淡水沉积物中VIBM约为10%[64], 海洋沉积物中VIBM为31%~89%[63-64].而Bellas等[21]的研究结果表明, 北极冰尘洞沉积物中VIBM值大于100%, 并且计算出病毒是造成冰尘洞中598%~1710%细菌产量减少的原因.由此得知, 冰尘洞中的VIBM远高于湖泊和海洋.这里计算的高VIBM意味着冰尘洞生态系统中的大部分碳流经病毒介导的过程, 因此, 病毒很有可能在冰尘洞等寒冷、 寡营养生态系统的碳和营养物质的循环中发挥着重要的作用[13,21].Säwström等[65]的研究结果也证明了病毒在溶解有机碳的循环和控制冰川中的细菌死亡率方面发挥着核心作用.因此, 病毒诱导细菌死亡的过程是冰上生态系统中碳和营养物质释放和再循环的主导过程. ...
Advances in viral ecology research
1
2016
... 目前病毒的研究虽然取得了迅速发展, 但是多集中于湖泊与海洋等水生环境[66], 冰尘洞内病毒的研究还存在较多的空白.下面是目前冰尘洞内病毒研究尚未涉及的几个方面, 在此提出, 供日后参考与完善: ...
病毒生态学研究进展
1
2016
... 目前病毒的研究虽然取得了迅速发展, 但是多集中于湖泊与海洋等水生环境[66], 冰尘洞内病毒的研究还存在较多的空白.下面是目前冰尘洞内病毒研究尚未涉及的几个方面, 在此提出, 供日后参考与完善: ...
Advances in the study of marine viruses
1
2015
... (1)形态多样性: 目前对病毒形态多样性的了解主要来自于透射电镜的观测结果.病毒形态结构多样, 主要包括蝌蚪状、 杆状、 丝状、 子弹状、 纺锤形、 球形、 六边形、 柠檬型、 多边形等[67].目前, 对两极地区湖泊与冰碛中病毒的形态结构进行了少量的观察研究, 结果表明, 除了常见的病毒形态外, 还存在少量较为少见的异常形态类型[68].但是, 关于冰尘洞内病毒形态多样性的研究目前仍是空白, 冰尘洞内病毒形态结构是否具有一定程度的特殊性, 我们依然无法知晓. ...
Transmission electron microscope analysis of virus-like particles in the freshwater lakes of Signy Island, Antarctica
1
2000
... (1)形态多样性: 目前对病毒形态多样性的了解主要来自于透射电镜的观测结果.病毒形态结构多样, 主要包括蝌蚪状、 杆状、 丝状、 子弹状、 纺锤形、 球形、 六边形、 柠檬型、 多边形等[67].目前, 对两极地区湖泊与冰碛中病毒的形态结构进行了少量的观察研究, 结果表明, 除了常见的病毒形态外, 还存在少量较为少见的异常形态类型[68].但是, 关于冰尘洞内病毒形态多样性的研究目前仍是空白, 冰尘洞内病毒形态结构是否具有一定程度的特殊性, 我们依然无法知晓. ...
Production and decay of viruses in aquatic environments
1
1991
... (2)病毒降解率: 病毒降解率是指病毒颗粒随时间的减少量[69], 是病毒生态功能研究中一个重要的参数.一般说来, 病毒降解率与营养条件、 太阳辐射、 温度等均密切相关[70-72].冰尘洞作为冰川生态系统一个独特的生境, 冰尘洞内病毒降解率是否与海洋或湖泊等水生环境存在差异, 至今仍未有研究. ...
Viruses and protists cause similar bacterial mortality in coastal seawater
1
1995
... (2)病毒降解率: 病毒降解率是指病毒颗粒随时间的减少量[69], 是病毒生态功能研究中一个重要的参数.一般说来, 病毒降解率与营养条件、 太阳辐射、 温度等均密切相关[70-72].冰尘洞作为冰川生态系统一个独特的生境, 冰尘洞内病毒降解率是否与海洋或湖泊等水生环境存在差异, 至今仍未有研究. ...
The role of sunlight in the removal and repair of viruses in the sea
0
1998
Effects of temperature and light on stability of microalgal viruses, HaV, HcV and HcRNAV
1
2005
... (2)病毒降解率: 病毒降解率是指病毒颗粒随时间的减少量[69], 是病毒生态功能研究中一个重要的参数.一般说来, 病毒降解率与营养条件、 太阳辐射、 温度等均密切相关[70-72].冰尘洞作为冰川生态系统一个独特的生境, 冰尘洞内病毒降解率是否与海洋或湖泊等水生环境存在差异, 至今仍未有研究. ...
Assessment and analysis of the freeze-thaw erosion sensitivity on the Tibetan Plateau
1
2017
... 但是, 目前关于冰川尤其是冰尘洞中病毒的研究依然稀少, 并且为数不多的研究大多集中于南北极的湖泊与北极冰尘, 关于青藏高原冰尘洞内病毒的研究至今仍未见报道.而青藏高原被称为“第三极”[73], 其具有独特的地理位置特征, 深居内陆, 纬度低, 但是海拔高, 是地球除了南北极之外人们最感兴趣的冰川研究热点地区[74].由于青藏高原冰川环境的特殊性, 其上的微生物研究具有重要的意义, 因而开展我国青藏高原冰川冰尘洞内病毒研究工作具有巨大研究前景.未来我们可以围绕以下几个方面展开: ...
青藏高原冻融侵蚀敏感性评价与分析
1
2017
... 但是, 目前关于冰川尤其是冰尘洞中病毒的研究依然稀少, 并且为数不多的研究大多集中于南北极的湖泊与北极冰尘, 关于青藏高原冰尘洞内病毒的研究至今仍未见报道.而青藏高原被称为“第三极”[73], 其具有独特的地理位置特征, 深居内陆, 纬度低, 但是海拔高, 是地球除了南北极之外人们最感兴趣的冰川研究热点地区[74].由于青藏高原冰川环境的特殊性, 其上的微生物研究具有重要的意义, 因而开展我国青藏高原冰川冰尘洞内病毒研究工作具有巨大研究前景.未来我们可以围绕以下几个方面展开: ...
Advances in researches of microorganisms in glacial snow and ice on the Qinghai-Tibet Plateau
1
2007
... 但是, 目前关于冰川尤其是冰尘洞中病毒的研究依然稀少, 并且为数不多的研究大多集中于南北极的湖泊与北极冰尘, 关于青藏高原冰尘洞内病毒的研究至今仍未见报道.而青藏高原被称为“第三极”[73], 其具有独特的地理位置特征, 深居内陆, 纬度低, 但是海拔高, 是地球除了南北极之外人们最感兴趣的冰川研究热点地区[74].由于青藏高原冰川环境的特殊性, 其上的微生物研究具有重要的意义, 因而开展我国青藏高原冰川冰尘洞内病毒研究工作具有巨大研究前景.未来我们可以围绕以下几个方面展开: ...
青藏高原冰川雪冰微生物研究进展
1
2007
... 但是, 目前关于冰川尤其是冰尘洞中病毒的研究依然稀少, 并且为数不多的研究大多集中于南北极的湖泊与北极冰尘, 关于青藏高原冰尘洞内病毒的研究至今仍未见报道.而青藏高原被称为“第三极”[73], 其具有独特的地理位置特征, 深居内陆, 纬度低, 但是海拔高, 是地球除了南北极之外人们最感兴趣的冰川研究热点地区[74].由于青藏高原冰川环境的特殊性, 其上的微生物研究具有重要的意义, 因而开展我国青藏高原冰川冰尘洞内病毒研究工作具有巨大研究前景.未来我们可以围绕以下几个方面展开: ...
/
|
〈
|
![]()
|
〉
|
| 

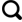



 甘公网安备 62010202000676号
甘公网安备 62010202000676号
{kind=link}
{kind=link}
{kind=link}
{kind=link}