5
2003
... 生物结皮是由生长在表层土壤的微生物、藻类、苔藓和地衣等个体微小的生物成分与土壤相互作用形成的复合层[1].其广泛分布于干旱、半干旱区和极地、亚极地等脆弱生态区[2-3].近年来,关于生物结皮生态功能的研究已详细展开,研究表明生物结皮的形成可以改善土壤理化性质,提高土壤稳定性,改善土壤肥力等,对干旱半干旱地区植被恢复与重建具有重要的生态学意义[3-4].目前关于生物结皮生态功能的研究已有很多,但这些研究主要集中在干旱半干旱的沙漠地区[5-14],例如美国科罗拉多沙漠及其西南部的莫哈维沙漠[6-7],以色列南部的内盖夫沙漠[8],阿根廷的蒙特沙漠[9]以及干旱半干旱的西班牙地中海地区[10].国内关于生物结皮的研究也主要集中在沙漠及干旱半干旱地区,如黄土丘陵沟壑区[11-12]、腾格尔沙漠地区[13-14]、西北及新疆沙漠区[15-16].我国青藏高原素有“世界屋脊”、“第三极”之称,其海拔高,气温低,辐射强烈,日照多,黄河源区也源自于此[17].黄河源区是我国青藏高原重要的水源涵养区,也是该区域最主要的畜牧业基地之一.源区内独特的气候条件及其丰富的自然生态资源为该区经济发展及生态平衡发挥了生态安全屏障的作用[17-18].然而,20世纪70年代以来,随着气候的逐渐干旱化和人口、牲畜数量的迅速增加,该区生态环境急剧恶化,大面积植被退化,20世纪末,植被退化面积占源区草地总面积1/3[19].2005年以来,随着三江源地区实施草地“生态保护和建设”过程的具体实施(围栏封育、退牧还草、沙漠化治理、湿地保护等)[19-20],生物结皮开始在该地区逐渐发育.笔者在野外实地考察过程中发现,生物结皮在黄河源区不同退化程度的草地斑块中均有较大面积的分布.黄河源区平均气温-5.38~4.14 ℃,年降水量262.2~772.8 mm[20],在如此相对湿润且温度极低的条件下,生物结皮的发育特征与干热的沙漠地区是否一样,目前还不得而知.生物结皮作为地表特殊的结构层,其属于变水生物[21],在湿润时才能够进行新陈代谢活动[22].而青藏高原高黄河源区属于高原亚寒带半干旱区,仅有冷暖两季,冷季漫长,从每年10月初至次年4月之间地表处于冻结状态[23],降水主要以固态降雪为主,生物结皮呈现休眠状态.在半干旱低温环境条件下,生物结皮的理化性状如何,这一问题也尚不明晰. ...
... 生物结皮可以在各种气候条件下广泛发育[1],国内外关于生物结皮的研究主要集中在荒漠地区及干旱半干旱的黄土高原等地区[4-16],关于青藏高原高寒冻土区生物结皮的发育特点鲜有报道.本研究结果表明冻土区生物结皮主要是以藻为主的生物结皮,结皮层厚度大约为6~22 mm.其在厚度上大于干旱的荒漠地区藻类结皮厚度(1~5 mm)[16].这与该地区特殊的生态环境条件密切相关,由于冻土区常年温度低,生物结皮活性较弱,呼吸等生理活性缓慢[4,21],其中积累的物质不易分解,因此,生物结皮层厚度较大.同时,生物结皮的厚度与生物结皮的发育年限密切相关[29].因此,该区域植被退化区域生物结皮(浅色藻结皮)较未退化区生物结皮(深色藻结皮)发育年限短,故其深色藻结皮厚度大于浅色藻结生物结皮小. ...
... 生物结皮对土壤理化属性具有显著影响[1,3,14-15,28].关于生物结皮对土壤颗粒组成的影响,在不同的研究区域所得的研究结论存在一定分歧.有些研究结果表明,生物结皮的存在,能够细化土壤颗粒[28,34-35].也有研究表明,生物结皮对土壤颗粒组成并没有显著影响[36].本研究结果表明,由于生物结皮的发育可增加土壤中粉粒含量,5~20 cm土层中粉粒表现为深色藻结皮>浅色藻结皮>裸地,但差异并不显著.来自沙区的研究结果表明,生物结皮能够黏化土壤,捕捉土壤中细颗粒,使得表层土壤中黏粒和粉粒含量呈显著增加趋势.在黄土区的研究结果也表明,随着生物结皮的发育,结皮层土壤细颗相对增加[28].这些研究结果均表明生物结皮对土壤颗粒组成的影响集中在结皮层及其下0~5 cm左右,细颗粒呈现明显的表聚现象[28].而本研究结果发现由于结皮层的覆盖土壤粉粒在20~30 cm土层显著增加,这可能与该区域强烈的冻融过程及水热迁移有关.由于该区域早晚、四季之间温差较大,呈现明显的冻融循环作用[24-25],因此生物结皮层细颗粒可能由于冻融作用及水分迁移随之向下移动,聚集在其下20~30 cm土层.这种迁移作用可能随着水分含量以及发育年限的不同而发生相应的变化.与此同时,本研究结果表明,冻土区生物结皮层土壤容重也显著低于下层土壤,同时深色藻结皮不仅显著降低了自身容重,也影响到了其下0~20 cm土层土壤容重.而浅色藻结皮则对下层土壤容重影响不明显.本研究区海拔高于4 200 m,海拔高、风力大,与沙漠区域不同,生物结皮在形成过程中,并没有显著的捕捉细颗粒的作用,使结皮层土壤细化从而降低结皮层土壤容重.生物结皮在形成过程中,通过菌丝体、藻丝、假根或分泌的多糖来捆绑、黏结土壤颗粒,使得土壤形成许多团聚结构,土壤中孔隙增大,土壤容重降低[34],而深色藻结皮发育年限较长,厚度较厚,其孔隙度高,因此,容重较浅色藻结皮小,田间持水量显著高于浅色藻结皮及裸地. ...
... 生物结皮的形成,不仅可以改善土壤结构[28,37],也可显著影响表层土壤的养分累积[9,11,15,21].生物结皮中的固氮蓝藻的存在,可将大气中的氮固定从而输入到表层土壤中[38].同时生物结皮中的微生物能够进行固碳作用,可以增加表层土壤有机质含量[1,14,28],增加土壤肥力.虽然生物结皮对土壤养分积累效应已经形成一致结论,但其作用程度在不同区域还有所不同[14,28,40].本研究结果表明该区两种类型生物结皮均显著增加了结皮层及其下0~20 cm土层土壤有机质含量.藻类是生物结皮发育初期的先锋物种,它的形成能够向体外分泌以多糖为主的物质,从而形成生物结皮[41].同时,在给予充足的水分和光照条件下,生物结皮能够进行固碳作用,其固碳潜力主要取决于生物结皮的生物组成,研究已表明藓结皮的固碳能力高于藻结皮[42].该区域两种类型生物结皮,均以藻类结皮为主,因此其固碳能力可能没有显著差异.同时死亡的藻体又成为土壤有机物,为其他异氧微生物提供碳源[1],因此两种类型生物结皮均依靠显著增加了结皮层及其下0~20 cm土层土壤有机质含量.然而,两种生物结皮对土壤全氮含量的影响则有所不同.深色藻结皮的形成同时也提高了结皮层及其下0~20 cm土层土壤全氮含量,浅色藻结皮则仅提高了结皮层土层土壤全氮含量,对其下0~20 cm土层土壤全氮含量没有显著影响.生物结皮有机体在湿润状态下才能够进行生理代谢活动,进行相应的固氮作用,并且不同的物种固氮所需最佳水温条件不同[6].生物结皮在湿润后,其固氮活性强烈受制于水分[43].以上研究也表明,深色藻结皮的结皮层田间持水量显著高于浅色藻结皮,因此其固氮持续时间可能较长,故其固氮作用较强,向土壤中输入的氮素较多.氮素在水热交换作用下,向下层输入.而浅色藻结皮持水量较低,固氮作用较弱,因此其固定的氮素仅仅存留在结皮层.同时,生物结皮的存在降低了结皮层pH,但其对结皮层及下层土壤pH并没有显著影响.由于生物结皮某些藻类和地衣能够在湿润状态下进行生理活动,释放有机酸,使土壤pH降低[43],青藏高原黄河源区这一高寒地区气温常年低下,并且降水多以固态或暴雨的形式[44],并且在低温条件下,生物结皮有机体产生的有机酸较少也不易挥发释放出来,因此对土壤pH影响较小.高寒地区生物结皮对土壤养分影响过程及机理还需进一步研究. ...
... [1],因此两种类型生物结皮均依靠显著增加了结皮层及其下0~20 cm土层土壤有机质含量.然而,两种生物结皮对土壤全氮含量的影响则有所不同.深色藻结皮的形成同时也提高了结皮层及其下0~20 cm土层土壤全氮含量,浅色藻结皮则仅提高了结皮层土层土壤全氮含量,对其下0~20 cm土层土壤全氮含量没有显著影响.生物结皮有机体在湿润状态下才能够进行生理代谢活动,进行相应的固氮作用,并且不同的物种固氮所需最佳水温条件不同[6].生物结皮在湿润后,其固氮活性强烈受制于水分[43].以上研究也表明,深色藻结皮的结皮层田间持水量显著高于浅色藻结皮,因此其固氮持续时间可能较长,故其固氮作用较强,向土壤中输入的氮素较多.氮素在水热交换作用下,向下层输入.而浅色藻结皮持水量较低,固氮作用较弱,因此其固定的氮素仅仅存留在结皮层.同时,生物结皮的存在降低了结皮层pH,但其对结皮层及下层土壤pH并没有显著影响.由于生物结皮某些藻类和地衣能够在湿润状态下进行生理活动,释放有机酸,使土壤pH降低[43],青藏高原黄河源区这一高寒地区气温常年低下,并且降水多以固态或暴雨的形式[44],并且在低温条件下,生物结皮有机体产生的有机酸较少也不易挥发释放出来,因此对土壤pH影响较小.高寒地区生物结皮对土壤养分影响过程及机理还需进一步研究. ...
Response of biological soil crusts (BSC) from arid-semiarid and polar region to global climate change
1
2008
... 生物结皮是由生长在表层土壤的微生物、藻类、苔藓和地衣等个体微小的生物成分与土壤相互作用形成的复合层[1].其广泛分布于干旱、半干旱区和极地、亚极地等脆弱生态区[2-3].近年来,关于生物结皮生态功能的研究已详细展开,研究表明生物结皮的形成可以改善土壤理化性质,提高土壤稳定性,改善土壤肥力等,对干旱半干旱地区植被恢复与重建具有重要的生态学意义[3-4].目前关于生物结皮生态功能的研究已有很多,但这些研究主要集中在干旱半干旱的沙漠地区[5-14],例如美国科罗拉多沙漠及其西南部的莫哈维沙漠[6-7],以色列南部的内盖夫沙漠[8],阿根廷的蒙特沙漠[9]以及干旱半干旱的西班牙地中海地区[10].国内关于生物结皮的研究也主要集中在沙漠及干旱半干旱地区,如黄土丘陵沟壑区[11-12]、腾格尔沙漠地区[13-14]、西北及新疆沙漠区[15-16].我国青藏高原素有“世界屋脊”、“第三极”之称,其海拔高,气温低,辐射强烈,日照多,黄河源区也源自于此[17].黄河源区是我国青藏高原重要的水源涵养区,也是该区域最主要的畜牧业基地之一.源区内独特的气候条件及其丰富的自然生态资源为该区经济发展及生态平衡发挥了生态安全屏障的作用[17-18].然而,20世纪70年代以来,随着气候的逐渐干旱化和人口、牲畜数量的迅速增加,该区生态环境急剧恶化,大面积植被退化,20世纪末,植被退化面积占源区草地总面积1/3[19].2005年以来,随着三江源地区实施草地“生态保护和建设”过程的具体实施(围栏封育、退牧还草、沙漠化治理、湿地保护等)[19-20],生物结皮开始在该地区逐渐发育.笔者在野外实地考察过程中发现,生物结皮在黄河源区不同退化程度的草地斑块中均有较大面积的分布.黄河源区平均气温-5.38~4.14 ℃,年降水量262.2~772.8 mm[20],在如此相对湿润且温度极低的条件下,生物结皮的发育特征与干热的沙漠地区是否一样,目前还不得而知.生物结皮作为地表特殊的结构层,其属于变水生物[21],在湿润时才能够进行新陈代谢活动[22].而青藏高原高黄河源区属于高原亚寒带半干旱区,仅有冷暖两季,冷季漫长,从每年10月初至次年4月之间地表处于冻结状态[23],降水主要以固态降雪为主,生物结皮呈现休眠状态.在半干旱低温环境条件下,生物结皮的理化性状如何,这一问题也尚不明晰. ...
生物土壤结皮对全球气候变化的响应
1
2008
... 生物结皮是由生长在表层土壤的微生物、藻类、苔藓和地衣等个体微小的生物成分与土壤相互作用形成的复合层[1].其广泛分布于干旱、半干旱区和极地、亚极地等脆弱生态区[2-3].近年来,关于生物结皮生态功能的研究已详细展开,研究表明生物结皮的形成可以改善土壤理化性质,提高土壤稳定性,改善土壤肥力等,对干旱半干旱地区植被恢复与重建具有重要的生态学意义[3-4].目前关于生物结皮生态功能的研究已有很多,但这些研究主要集中在干旱半干旱的沙漠地区[5-14],例如美国科罗拉多沙漠及其西南部的莫哈维沙漠[6-7],以色列南部的内盖夫沙漠[8],阿根廷的蒙特沙漠[9]以及干旱半干旱的西班牙地中海地区[10].国内关于生物结皮的研究也主要集中在沙漠及干旱半干旱地区,如黄土丘陵沟壑区[11-12]、腾格尔沙漠地区[13-14]、西北及新疆沙漠区[15-16].我国青藏高原素有“世界屋脊”、“第三极”之称,其海拔高,气温低,辐射强烈,日照多,黄河源区也源自于此[17].黄河源区是我国青藏高原重要的水源涵养区,也是该区域最主要的畜牧业基地之一.源区内独特的气候条件及其丰富的自然生态资源为该区经济发展及生态平衡发挥了生态安全屏障的作用[17-18].然而,20世纪70年代以来,随着气候的逐渐干旱化和人口、牲畜数量的迅速增加,该区生态环境急剧恶化,大面积植被退化,20世纪末,植被退化面积占源区草地总面积1/3[19].2005年以来,随着三江源地区实施草地“生态保护和建设”过程的具体实施(围栏封育、退牧还草、沙漠化治理、湿地保护等)[19-20],生物结皮开始在该地区逐渐发育.笔者在野外实地考察过程中发现,生物结皮在黄河源区不同退化程度的草地斑块中均有较大面积的分布.黄河源区平均气温-5.38~4.14 ℃,年降水量262.2~772.8 mm[20],在如此相对湿润且温度极低的条件下,生物结皮的发育特征与干热的沙漠地区是否一样,目前还不得而知.生物结皮作为地表特殊的结构层,其属于变水生物[21],在湿润时才能够进行新陈代谢活动[22].而青藏高原高黄河源区属于高原亚寒带半干旱区,仅有冷暖两季,冷季漫长,从每年10月初至次年4月之间地表处于冻结状态[23],降水主要以固态降雪为主,生物结皮呈现休眠状态.在半干旱低温环境条件下,生物结皮的理化性状如何,这一问题也尚不明晰. ...
Ecological function of biological soil crusts
3
2002
... 生物结皮是由生长在表层土壤的微生物、藻类、苔藓和地衣等个体微小的生物成分与土壤相互作用形成的复合层[1].其广泛分布于干旱、半干旱区和极地、亚极地等脆弱生态区[2-3].近年来,关于生物结皮生态功能的研究已详细展开,研究表明生物结皮的形成可以改善土壤理化性质,提高土壤稳定性,改善土壤肥力等,对干旱半干旱地区植被恢复与重建具有重要的生态学意义[3-4].目前关于生物结皮生态功能的研究已有很多,但这些研究主要集中在干旱半干旱的沙漠地区[5-14],例如美国科罗拉多沙漠及其西南部的莫哈维沙漠[6-7],以色列南部的内盖夫沙漠[8],阿根廷的蒙特沙漠[9]以及干旱半干旱的西班牙地中海地区[10].国内关于生物结皮的研究也主要集中在沙漠及干旱半干旱地区,如黄土丘陵沟壑区[11-12]、腾格尔沙漠地区[13-14]、西北及新疆沙漠区[15-16].我国青藏高原素有“世界屋脊”、“第三极”之称,其海拔高,气温低,辐射强烈,日照多,黄河源区也源自于此[17].黄河源区是我国青藏高原重要的水源涵养区,也是该区域最主要的畜牧业基地之一.源区内独特的气候条件及其丰富的自然生态资源为该区经济发展及生态平衡发挥了生态安全屏障的作用[17-18].然而,20世纪70年代以来,随着气候的逐渐干旱化和人口、牲畜数量的迅速增加,该区生态环境急剧恶化,大面积植被退化,20世纪末,植被退化面积占源区草地总面积1/3[19].2005年以来,随着三江源地区实施草地“生态保护和建设”过程的具体实施(围栏封育、退牧还草、沙漠化治理、湿地保护等)[19-20],生物结皮开始在该地区逐渐发育.笔者在野外实地考察过程中发现,生物结皮在黄河源区不同退化程度的草地斑块中均有较大面积的分布.黄河源区平均气温-5.38~4.14 ℃,年降水量262.2~772.8 mm[20],在如此相对湿润且温度极低的条件下,生物结皮的发育特征与干热的沙漠地区是否一样,目前还不得而知.生物结皮作为地表特殊的结构层,其属于变水生物[21],在湿润时才能够进行新陈代谢活动[22].而青藏高原高黄河源区属于高原亚寒带半干旱区,仅有冷暖两季,冷季漫长,从每年10月初至次年4月之间地表处于冻结状态[23],降水主要以固态降雪为主,生物结皮呈现休眠状态.在半干旱低温环境条件下,生物结皮的理化性状如何,这一问题也尚不明晰. ...
... [3-4].目前关于生物结皮生态功能的研究已有很多,但这些研究主要集中在干旱半干旱的沙漠地区[5-14],例如美国科罗拉多沙漠及其西南部的莫哈维沙漠[6-7],以色列南部的内盖夫沙漠[8],阿根廷的蒙特沙漠[9]以及干旱半干旱的西班牙地中海地区[10].国内关于生物结皮的研究也主要集中在沙漠及干旱半干旱地区,如黄土丘陵沟壑区[11-12]、腾格尔沙漠地区[13-14]、西北及新疆沙漠区[15-16].我国青藏高原素有“世界屋脊”、“第三极”之称,其海拔高,气温低,辐射强烈,日照多,黄河源区也源自于此[17].黄河源区是我国青藏高原重要的水源涵养区,也是该区域最主要的畜牧业基地之一.源区内独特的气候条件及其丰富的自然生态资源为该区经济发展及生态平衡发挥了生态安全屏障的作用[17-18].然而,20世纪70年代以来,随着气候的逐渐干旱化和人口、牲畜数量的迅速增加,该区生态环境急剧恶化,大面积植被退化,20世纪末,植被退化面积占源区草地总面积1/3[19].2005年以来,随着三江源地区实施草地“生态保护和建设”过程的具体实施(围栏封育、退牧还草、沙漠化治理、湿地保护等)[19-20],生物结皮开始在该地区逐渐发育.笔者在野外实地考察过程中发现,生物结皮在黄河源区不同退化程度的草地斑块中均有较大面积的分布.黄河源区平均气温-5.38~4.14 ℃,年降水量262.2~772.8 mm[20],在如此相对湿润且温度极低的条件下,生物结皮的发育特征与干热的沙漠地区是否一样,目前还不得而知.生物结皮作为地表特殊的结构层,其属于变水生物[21],在湿润时才能够进行新陈代谢活动[22].而青藏高原高黄河源区属于高原亚寒带半干旱区,仅有冷暖两季,冷季漫长,从每年10月初至次年4月之间地表处于冻结状态[23],降水主要以固态降雪为主,生物结皮呈现休眠状态.在半干旱低温环境条件下,生物结皮的理化性状如何,这一问题也尚不明晰. ...
... 生物结皮对土壤理化属性具有显著影响[1,3,14-15,28].关于生物结皮对土壤颗粒组成的影响,在不同的研究区域所得的研究结论存在一定分歧.有些研究结果表明,生物结皮的存在,能够细化土壤颗粒[28,34-35].也有研究表明,生物结皮对土壤颗粒组成并没有显著影响[36].本研究结果表明,由于生物结皮的发育可增加土壤中粉粒含量,5~20 cm土层中粉粒表现为深色藻结皮>浅色藻结皮>裸地,但差异并不显著.来自沙区的研究结果表明,生物结皮能够黏化土壤,捕捉土壤中细颗粒,使得表层土壤中黏粒和粉粒含量呈显著增加趋势.在黄土区的研究结果也表明,随着生物结皮的发育,结皮层土壤细颗相对增加[28].这些研究结果均表明生物结皮对土壤颗粒组成的影响集中在结皮层及其下0~5 cm左右,细颗粒呈现明显的表聚现象[28].而本研究结果发现由于结皮层的覆盖土壤粉粒在20~30 cm土层显著增加,这可能与该区域强烈的冻融过程及水热迁移有关.由于该区域早晚、四季之间温差较大,呈现明显的冻融循环作用[24-25],因此生物结皮层细颗粒可能由于冻融作用及水分迁移随之向下移动,聚集在其下20~30 cm土层.这种迁移作用可能随着水分含量以及发育年限的不同而发生相应的变化.与此同时,本研究结果表明,冻土区生物结皮层土壤容重也显著低于下层土壤,同时深色藻结皮不仅显著降低了自身容重,也影响到了其下0~20 cm土层土壤容重.而浅色藻结皮则对下层土壤容重影响不明显.本研究区海拔高于4 200 m,海拔高、风力大,与沙漠区域不同,生物结皮在形成过程中,并没有显著的捕捉细颗粒的作用,使结皮层土壤细化从而降低结皮层土壤容重.生物结皮在形成过程中,通过菌丝体、藻丝、假根或分泌的多糖来捆绑、黏结土壤颗粒,使得土壤形成许多团聚结构,土壤中孔隙增大,土壤容重降低[34],而深色藻结皮发育年限较长,厚度较厚,其孔隙度高,因此,容重较浅色藻结皮小,田间持水量显著高于浅色藻结皮及裸地. ...
生物土壤结皮的生态功能
3
2002
... 生物结皮是由生长在表层土壤的微生物、藻类、苔藓和地衣等个体微小的生物成分与土壤相互作用形成的复合层[1].其广泛分布于干旱、半干旱区和极地、亚极地等脆弱生态区[2-3].近年来,关于生物结皮生态功能的研究已详细展开,研究表明生物结皮的形成可以改善土壤理化性质,提高土壤稳定性,改善土壤肥力等,对干旱半干旱地区植被恢复与重建具有重要的生态学意义[3-4].目前关于生物结皮生态功能的研究已有很多,但这些研究主要集中在干旱半干旱的沙漠地区[5-14],例如美国科罗拉多沙漠及其西南部的莫哈维沙漠[6-7],以色列南部的内盖夫沙漠[8],阿根廷的蒙特沙漠[9]以及干旱半干旱的西班牙地中海地区[10].国内关于生物结皮的研究也主要集中在沙漠及干旱半干旱地区,如黄土丘陵沟壑区[11-12]、腾格尔沙漠地区[13-14]、西北及新疆沙漠区[15-16].我国青藏高原素有“世界屋脊”、“第三极”之称,其海拔高,气温低,辐射强烈,日照多,黄河源区也源自于此[17].黄河源区是我国青藏高原重要的水源涵养区,也是该区域最主要的畜牧业基地之一.源区内独特的气候条件及其丰富的自然生态资源为该区经济发展及生态平衡发挥了生态安全屏障的作用[17-18].然而,20世纪70年代以来,随着气候的逐渐干旱化和人口、牲畜数量的迅速增加,该区生态环境急剧恶化,大面积植被退化,20世纪末,植被退化面积占源区草地总面积1/3[19].2005年以来,随着三江源地区实施草地“生态保护和建设”过程的具体实施(围栏封育、退牧还草、沙漠化治理、湿地保护等)[19-20],生物结皮开始在该地区逐渐发育.笔者在野外实地考察过程中发现,生物结皮在黄河源区不同退化程度的草地斑块中均有较大面积的分布.黄河源区平均气温-5.38~4.14 ℃,年降水量262.2~772.8 mm[20],在如此相对湿润且温度极低的条件下,生物结皮的发育特征与干热的沙漠地区是否一样,目前还不得而知.生物结皮作为地表特殊的结构层,其属于变水生物[21],在湿润时才能够进行新陈代谢活动[22].而青藏高原高黄河源区属于高原亚寒带半干旱区,仅有冷暖两季,冷季漫长,从每年10月初至次年4月之间地表处于冻结状态[23],降水主要以固态降雪为主,生物结皮呈现休眠状态.在半干旱低温环境条件下,生物结皮的理化性状如何,这一问题也尚不明晰. ...
... [3-4].目前关于生物结皮生态功能的研究已有很多,但这些研究主要集中在干旱半干旱的沙漠地区[5-14],例如美国科罗拉多沙漠及其西南部的莫哈维沙漠[6-7],以色列南部的内盖夫沙漠[8],阿根廷的蒙特沙漠[9]以及干旱半干旱的西班牙地中海地区[10].国内关于生物结皮的研究也主要集中在沙漠及干旱半干旱地区,如黄土丘陵沟壑区[11-12]、腾格尔沙漠地区[13-14]、西北及新疆沙漠区[15-16].我国青藏高原素有“世界屋脊”、“第三极”之称,其海拔高,气温低,辐射强烈,日照多,黄河源区也源自于此[17].黄河源区是我国青藏高原重要的水源涵养区,也是该区域最主要的畜牧业基地之一.源区内独特的气候条件及其丰富的自然生态资源为该区经济发展及生态平衡发挥了生态安全屏障的作用[17-18].然而,20世纪70年代以来,随着气候的逐渐干旱化和人口、牲畜数量的迅速增加,该区生态环境急剧恶化,大面积植被退化,20世纪末,植被退化面积占源区草地总面积1/3[19].2005年以来,随着三江源地区实施草地“生态保护和建设”过程的具体实施(围栏封育、退牧还草、沙漠化治理、湿地保护等)[19-20],生物结皮开始在该地区逐渐发育.笔者在野外实地考察过程中发现,生物结皮在黄河源区不同退化程度的草地斑块中均有较大面积的分布.黄河源区平均气温-5.38~4.14 ℃,年降水量262.2~772.8 mm[20],在如此相对湿润且温度极低的条件下,生物结皮的发育特征与干热的沙漠地区是否一样,目前还不得而知.生物结皮作为地表特殊的结构层,其属于变水生物[21],在湿润时才能够进行新陈代谢活动[22].而青藏高原高黄河源区属于高原亚寒带半干旱区,仅有冷暖两季,冷季漫长,从每年10月初至次年4月之间地表处于冻结状态[23],降水主要以固态降雪为主,生物结皮呈现休眠状态.在半干旱低温环境条件下,生物结皮的理化性状如何,这一问题也尚不明晰. ...
... 生物结皮对土壤理化属性具有显著影响[1,3,14-15,28].关于生物结皮对土壤颗粒组成的影响,在不同的研究区域所得的研究结论存在一定分歧.有些研究结果表明,生物结皮的存在,能够细化土壤颗粒[28,34-35].也有研究表明,生物结皮对土壤颗粒组成并没有显著影响[36].本研究结果表明,由于生物结皮的发育可增加土壤中粉粒含量,5~20 cm土层中粉粒表现为深色藻结皮>浅色藻结皮>裸地,但差异并不显著.来自沙区的研究结果表明,生物结皮能够黏化土壤,捕捉土壤中细颗粒,使得表层土壤中黏粒和粉粒含量呈显著增加趋势.在黄土区的研究结果也表明,随着生物结皮的发育,结皮层土壤细颗相对增加[28].这些研究结果均表明生物结皮对土壤颗粒组成的影响集中在结皮层及其下0~5 cm左右,细颗粒呈现明显的表聚现象[28].而本研究结果发现由于结皮层的覆盖土壤粉粒在20~30 cm土层显著增加,这可能与该区域强烈的冻融过程及水热迁移有关.由于该区域早晚、四季之间温差较大,呈现明显的冻融循环作用[24-25],因此生物结皮层细颗粒可能由于冻融作用及水分迁移随之向下移动,聚集在其下20~30 cm土层.这种迁移作用可能随着水分含量以及发育年限的不同而发生相应的变化.与此同时,本研究结果表明,冻土区生物结皮层土壤容重也显著低于下层土壤,同时深色藻结皮不仅显著降低了自身容重,也影响到了其下0~20 cm土层土壤容重.而浅色藻结皮则对下层土壤容重影响不明显.本研究区海拔高于4 200 m,海拔高、风力大,与沙漠区域不同,生物结皮在形成过程中,并没有显著的捕捉细颗粒的作用,使结皮层土壤细化从而降低结皮层土壤容重.生物结皮在形成过程中,通过菌丝体、藻丝、假根或分泌的多糖来捆绑、黏结土壤颗粒,使得土壤形成许多团聚结构,土壤中孔隙增大,土壤容重降低[34],而深色藻结皮发育年限较长,厚度较厚,其孔隙度高,因此,容重较浅色藻结皮小,田间持水量显著高于浅色藻结皮及裸地. ...
3
2016
... 生物结皮是由生长在表层土壤的微生物、藻类、苔藓和地衣等个体微小的生物成分与土壤相互作用形成的复合层[1].其广泛分布于干旱、半干旱区和极地、亚极地等脆弱生态区[2-3].近年来,关于生物结皮生态功能的研究已详细展开,研究表明生物结皮的形成可以改善土壤理化性质,提高土壤稳定性,改善土壤肥力等,对干旱半干旱地区植被恢复与重建具有重要的生态学意义[3-4].目前关于生物结皮生态功能的研究已有很多,但这些研究主要集中在干旱半干旱的沙漠地区[5-14],例如美国科罗拉多沙漠及其西南部的莫哈维沙漠[6-7],以色列南部的内盖夫沙漠[8],阿根廷的蒙特沙漠[9]以及干旱半干旱的西班牙地中海地区[10].国内关于生物结皮的研究也主要集中在沙漠及干旱半干旱地区,如黄土丘陵沟壑区[11-12]、腾格尔沙漠地区[13-14]、西北及新疆沙漠区[15-16].我国青藏高原素有“世界屋脊”、“第三极”之称,其海拔高,气温低,辐射强烈,日照多,黄河源区也源自于此[17].黄河源区是我国青藏高原重要的水源涵养区,也是该区域最主要的畜牧业基地之一.源区内独特的气候条件及其丰富的自然生态资源为该区经济发展及生态平衡发挥了生态安全屏障的作用[17-18].然而,20世纪70年代以来,随着气候的逐渐干旱化和人口、牲畜数量的迅速增加,该区生态环境急剧恶化,大面积植被退化,20世纪末,植被退化面积占源区草地总面积1/3[19].2005年以来,随着三江源地区实施草地“生态保护和建设”过程的具体实施(围栏封育、退牧还草、沙漠化治理、湿地保护等)[19-20],生物结皮开始在该地区逐渐发育.笔者在野外实地考察过程中发现,生物结皮在黄河源区不同退化程度的草地斑块中均有较大面积的分布.黄河源区平均气温-5.38~4.14 ℃,年降水量262.2~772.8 mm[20],在如此相对湿润且温度极低的条件下,生物结皮的发育特征与干热的沙漠地区是否一样,目前还不得而知.生物结皮作为地表特殊的结构层,其属于变水生物[21],在湿润时才能够进行新陈代谢活动[22].而青藏高原高黄河源区属于高原亚寒带半干旱区,仅有冷暖两季,冷季漫长,从每年10月初至次年4月之间地表处于冻结状态[23],降水主要以固态降雪为主,生物结皮呈现休眠状态.在半干旱低温环境条件下,生物结皮的理化性状如何,这一问题也尚不明晰. ...
... 生物结皮可以在各种气候条件下广泛发育[1],国内外关于生物结皮的研究主要集中在荒漠地区及干旱半干旱的黄土高原等地区[4-16],关于青藏高原高寒冻土区生物结皮的发育特点鲜有报道.本研究结果表明冻土区生物结皮主要是以藻为主的生物结皮,结皮层厚度大约为6~22 mm.其在厚度上大于干旱的荒漠地区藻类结皮厚度(1~5 mm)[16].这与该地区特殊的生态环境条件密切相关,由于冻土区常年温度低,生物结皮活性较弱,呼吸等生理活性缓慢[4,21],其中积累的物质不易分解,因此,生物结皮层厚度较大.同时,生物结皮的厚度与生物结皮的发育年限密切相关[29].因此,该区域植被退化区域生物结皮(浅色藻结皮)较未退化区生物结皮(深色藻结皮)发育年限短,故其深色藻结皮厚度大于浅色藻结生物结皮小. ...
... [4,21],其中积累的物质不易分解,因此,生物结皮层厚度较大.同时,生物结皮的厚度与生物结皮的发育年限密切相关[29].因此,该区域植被退化区域生物结皮(浅色藻结皮)较未退化区生物结皮(深色藻结皮)发育年限短,故其深色藻结皮厚度大于浅色藻结生物结皮小. ...
Impacts of off-road vehicles on nitrogen cycles in biological soil crusts: resistance in different U.S. deserts
1
2002
... 生物结皮是由生长在表层土壤的微生物、藻类、苔藓和地衣等个体微小的生物成分与土壤相互作用形成的复合层[1].其广泛分布于干旱、半干旱区和极地、亚极地等脆弱生态区[2-3].近年来,关于生物结皮生态功能的研究已详细展开,研究表明生物结皮的形成可以改善土壤理化性质,提高土壤稳定性,改善土壤肥力等,对干旱半干旱地区植被恢复与重建具有重要的生态学意义[3-4].目前关于生物结皮生态功能的研究已有很多,但这些研究主要集中在干旱半干旱的沙漠地区[5-14],例如美国科罗拉多沙漠及其西南部的莫哈维沙漠[6-7],以色列南部的内盖夫沙漠[8],阿根廷的蒙特沙漠[9]以及干旱半干旱的西班牙地中海地区[10].国内关于生物结皮的研究也主要集中在沙漠及干旱半干旱地区,如黄土丘陵沟壑区[11-12]、腾格尔沙漠地区[13-14]、西北及新疆沙漠区[15-16].我国青藏高原素有“世界屋脊”、“第三极”之称,其海拔高,气温低,辐射强烈,日照多,黄河源区也源自于此[17].黄河源区是我国青藏高原重要的水源涵养区,也是该区域最主要的畜牧业基地之一.源区内独特的气候条件及其丰富的自然生态资源为该区经济发展及生态平衡发挥了生态安全屏障的作用[17-18].然而,20世纪70年代以来,随着气候的逐渐干旱化和人口、牲畜数量的迅速增加,该区生态环境急剧恶化,大面积植被退化,20世纪末,植被退化面积占源区草地总面积1/3[19].2005年以来,随着三江源地区实施草地“生态保护和建设”过程的具体实施(围栏封育、退牧还草、沙漠化治理、湿地保护等)[19-20],生物结皮开始在该地区逐渐发育.笔者在野外实地考察过程中发现,生物结皮在黄河源区不同退化程度的草地斑块中均有较大面积的分布.黄河源区平均气温-5.38~4.14 ℃,年降水量262.2~772.8 mm[20],在如此相对湿润且温度极低的条件下,生物结皮的发育特征与干热的沙漠地区是否一样,目前还不得而知.生物结皮作为地表特殊的结构层,其属于变水生物[21],在湿润时才能够进行新陈代谢活动[22].而青藏高原高黄河源区属于高原亚寒带半干旱区,仅有冷暖两季,冷季漫长,从每年10月初至次年4月之间地表处于冻结状态[23],降水主要以固态降雪为主,生物结皮呈现休眠状态.在半干旱低温环境条件下,生物结皮的理化性状如何,这一问题也尚不明晰. ...
Nitrogen fixation by biological soil crusts and heterotrpphic bacteria in an intact Mojave Desert ecosystem with elevated CO2 and added soil carbon
2
2003
... 生物结皮是由生长在表层土壤的微生物、藻类、苔藓和地衣等个体微小的生物成分与土壤相互作用形成的复合层[1].其广泛分布于干旱、半干旱区和极地、亚极地等脆弱生态区[2-3].近年来,关于生物结皮生态功能的研究已详细展开,研究表明生物结皮的形成可以改善土壤理化性质,提高土壤稳定性,改善土壤肥力等,对干旱半干旱地区植被恢复与重建具有重要的生态学意义[3-4].目前关于生物结皮生态功能的研究已有很多,但这些研究主要集中在干旱半干旱的沙漠地区[5-14],例如美国科罗拉多沙漠及其西南部的莫哈维沙漠[6-7],以色列南部的内盖夫沙漠[8],阿根廷的蒙特沙漠[9]以及干旱半干旱的西班牙地中海地区[10].国内关于生物结皮的研究也主要集中在沙漠及干旱半干旱地区,如黄土丘陵沟壑区[11-12]、腾格尔沙漠地区[13-14]、西北及新疆沙漠区[15-16].我国青藏高原素有“世界屋脊”、“第三极”之称,其海拔高,气温低,辐射强烈,日照多,黄河源区也源自于此[17].黄河源区是我国青藏高原重要的水源涵养区,也是该区域最主要的畜牧业基地之一.源区内独特的气候条件及其丰富的自然生态资源为该区经济发展及生态平衡发挥了生态安全屏障的作用[17-18].然而,20世纪70年代以来,随着气候的逐渐干旱化和人口、牲畜数量的迅速增加,该区生态环境急剧恶化,大面积植被退化,20世纪末,植被退化面积占源区草地总面积1/3[19].2005年以来,随着三江源地区实施草地“生态保护和建设”过程的具体实施(围栏封育、退牧还草、沙漠化治理、湿地保护等)[19-20],生物结皮开始在该地区逐渐发育.笔者在野外实地考察过程中发现,生物结皮在黄河源区不同退化程度的草地斑块中均有较大面积的分布.黄河源区平均气温-5.38~4.14 ℃,年降水量262.2~772.8 mm[20],在如此相对湿润且温度极低的条件下,生物结皮的发育特征与干热的沙漠地区是否一样,目前还不得而知.生物结皮作为地表特殊的结构层,其属于变水生物[21],在湿润时才能够进行新陈代谢活动[22].而青藏高原高黄河源区属于高原亚寒带半干旱区,仅有冷暖两季,冷季漫长,从每年10月初至次年4月之间地表处于冻结状态[23],降水主要以固态降雪为主,生物结皮呈现休眠状态.在半干旱低温环境条件下,生物结皮的理化性状如何,这一问题也尚不明晰. ...
... 生物结皮的形成,不仅可以改善土壤结构[28,37],也可显著影响表层土壤的养分累积[9,11,15,21].生物结皮中的固氮蓝藻的存在,可将大气中的氮固定从而输入到表层土壤中[38].同时生物结皮中的微生物能够进行固碳作用,可以增加表层土壤有机质含量[1,14,28],增加土壤肥力.虽然生物结皮对土壤养分积累效应已经形成一致结论,但其作用程度在不同区域还有所不同[14,28,40].本研究结果表明该区两种类型生物结皮均显著增加了结皮层及其下0~20 cm土层土壤有机质含量.藻类是生物结皮发育初期的先锋物种,它的形成能够向体外分泌以多糖为主的物质,从而形成生物结皮[41].同时,在给予充足的水分和光照条件下,生物结皮能够进行固碳作用,其固碳潜力主要取决于生物结皮的生物组成,研究已表明藓结皮的固碳能力高于藻结皮[42].该区域两种类型生物结皮,均以藻类结皮为主,因此其固碳能力可能没有显著差异.同时死亡的藻体又成为土壤有机物,为其他异氧微生物提供碳源[1],因此两种类型生物结皮均依靠显著增加了结皮层及其下0~20 cm土层土壤有机质含量.然而,两种生物结皮对土壤全氮含量的影响则有所不同.深色藻结皮的形成同时也提高了结皮层及其下0~20 cm土层土壤全氮含量,浅色藻结皮则仅提高了结皮层土层土壤全氮含量,对其下0~20 cm土层土壤全氮含量没有显著影响.生物结皮有机体在湿润状态下才能够进行生理代谢活动,进行相应的固氮作用,并且不同的物种固氮所需最佳水温条件不同[6].生物结皮在湿润后,其固氮活性强烈受制于水分[43].以上研究也表明,深色藻结皮的结皮层田间持水量显著高于浅色藻结皮,因此其固氮持续时间可能较长,故其固氮作用较强,向土壤中输入的氮素较多.氮素在水热交换作用下,向下层输入.而浅色藻结皮持水量较低,固氮作用较弱,因此其固定的氮素仅仅存留在结皮层.同时,生物结皮的存在降低了结皮层pH,但其对结皮层及下层土壤pH并没有显著影响.由于生物结皮某些藻类和地衣能够在湿润状态下进行生理活动,释放有机酸,使土壤pH降低[43],青藏高原黄河源区这一高寒地区气温常年低下,并且降水多以固态或暴雨的形式[44],并且在低温条件下,生物结皮有机体产生的有机酸较少也不易挥发释放出来,因此对土壤pH影响较小.高寒地区生物结皮对土壤养分影响过程及机理还需进一步研究. ...
Photosynthesis of crypto biotic soil crusts in a seasonally inundated system of pans and dunes in the western Mojave Desert, CA: Field studies
1
2005
... 生物结皮是由生长在表层土壤的微生物、藻类、苔藓和地衣等个体微小的生物成分与土壤相互作用形成的复合层[1].其广泛分布于干旱、半干旱区和极地、亚极地等脆弱生态区[2-3].近年来,关于生物结皮生态功能的研究已详细展开,研究表明生物结皮的形成可以改善土壤理化性质,提高土壤稳定性,改善土壤肥力等,对干旱半干旱地区植被恢复与重建具有重要的生态学意义[3-4].目前关于生物结皮生态功能的研究已有很多,但这些研究主要集中在干旱半干旱的沙漠地区[5-14],例如美国科罗拉多沙漠及其西南部的莫哈维沙漠[6-7],以色列南部的内盖夫沙漠[8],阿根廷的蒙特沙漠[9]以及干旱半干旱的西班牙地中海地区[10].国内关于生物结皮的研究也主要集中在沙漠及干旱半干旱地区,如黄土丘陵沟壑区[11-12]、腾格尔沙漠地区[13-14]、西北及新疆沙漠区[15-16].我国青藏高原素有“世界屋脊”、“第三极”之称,其海拔高,气温低,辐射强烈,日照多,黄河源区也源自于此[17].黄河源区是我国青藏高原重要的水源涵养区,也是该区域最主要的畜牧业基地之一.源区内独特的气候条件及其丰富的自然生态资源为该区经济发展及生态平衡发挥了生态安全屏障的作用[17-18].然而,20世纪70年代以来,随着气候的逐渐干旱化和人口、牲畜数量的迅速增加,该区生态环境急剧恶化,大面积植被退化,20世纪末,植被退化面积占源区草地总面积1/3[19].2005年以来,随着三江源地区实施草地“生态保护和建设”过程的具体实施(围栏封育、退牧还草、沙漠化治理、湿地保护等)[19-20],生物结皮开始在该地区逐渐发育.笔者在野外实地考察过程中发现,生物结皮在黄河源区不同退化程度的草地斑块中均有较大面积的分布.黄河源区平均气温-5.38~4.14 ℃,年降水量262.2~772.8 mm[20],在如此相对湿润且温度极低的条件下,生物结皮的发育特征与干热的沙漠地区是否一样,目前还不得而知.生物结皮作为地表特殊的结构层,其属于变水生物[21],在湿润时才能够进行新陈代谢活动[22].而青藏高原高黄河源区属于高原亚寒带半干旱区,仅有冷暖两季,冷季漫长,从每年10月初至次年4月之间地表处于冻结状态[23],降水主要以固态降雪为主,生物结皮呈现休眠状态.在半干旱低温环境条件下,生物结皮的理化性状如何,这一问题也尚不明晰. ...
Recovery rates of macrobiotic crusts within a dune ecosystem in the Negev Desert
1
2008
... 生物结皮是由生长在表层土壤的微生物、藻类、苔藓和地衣等个体微小的生物成分与土壤相互作用形成的复合层[1].其广泛分布于干旱、半干旱区和极地、亚极地等脆弱生态区[2-3].近年来,关于生物结皮生态功能的研究已详细展开,研究表明生物结皮的形成可以改善土壤理化性质,提高土壤稳定性,改善土壤肥力等,对干旱半干旱地区植被恢复与重建具有重要的生态学意义[3-4].目前关于生物结皮生态功能的研究已有很多,但这些研究主要集中在干旱半干旱的沙漠地区[5-14],例如美国科罗拉多沙漠及其西南部的莫哈维沙漠[6-7],以色列南部的内盖夫沙漠[8],阿根廷的蒙特沙漠[9]以及干旱半干旱的西班牙地中海地区[10].国内关于生物结皮的研究也主要集中在沙漠及干旱半干旱地区,如黄土丘陵沟壑区[11-12]、腾格尔沙漠地区[13-14]、西北及新疆沙漠区[15-16].我国青藏高原素有“世界屋脊”、“第三极”之称,其海拔高,气温低,辐射强烈,日照多,黄河源区也源自于此[17].黄河源区是我国青藏高原重要的水源涵养区,也是该区域最主要的畜牧业基地之一.源区内独特的气候条件及其丰富的自然生态资源为该区经济发展及生态平衡发挥了生态安全屏障的作用[17-18].然而,20世纪70年代以来,随着气候的逐渐干旱化和人口、牲畜数量的迅速增加,该区生态环境急剧恶化,大面积植被退化,20世纪末,植被退化面积占源区草地总面积1/3[19].2005年以来,随着三江源地区实施草地“生态保护和建设”过程的具体实施(围栏封育、退牧还草、沙漠化治理、湿地保护等)[19-20],生物结皮开始在该地区逐渐发育.笔者在野外实地考察过程中发现,生物结皮在黄河源区不同退化程度的草地斑块中均有较大面积的分布.黄河源区平均气温-5.38~4.14 ℃,年降水量262.2~772.8 mm[20],在如此相对湿润且温度极低的条件下,生物结皮的发育特征与干热的沙漠地区是否一样,目前还不得而知.生物结皮作为地表特殊的结构层,其属于变水生物[21],在湿润时才能够进行新陈代谢活动[22].而青藏高原高黄河源区属于高原亚寒带半干旱区,仅有冷暖两季,冷季漫长,从每年10月初至次年4月之间地表处于冻结状态[23],降水主要以固态降雪为主,生物结皮呈现休眠状态.在半干旱低温环境条件下,生物结皮的理化性状如何,这一问题也尚不明晰. ...
Biological soil crust recovery after long-term grazing exclusion in the Monte Desert (Argentina). Changes in coverage, spatial distribution, and soil nitrogen
2
2012
... 生物结皮是由生长在表层土壤的微生物、藻类、苔藓和地衣等个体微小的生物成分与土壤相互作用形成的复合层[1].其广泛分布于干旱、半干旱区和极地、亚极地等脆弱生态区[2-3].近年来,关于生物结皮生态功能的研究已详细展开,研究表明生物结皮的形成可以改善土壤理化性质,提高土壤稳定性,改善土壤肥力等,对干旱半干旱地区植被恢复与重建具有重要的生态学意义[3-4].目前关于生物结皮生态功能的研究已有很多,但这些研究主要集中在干旱半干旱的沙漠地区[5-14],例如美国科罗拉多沙漠及其西南部的莫哈维沙漠[6-7],以色列南部的内盖夫沙漠[8],阿根廷的蒙特沙漠[9]以及干旱半干旱的西班牙地中海地区[10].国内关于生物结皮的研究也主要集中在沙漠及干旱半干旱地区,如黄土丘陵沟壑区[11-12]、腾格尔沙漠地区[13-14]、西北及新疆沙漠区[15-16].我国青藏高原素有“世界屋脊”、“第三极”之称,其海拔高,气温低,辐射强烈,日照多,黄河源区也源自于此[17].黄河源区是我国青藏高原重要的水源涵养区,也是该区域最主要的畜牧业基地之一.源区内独特的气候条件及其丰富的自然生态资源为该区经济发展及生态平衡发挥了生态安全屏障的作用[17-18].然而,20世纪70年代以来,随着气候的逐渐干旱化和人口、牲畜数量的迅速增加,该区生态环境急剧恶化,大面积植被退化,20世纪末,植被退化面积占源区草地总面积1/3[19].2005年以来,随着三江源地区实施草地“生态保护和建设”过程的具体实施(围栏封育、退牧还草、沙漠化治理、湿地保护等)[19-20],生物结皮开始在该地区逐渐发育.笔者在野外实地考察过程中发现,生物结皮在黄河源区不同退化程度的草地斑块中均有较大面积的分布.黄河源区平均气温-5.38~4.14 ℃,年降水量262.2~772.8 mm[20],在如此相对湿润且温度极低的条件下,生物结皮的发育特征与干热的沙漠地区是否一样,目前还不得而知.生物结皮作为地表特殊的结构层,其属于变水生物[21],在湿润时才能够进行新陈代谢活动[22].而青藏高原高黄河源区属于高原亚寒带半干旱区,仅有冷暖两季,冷季漫长,从每年10月初至次年4月之间地表处于冻结状态[23],降水主要以固态降雪为主,生物结皮呈现休眠状态.在半干旱低温环境条件下,生物结皮的理化性状如何,这一问题也尚不明晰. ...
... 生物结皮的形成,不仅可以改善土壤结构[28,37],也可显著影响表层土壤的养分累积[9,11,15,21].生物结皮中的固氮蓝藻的存在,可将大气中的氮固定从而输入到表层土壤中[38].同时生物结皮中的微生物能够进行固碳作用,可以增加表层土壤有机质含量[1,14,28],增加土壤肥力.虽然生物结皮对土壤养分积累效应已经形成一致结论,但其作用程度在不同区域还有所不同[14,28,40].本研究结果表明该区两种类型生物结皮均显著增加了结皮层及其下0~20 cm土层土壤有机质含量.藻类是生物结皮发育初期的先锋物种,它的形成能够向体外分泌以多糖为主的物质,从而形成生物结皮[41].同时,在给予充足的水分和光照条件下,生物结皮能够进行固碳作用,其固碳潜力主要取决于生物结皮的生物组成,研究已表明藓结皮的固碳能力高于藻结皮[42].该区域两种类型生物结皮,均以藻类结皮为主,因此其固碳能力可能没有显著差异.同时死亡的藻体又成为土壤有机物,为其他异氧微生物提供碳源[1],因此两种类型生物结皮均依靠显著增加了结皮层及其下0~20 cm土层土壤有机质含量.然而,两种生物结皮对土壤全氮含量的影响则有所不同.深色藻结皮的形成同时也提高了结皮层及其下0~20 cm土层土壤全氮含量,浅色藻结皮则仅提高了结皮层土层土壤全氮含量,对其下0~20 cm土层土壤全氮含量没有显著影响.生物结皮有机体在湿润状态下才能够进行生理代谢活动,进行相应的固氮作用,并且不同的物种固氮所需最佳水温条件不同[6].生物结皮在湿润后,其固氮活性强烈受制于水分[43].以上研究也表明,深色藻结皮的结皮层田间持水量显著高于浅色藻结皮,因此其固氮持续时间可能较长,故其固氮作用较强,向土壤中输入的氮素较多.氮素在水热交换作用下,向下层输入.而浅色藻结皮持水量较低,固氮作用较弱,因此其固定的氮素仅仅存留在结皮层.同时,生物结皮的存在降低了结皮层pH,但其对结皮层及下层土壤pH并没有显著影响.由于生物结皮某些藻类和地衣能够在湿润状态下进行生理活动,释放有机酸,使土壤pH降低[43],青藏高原黄河源区这一高寒地区气温常年低下,并且降水多以固态或暴雨的形式[44],并且在低温条件下,生物结皮有机体产生的有机酸较少也不易挥发释放出来,因此对土壤pH影响较小.高寒地区生物结皮对土壤养分影响过程及机理还需进一步研究. ...
Labile carbon in biological soil crusts in the Tabernas desert, SE Spain
1
2013
... 生物结皮是由生长在表层土壤的微生物、藻类、苔藓和地衣等个体微小的生物成分与土壤相互作用形成的复合层[1].其广泛分布于干旱、半干旱区和极地、亚极地等脆弱生态区[2-3].近年来,关于生物结皮生态功能的研究已详细展开,研究表明生物结皮的形成可以改善土壤理化性质,提高土壤稳定性,改善土壤肥力等,对干旱半干旱地区植被恢复与重建具有重要的生态学意义[3-4].目前关于生物结皮生态功能的研究已有很多,但这些研究主要集中在干旱半干旱的沙漠地区[5-14],例如美国科罗拉多沙漠及其西南部的莫哈维沙漠[6-7],以色列南部的内盖夫沙漠[8],阿根廷的蒙特沙漠[9]以及干旱半干旱的西班牙地中海地区[10].国内关于生物结皮的研究也主要集中在沙漠及干旱半干旱地区,如黄土丘陵沟壑区[11-12]、腾格尔沙漠地区[13-14]、西北及新疆沙漠区[15-16].我国青藏高原素有“世界屋脊”、“第三极”之称,其海拔高,气温低,辐射强烈,日照多,黄河源区也源自于此[17].黄河源区是我国青藏高原重要的水源涵养区,也是该区域最主要的畜牧业基地之一.源区内独特的气候条件及其丰富的自然生态资源为该区经济发展及生态平衡发挥了生态安全屏障的作用[17-18].然而,20世纪70年代以来,随着气候的逐渐干旱化和人口、牲畜数量的迅速增加,该区生态环境急剧恶化,大面积植被退化,20世纪末,植被退化面积占源区草地总面积1/3[19].2005年以来,随着三江源地区实施草地“生态保护和建设”过程的具体实施(围栏封育、退牧还草、沙漠化治理、湿地保护等)[19-20],生物结皮开始在该地区逐渐发育.笔者在野外实地考察过程中发现,生物结皮在黄河源区不同退化程度的草地斑块中均有较大面积的分布.黄河源区平均气温-5.38~4.14 ℃,年降水量262.2~772.8 mm[20],在如此相对湿润且温度极低的条件下,生物结皮的发育特征与干热的沙漠地区是否一样,目前还不得而知.生物结皮作为地表特殊的结构层,其属于变水生物[21],在湿润时才能够进行新陈代谢活动[22].而青藏高原高黄河源区属于高原亚寒带半干旱区,仅有冷暖两季,冷季漫长,从每年10月初至次年4月之间地表处于冻结状态[23],降水主要以固态降雪为主,生物结皮呈现休眠状态.在半干旱低温环境条件下,生物结皮的理化性状如何,这一问题也尚不明晰. ...
Impact of biological soil crust on soil physical and chemical properties of rehabilitated grassland in Hilly Loess Plateau, China
2
2006
... 生物结皮是由生长在表层土壤的微生物、藻类、苔藓和地衣等个体微小的生物成分与土壤相互作用形成的复合层[1].其广泛分布于干旱、半干旱区和极地、亚极地等脆弱生态区[2-3].近年来,关于生物结皮生态功能的研究已详细展开,研究表明生物结皮的形成可以改善土壤理化性质,提高土壤稳定性,改善土壤肥力等,对干旱半干旱地区植被恢复与重建具有重要的生态学意义[3-4].目前关于生物结皮生态功能的研究已有很多,但这些研究主要集中在干旱半干旱的沙漠地区[5-14],例如美国科罗拉多沙漠及其西南部的莫哈维沙漠[6-7],以色列南部的内盖夫沙漠[8],阿根廷的蒙特沙漠[9]以及干旱半干旱的西班牙地中海地区[10].国内关于生物结皮的研究也主要集中在沙漠及干旱半干旱地区,如黄土丘陵沟壑区[11-12]、腾格尔沙漠地区[13-14]、西北及新疆沙漠区[15-16].我国青藏高原素有“世界屋脊”、“第三极”之称,其海拔高,气温低,辐射强烈,日照多,黄河源区也源自于此[17].黄河源区是我国青藏高原重要的水源涵养区,也是该区域最主要的畜牧业基地之一.源区内独特的气候条件及其丰富的自然生态资源为该区经济发展及生态平衡发挥了生态安全屏障的作用[17-18].然而,20世纪70年代以来,随着气候的逐渐干旱化和人口、牲畜数量的迅速增加,该区生态环境急剧恶化,大面积植被退化,20世纪末,植被退化面积占源区草地总面积1/3[19].2005年以来,随着三江源地区实施草地“生态保护和建设”过程的具体实施(围栏封育、退牧还草、沙漠化治理、湿地保护等)[19-20],生物结皮开始在该地区逐渐发育.笔者在野外实地考察过程中发现,生物结皮在黄河源区不同退化程度的草地斑块中均有较大面积的分布.黄河源区平均气温-5.38~4.14 ℃,年降水量262.2~772.8 mm[20],在如此相对湿润且温度极低的条件下,生物结皮的发育特征与干热的沙漠地区是否一样,目前还不得而知.生物结皮作为地表特殊的结构层,其属于变水生物[21],在湿润时才能够进行新陈代谢活动[22].而青藏高原高黄河源区属于高原亚寒带半干旱区,仅有冷暖两季,冷季漫长,从每年10月初至次年4月之间地表处于冻结状态[23],降水主要以固态降雪为主,生物结皮呈现休眠状态.在半干旱低温环境条件下,生物结皮的理化性状如何,这一问题也尚不明晰. ...
... 生物结皮的形成,不仅可以改善土壤结构[28,37],也可显著影响表层土壤的养分累积[9,11,15,21].生物结皮中的固氮蓝藻的存在,可将大气中的氮固定从而输入到表层土壤中[38].同时生物结皮中的微生物能够进行固碳作用,可以增加表层土壤有机质含量[1,14,28],增加土壤肥力.虽然生物结皮对土壤养分积累效应已经形成一致结论,但其作用程度在不同区域还有所不同[14,28,40].本研究结果表明该区两种类型生物结皮均显著增加了结皮层及其下0~20 cm土层土壤有机质含量.藻类是生物结皮发育初期的先锋物种,它的形成能够向体外分泌以多糖为主的物质,从而形成生物结皮[41].同时,在给予充足的水分和光照条件下,生物结皮能够进行固碳作用,其固碳潜力主要取决于生物结皮的生物组成,研究已表明藓结皮的固碳能力高于藻结皮[42].该区域两种类型生物结皮,均以藻类结皮为主,因此其固碳能力可能没有显著差异.同时死亡的藻体又成为土壤有机物,为其他异氧微生物提供碳源[1],因此两种类型生物结皮均依靠显著增加了结皮层及其下0~20 cm土层土壤有机质含量.然而,两种生物结皮对土壤全氮含量的影响则有所不同.深色藻结皮的形成同时也提高了结皮层及其下0~20 cm土层土壤全氮含量,浅色藻结皮则仅提高了结皮层土层土壤全氮含量,对其下0~20 cm土层土壤全氮含量没有显著影响.生物结皮有机体在湿润状态下才能够进行生理代谢活动,进行相应的固氮作用,并且不同的物种固氮所需最佳水温条件不同[6].生物结皮在湿润后,其固氮活性强烈受制于水分[43].以上研究也表明,深色藻结皮的结皮层田间持水量显著高于浅色藻结皮,因此其固氮持续时间可能较长,故其固氮作用较强,向土壤中输入的氮素较多.氮素在水热交换作用下,向下层输入.而浅色藻结皮持水量较低,固氮作用较弱,因此其固定的氮素仅仅存留在结皮层.同时,生物结皮的存在降低了结皮层pH,但其对结皮层及下层土壤pH并没有显著影响.由于生物结皮某些藻类和地衣能够在湿润状态下进行生理活动,释放有机酸,使土壤pH降低[43],青藏高原黄河源区这一高寒地区气温常年低下,并且降水多以固态或暴雨的形式[44],并且在低温条件下,生物结皮有机体产生的有机酸较少也不易挥发释放出来,因此对土壤pH影响较小.高寒地区生物结皮对土壤养分影响过程及机理还需进一步研究. ...
黄土丘陵区退耕地生物结皮对土壤理化性状的影响
2
2006
... 生物结皮是由生长在表层土壤的微生物、藻类、苔藓和地衣等个体微小的生物成分与土壤相互作用形成的复合层[1].其广泛分布于干旱、半干旱区和极地、亚极地等脆弱生态区[2-3].近年来,关于生物结皮生态功能的研究已详细展开,研究表明生物结皮的形成可以改善土壤理化性质,提高土壤稳定性,改善土壤肥力等,对干旱半干旱地区植被恢复与重建具有重要的生态学意义[3-4].目前关于生物结皮生态功能的研究已有很多,但这些研究主要集中在干旱半干旱的沙漠地区[5-14],例如美国科罗拉多沙漠及其西南部的莫哈维沙漠[6-7],以色列南部的内盖夫沙漠[8],阿根廷的蒙特沙漠[9]以及干旱半干旱的西班牙地中海地区[10].国内关于生物结皮的研究也主要集中在沙漠及干旱半干旱地区,如黄土丘陵沟壑区[11-12]、腾格尔沙漠地区[13-14]、西北及新疆沙漠区[15-16].我国青藏高原素有“世界屋脊”、“第三极”之称,其海拔高,气温低,辐射强烈,日照多,黄河源区也源自于此[17].黄河源区是我国青藏高原重要的水源涵养区,也是该区域最主要的畜牧业基地之一.源区内独特的气候条件及其丰富的自然生态资源为该区经济发展及生态平衡发挥了生态安全屏障的作用[17-18].然而,20世纪70年代以来,随着气候的逐渐干旱化和人口、牲畜数量的迅速增加,该区生态环境急剧恶化,大面积植被退化,20世纪末,植被退化面积占源区草地总面积1/3[19].2005年以来,随着三江源地区实施草地“生态保护和建设”过程的具体实施(围栏封育、退牧还草、沙漠化治理、湿地保护等)[19-20],生物结皮开始在该地区逐渐发育.笔者在野外实地考察过程中发现,生物结皮在黄河源区不同退化程度的草地斑块中均有较大面积的分布.黄河源区平均气温-5.38~4.14 ℃,年降水量262.2~772.8 mm[20],在如此相对湿润且温度极低的条件下,生物结皮的发育特征与干热的沙漠地区是否一样,目前还不得而知.生物结皮作为地表特殊的结构层,其属于变水生物[21],在湿润时才能够进行新陈代谢活动[22].而青藏高原高黄河源区属于高原亚寒带半干旱区,仅有冷暖两季,冷季漫长,从每年10月初至次年4月之间地表处于冻结状态[23],降水主要以固态降雪为主,生物结皮呈现休眠状态.在半干旱低温环境条件下,生物结皮的理化性状如何,这一问题也尚不明晰. ...
... 生物结皮的形成,不仅可以改善土壤结构[28,37],也可显著影响表层土壤的养分累积[9,11,15,21].生物结皮中的固氮蓝藻的存在,可将大气中的氮固定从而输入到表层土壤中[38].同时生物结皮中的微生物能够进行固碳作用,可以增加表层土壤有机质含量[1,14,28],增加土壤肥力.虽然生物结皮对土壤养分积累效应已经形成一致结论,但其作用程度在不同区域还有所不同[14,28,40].本研究结果表明该区两种类型生物结皮均显著增加了结皮层及其下0~20 cm土层土壤有机质含量.藻类是生物结皮发育初期的先锋物种,它的形成能够向体外分泌以多糖为主的物质,从而形成生物结皮[41].同时,在给予充足的水分和光照条件下,生物结皮能够进行固碳作用,其固碳潜力主要取决于生物结皮的生物组成,研究已表明藓结皮的固碳能力高于藻结皮[42].该区域两种类型生物结皮,均以藻类结皮为主,因此其固碳能力可能没有显著差异.同时死亡的藻体又成为土壤有机物,为其他异氧微生物提供碳源[1],因此两种类型生物结皮均依靠显著增加了结皮层及其下0~20 cm土层土壤有机质含量.然而,两种生物结皮对土壤全氮含量的影响则有所不同.深色藻结皮的形成同时也提高了结皮层及其下0~20 cm土层土壤全氮含量,浅色藻结皮则仅提高了结皮层土层土壤全氮含量,对其下0~20 cm土层土壤全氮含量没有显著影响.生物结皮有机体在湿润状态下才能够进行生理代谢活动,进行相应的固氮作用,并且不同的物种固氮所需最佳水温条件不同[6].生物结皮在湿润后,其固氮活性强烈受制于水分[43].以上研究也表明,深色藻结皮的结皮层田间持水量显著高于浅色藻结皮,因此其固氮持续时间可能较长,故其固氮作用较强,向土壤中输入的氮素较多.氮素在水热交换作用下,向下层输入.而浅色藻结皮持水量较低,固氮作用较弱,因此其固定的氮素仅仅存留在结皮层.同时,生物结皮的存在降低了结皮层pH,但其对结皮层及下层土壤pH并没有显著影响.由于生物结皮某些藻类和地衣能够在湿润状态下进行生理活动,释放有机酸,使土壤pH降低[43],青藏高原黄河源区这一高寒地区气温常年低下,并且降水多以固态或暴雨的形式[44],并且在低温条件下,生物结皮有机体产生的有机酸较少也不易挥发释放出来,因此对土壤pH影响较小.高寒地区生物结皮对土壤养分影响过程及机理还需进一步研究. ...
The effects of biological soil crust succession on soil ecological stoichiometry characteristics
1
2018
... 生物结皮是由生长在表层土壤的微生物、藻类、苔藓和地衣等个体微小的生物成分与土壤相互作用形成的复合层[1].其广泛分布于干旱、半干旱区和极地、亚极地等脆弱生态区[2-3].近年来,关于生物结皮生态功能的研究已详细展开,研究表明生物结皮的形成可以改善土壤理化性质,提高土壤稳定性,改善土壤肥力等,对干旱半干旱地区植被恢复与重建具有重要的生态学意义[3-4].目前关于生物结皮生态功能的研究已有很多,但这些研究主要集中在干旱半干旱的沙漠地区[5-14],例如美国科罗拉多沙漠及其西南部的莫哈维沙漠[6-7],以色列南部的内盖夫沙漠[8],阿根廷的蒙特沙漠[9]以及干旱半干旱的西班牙地中海地区[10].国内关于生物结皮的研究也主要集中在沙漠及干旱半干旱地区,如黄土丘陵沟壑区[11-12]、腾格尔沙漠地区[13-14]、西北及新疆沙漠区[15-16].我国青藏高原素有“世界屋脊”、“第三极”之称,其海拔高,气温低,辐射强烈,日照多,黄河源区也源自于此[17].黄河源区是我国青藏高原重要的水源涵养区,也是该区域最主要的畜牧业基地之一.源区内独特的气候条件及其丰富的自然生态资源为该区经济发展及生态平衡发挥了生态安全屏障的作用[17-18].然而,20世纪70年代以来,随着气候的逐渐干旱化和人口、牲畜数量的迅速增加,该区生态环境急剧恶化,大面积植被退化,20世纪末,植被退化面积占源区草地总面积1/3[19].2005年以来,随着三江源地区实施草地“生态保护和建设”过程的具体实施(围栏封育、退牧还草、沙漠化治理、湿地保护等)[19-20],生物结皮开始在该地区逐渐发育.笔者在野外实地考察过程中发现,生物结皮在黄河源区不同退化程度的草地斑块中均有较大面积的分布.黄河源区平均气温-5.38~4.14 ℃,年降水量262.2~772.8 mm[20],在如此相对湿润且温度极低的条件下,生物结皮的发育特征与干热的沙漠地区是否一样,目前还不得而知.生物结皮作为地表特殊的结构层,其属于变水生物[21],在湿润时才能够进行新陈代谢活动[22].而青藏高原高黄河源区属于高原亚寒带半干旱区,仅有冷暖两季,冷季漫长,从每年10月初至次年4月之间地表处于冻结状态[23],降水主要以固态降雪为主,生物结皮呈现休眠状态.在半干旱低温环境条件下,生物结皮的理化性状如何,这一问题也尚不明晰. ...
生物土壤结皮演替对土壤生态化学计量特征的影响
1
2018
... 生物结皮是由生长在表层土壤的微生物、藻类、苔藓和地衣等个体微小的生物成分与土壤相互作用形成的复合层[1].其广泛分布于干旱、半干旱区和极地、亚极地等脆弱生态区[2-3].近年来,关于生物结皮生态功能的研究已详细展开,研究表明生物结皮的形成可以改善土壤理化性质,提高土壤稳定性,改善土壤肥力等,对干旱半干旱地区植被恢复与重建具有重要的生态学意义[3-4].目前关于生物结皮生态功能的研究已有很多,但这些研究主要集中在干旱半干旱的沙漠地区[5-14],例如美国科罗拉多沙漠及其西南部的莫哈维沙漠[6-7],以色列南部的内盖夫沙漠[8],阿根廷的蒙特沙漠[9]以及干旱半干旱的西班牙地中海地区[10].国内关于生物结皮的研究也主要集中在沙漠及干旱半干旱地区,如黄土丘陵沟壑区[11-12]、腾格尔沙漠地区[13-14]、西北及新疆沙漠区[15-16].我国青藏高原素有“世界屋脊”、“第三极”之称,其海拔高,气温低,辐射强烈,日照多,黄河源区也源自于此[17].黄河源区是我国青藏高原重要的水源涵养区,也是该区域最主要的畜牧业基地之一.源区内独特的气候条件及其丰富的自然生态资源为该区经济发展及生态平衡发挥了生态安全屏障的作用[17-18].然而,20世纪70年代以来,随着气候的逐渐干旱化和人口、牲畜数量的迅速增加,该区生态环境急剧恶化,大面积植被退化,20世纪末,植被退化面积占源区草地总面积1/3[19].2005年以来,随着三江源地区实施草地“生态保护和建设”过程的具体实施(围栏封育、退牧还草、沙漠化治理、湿地保护等)[19-20],生物结皮开始在该地区逐渐发育.笔者在野外实地考察过程中发现,生物结皮在黄河源区不同退化程度的草地斑块中均有较大面积的分布.黄河源区平均气温-5.38~4.14 ℃,年降水量262.2~772.8 mm[20],在如此相对湿润且温度极低的条件下,生物结皮的发育特征与干热的沙漠地区是否一样,目前还不得而知.生物结皮作为地表特殊的结构层,其属于变水生物[21],在湿润时才能够进行新陈代谢活动[22].而青藏高原高黄河源区属于高原亚寒带半干旱区,仅有冷暖两季,冷季漫长,从每年10月初至次年4月之间地表处于冻结状态[23],降水主要以固态降雪为主,生物结皮呈现休眠状态.在半干旱低温环境条件下,生物结皮的理化性状如何,这一问题也尚不明晰. ...
Microbiotic soil crust and its effect on vegetation and habitat on artificially stabilize desert dunes in Tengger Desert, North China
1
2012
... 生物结皮是由生长在表层土壤的微生物、藻类、苔藓和地衣等个体微小的生物成分与土壤相互作用形成的复合层[1].其广泛分布于干旱、半干旱区和极地、亚极地等脆弱生态区[2-3].近年来,关于生物结皮生态功能的研究已详细展开,研究表明生物结皮的形成可以改善土壤理化性质,提高土壤稳定性,改善土壤肥力等,对干旱半干旱地区植被恢复与重建具有重要的生态学意义[3-4].目前关于生物结皮生态功能的研究已有很多,但这些研究主要集中在干旱半干旱的沙漠地区[5-14],例如美国科罗拉多沙漠及其西南部的莫哈维沙漠[6-7],以色列南部的内盖夫沙漠[8],阿根廷的蒙特沙漠[9]以及干旱半干旱的西班牙地中海地区[10].国内关于生物结皮的研究也主要集中在沙漠及干旱半干旱地区,如黄土丘陵沟壑区[11-12]、腾格尔沙漠地区[13-14]、西北及新疆沙漠区[15-16].我国青藏高原素有“世界屋脊”、“第三极”之称,其海拔高,气温低,辐射强烈,日照多,黄河源区也源自于此[17].黄河源区是我国青藏高原重要的水源涵养区,也是该区域最主要的畜牧业基地之一.源区内独特的气候条件及其丰富的自然生态资源为该区经济发展及生态平衡发挥了生态安全屏障的作用[17-18].然而,20世纪70年代以来,随着气候的逐渐干旱化和人口、牲畜数量的迅速增加,该区生态环境急剧恶化,大面积植被退化,20世纪末,植被退化面积占源区草地总面积1/3[19].2005年以来,随着三江源地区实施草地“生态保护和建设”过程的具体实施(围栏封育、退牧还草、沙漠化治理、湿地保护等)[19-20],生物结皮开始在该地区逐渐发育.笔者在野外实地考察过程中发现,生物结皮在黄河源区不同退化程度的草地斑块中均有较大面积的分布.黄河源区平均气温-5.38~4.14 ℃,年降水量262.2~772.8 mm[20],在如此相对湿润且温度极低的条件下,生物结皮的发育特征与干热的沙漠地区是否一样,目前还不得而知.生物结皮作为地表特殊的结构层,其属于变水生物[21],在湿润时才能够进行新陈代谢活动[22].而青藏高原高黄河源区属于高原亚寒带半干旱区,仅有冷暖两季,冷季漫长,从每年10月初至次年4月之间地表处于冻结状态[23],降水主要以固态降雪为主,生物结皮呈现休眠状态.在半干旱低温环境条件下,生物结皮的理化性状如何,这一问题也尚不明晰. ...
Effects of biological soil crusts on seed bank, germination and establishment of two annual plant species in the Tengger Desert (N China)
5
2005
... 生物结皮是由生长在表层土壤的微生物、藻类、苔藓和地衣等个体微小的生物成分与土壤相互作用形成的复合层[1].其广泛分布于干旱、半干旱区和极地、亚极地等脆弱生态区[2-3].近年来,关于生物结皮生态功能的研究已详细展开,研究表明生物结皮的形成可以改善土壤理化性质,提高土壤稳定性,改善土壤肥力等,对干旱半干旱地区植被恢复与重建具有重要的生态学意义[3-4].目前关于生物结皮生态功能的研究已有很多,但这些研究主要集中在干旱半干旱的沙漠地区[5-14],例如美国科罗拉多沙漠及其西南部的莫哈维沙漠[6-7],以色列南部的内盖夫沙漠[8],阿根廷的蒙特沙漠[9]以及干旱半干旱的西班牙地中海地区[10].国内关于生物结皮的研究也主要集中在沙漠及干旱半干旱地区,如黄土丘陵沟壑区[11-12]、腾格尔沙漠地区[13-14]、西北及新疆沙漠区[15-16].我国青藏高原素有“世界屋脊”、“第三极”之称,其海拔高,气温低,辐射强烈,日照多,黄河源区也源自于此[17].黄河源区是我国青藏高原重要的水源涵养区,也是该区域最主要的畜牧业基地之一.源区内独特的气候条件及其丰富的自然生态资源为该区经济发展及生态平衡发挥了生态安全屏障的作用[17-18].然而,20世纪70年代以来,随着气候的逐渐干旱化和人口、牲畜数量的迅速增加,该区生态环境急剧恶化,大面积植被退化,20世纪末,植被退化面积占源区草地总面积1/3[19].2005年以来,随着三江源地区实施草地“生态保护和建设”过程的具体实施(围栏封育、退牧还草、沙漠化治理、湿地保护等)[19-20],生物结皮开始在该地区逐渐发育.笔者在野外实地考察过程中发现,生物结皮在黄河源区不同退化程度的草地斑块中均有较大面积的分布.黄河源区平均气温-5.38~4.14 ℃,年降水量262.2~772.8 mm[20],在如此相对湿润且温度极低的条件下,生物结皮的发育特征与干热的沙漠地区是否一样,目前还不得而知.生物结皮作为地表特殊的结构层,其属于变水生物[21],在湿润时才能够进行新陈代谢活动[22].而青藏高原高黄河源区属于高原亚寒带半干旱区,仅有冷暖两季,冷季漫长,从每年10月初至次年4月之间地表处于冻结状态[23],降水主要以固态降雪为主,生物结皮呈现休眠状态.在半干旱低温环境条件下,生物结皮的理化性状如何,这一问题也尚不明晰. ...
... -14]、西北及新疆沙漠区[15-16].我国青藏高原素有“世界屋脊”、“第三极”之称,其海拔高,气温低,辐射强烈,日照多,黄河源区也源自于此[17].黄河源区是我国青藏高原重要的水源涵养区,也是该区域最主要的畜牧业基地之一.源区内独特的气候条件及其丰富的自然生态资源为该区经济发展及生态平衡发挥了生态安全屏障的作用[17-18].然而,20世纪70年代以来,随着气候的逐渐干旱化和人口、牲畜数量的迅速增加,该区生态环境急剧恶化,大面积植被退化,20世纪末,植被退化面积占源区草地总面积1/3[19].2005年以来,随着三江源地区实施草地“生态保护和建设”过程的具体实施(围栏封育、退牧还草、沙漠化治理、湿地保护等)[19-20],生物结皮开始在该地区逐渐发育.笔者在野外实地考察过程中发现,生物结皮在黄河源区不同退化程度的草地斑块中均有较大面积的分布.黄河源区平均气温-5.38~4.14 ℃,年降水量262.2~772.8 mm[20],在如此相对湿润且温度极低的条件下,生物结皮的发育特征与干热的沙漠地区是否一样,目前还不得而知.生物结皮作为地表特殊的结构层,其属于变水生物[21],在湿润时才能够进行新陈代谢活动[22].而青藏高原高黄河源区属于高原亚寒带半干旱区,仅有冷暖两季,冷季漫长,从每年10月初至次年4月之间地表处于冻结状态[23],降水主要以固态降雪为主,生物结皮呈现休眠状态.在半干旱低温环境条件下,生物结皮的理化性状如何,这一问题也尚不明晰. ...
... 生物结皮对土壤理化属性具有显著影响[1,3,14-15,28].关于生物结皮对土壤颗粒组成的影响,在不同的研究区域所得的研究结论存在一定分歧.有些研究结果表明,生物结皮的存在,能够细化土壤颗粒[28,34-35].也有研究表明,生物结皮对土壤颗粒组成并没有显著影响[36].本研究结果表明,由于生物结皮的发育可增加土壤中粉粒含量,5~20 cm土层中粉粒表现为深色藻结皮>浅色藻结皮>裸地,但差异并不显著.来自沙区的研究结果表明,生物结皮能够黏化土壤,捕捉土壤中细颗粒,使得表层土壤中黏粒和粉粒含量呈显著增加趋势.在黄土区的研究结果也表明,随着生物结皮的发育,结皮层土壤细颗相对增加[28].这些研究结果均表明生物结皮对土壤颗粒组成的影响集中在结皮层及其下0~5 cm左右,细颗粒呈现明显的表聚现象[28].而本研究结果发现由于结皮层的覆盖土壤粉粒在20~30 cm土层显著增加,这可能与该区域强烈的冻融过程及水热迁移有关.由于该区域早晚、四季之间温差较大,呈现明显的冻融循环作用[24-25],因此生物结皮层细颗粒可能由于冻融作用及水分迁移随之向下移动,聚集在其下20~30 cm土层.这种迁移作用可能随着水分含量以及发育年限的不同而发生相应的变化.与此同时,本研究结果表明,冻土区生物结皮层土壤容重也显著低于下层土壤,同时深色藻结皮不仅显著降低了自身容重,也影响到了其下0~20 cm土层土壤容重.而浅色藻结皮则对下层土壤容重影响不明显.本研究区海拔高于4 200 m,海拔高、风力大,与沙漠区域不同,生物结皮在形成过程中,并没有显著的捕捉细颗粒的作用,使结皮层土壤细化从而降低结皮层土壤容重.生物结皮在形成过程中,通过菌丝体、藻丝、假根或分泌的多糖来捆绑、黏结土壤颗粒,使得土壤形成许多团聚结构,土壤中孔隙增大,土壤容重降低[34],而深色藻结皮发育年限较长,厚度较厚,其孔隙度高,因此,容重较浅色藻结皮小,田间持水量显著高于浅色藻结皮及裸地. ...
... 生物结皮的形成,不仅可以改善土壤结构[28,37],也可显著影响表层土壤的养分累积[9,11,15,21].生物结皮中的固氮蓝藻的存在,可将大气中的氮固定从而输入到表层土壤中[38].同时生物结皮中的微生物能够进行固碳作用,可以增加表层土壤有机质含量[1,14,28],增加土壤肥力.虽然生物结皮对土壤养分积累效应已经形成一致结论,但其作用程度在不同区域还有所不同[14,28,40].本研究结果表明该区两种类型生物结皮均显著增加了结皮层及其下0~20 cm土层土壤有机质含量.藻类是生物结皮发育初期的先锋物种,它的形成能够向体外分泌以多糖为主的物质,从而形成生物结皮[41].同时,在给予充足的水分和光照条件下,生物结皮能够进行固碳作用,其固碳潜力主要取决于生物结皮的生物组成,研究已表明藓结皮的固碳能力高于藻结皮[42].该区域两种类型生物结皮,均以藻类结皮为主,因此其固碳能力可能没有显著差异.同时死亡的藻体又成为土壤有机物,为其他异氧微生物提供碳源[1],因此两种类型生物结皮均依靠显著增加了结皮层及其下0~20 cm土层土壤有机质含量.然而,两种生物结皮对土壤全氮含量的影响则有所不同.深色藻结皮的形成同时也提高了结皮层及其下0~20 cm土层土壤全氮含量,浅色藻结皮则仅提高了结皮层土层土壤全氮含量,对其下0~20 cm土层土壤全氮含量没有显著影响.生物结皮有机体在湿润状态下才能够进行生理代谢活动,进行相应的固氮作用,并且不同的物种固氮所需最佳水温条件不同[6].生物结皮在湿润后,其固氮活性强烈受制于水分[43].以上研究也表明,深色藻结皮的结皮层田间持水量显著高于浅色藻结皮,因此其固氮持续时间可能较长,故其固氮作用较强,向土壤中输入的氮素较多.氮素在水热交换作用下,向下层输入.而浅色藻结皮持水量较低,固氮作用较弱,因此其固定的氮素仅仅存留在结皮层.同时,生物结皮的存在降低了结皮层pH,但其对结皮层及下层土壤pH并没有显著影响.由于生物结皮某些藻类和地衣能够在湿润状态下进行生理活动,释放有机酸,使土壤pH降低[43],青藏高原黄河源区这一高寒地区气温常年低下,并且降水多以固态或暴雨的形式[44],并且在低温条件下,生物结皮有机体产生的有机酸较少也不易挥发释放出来,因此对土壤pH影响较小.高寒地区生物结皮对土壤养分影响过程及机理还需进一步研究. ...
... [14,28,40].本研究结果表明该区两种类型生物结皮均显著增加了结皮层及其下0~20 cm土层土壤有机质含量.藻类是生物结皮发育初期的先锋物种,它的形成能够向体外分泌以多糖为主的物质,从而形成生物结皮[41].同时,在给予充足的水分和光照条件下,生物结皮能够进行固碳作用,其固碳潜力主要取决于生物结皮的生物组成,研究已表明藓结皮的固碳能力高于藻结皮[42].该区域两种类型生物结皮,均以藻类结皮为主,因此其固碳能力可能没有显著差异.同时死亡的藻体又成为土壤有机物,为其他异氧微生物提供碳源[1],因此两种类型生物结皮均依靠显著增加了结皮层及其下0~20 cm土层土壤有机质含量.然而,两种生物结皮对土壤全氮含量的影响则有所不同.深色藻结皮的形成同时也提高了结皮层及其下0~20 cm土层土壤全氮含量,浅色藻结皮则仅提高了结皮层土层土壤全氮含量,对其下0~20 cm土层土壤全氮含量没有显著影响.生物结皮有机体在湿润状态下才能够进行生理代谢活动,进行相应的固氮作用,并且不同的物种固氮所需最佳水温条件不同[6].生物结皮在湿润后,其固氮活性强烈受制于水分[43].以上研究也表明,深色藻结皮的结皮层田间持水量显著高于浅色藻结皮,因此其固氮持续时间可能较长,故其固氮作用较强,向土壤中输入的氮素较多.氮素在水热交换作用下,向下层输入.而浅色藻结皮持水量较低,固氮作用较弱,因此其固定的氮素仅仅存留在结皮层.同时,生物结皮的存在降低了结皮层pH,但其对结皮层及下层土壤pH并没有显著影响.由于生物结皮某些藻类和地衣能够在湿润状态下进行生理活动,释放有机酸,使土壤pH降低[43],青藏高原黄河源区这一高寒地区气温常年低下,并且降水多以固态或暴雨的形式[44],并且在低温条件下,生物结皮有机体产生的有机酸较少也不易挥发释放出来,因此对土壤pH影响较小.高寒地区生物结皮对土壤养分影响过程及机理还需进一步研究. ...
Species composition, distribution patterns and ecological functions of biological soil crusts in the Gurbantunggut Desert
3
2010
... 生物结皮是由生长在表层土壤的微生物、藻类、苔藓和地衣等个体微小的生物成分与土壤相互作用形成的复合层[1].其广泛分布于干旱、半干旱区和极地、亚极地等脆弱生态区[2-3].近年来,关于生物结皮生态功能的研究已详细展开,研究表明生物结皮的形成可以改善土壤理化性质,提高土壤稳定性,改善土壤肥力等,对干旱半干旱地区植被恢复与重建具有重要的生态学意义[3-4].目前关于生物结皮生态功能的研究已有很多,但这些研究主要集中在干旱半干旱的沙漠地区[5-14],例如美国科罗拉多沙漠及其西南部的莫哈维沙漠[6-7],以色列南部的内盖夫沙漠[8],阿根廷的蒙特沙漠[9]以及干旱半干旱的西班牙地中海地区[10].国内关于生物结皮的研究也主要集中在沙漠及干旱半干旱地区,如黄土丘陵沟壑区[11-12]、腾格尔沙漠地区[13-14]、西北及新疆沙漠区[15-16].我国青藏高原素有“世界屋脊”、“第三极”之称,其海拔高,气温低,辐射强烈,日照多,黄河源区也源自于此[17].黄河源区是我国青藏高原重要的水源涵养区,也是该区域最主要的畜牧业基地之一.源区内独特的气候条件及其丰富的自然生态资源为该区经济发展及生态平衡发挥了生态安全屏障的作用[17-18].然而,20世纪70年代以来,随着气候的逐渐干旱化和人口、牲畜数量的迅速增加,该区生态环境急剧恶化,大面积植被退化,20世纪末,植被退化面积占源区草地总面积1/3[19].2005年以来,随着三江源地区实施草地“生态保护和建设”过程的具体实施(围栏封育、退牧还草、沙漠化治理、湿地保护等)[19-20],生物结皮开始在该地区逐渐发育.笔者在野外实地考察过程中发现,生物结皮在黄河源区不同退化程度的草地斑块中均有较大面积的分布.黄河源区平均气温-5.38~4.14 ℃,年降水量262.2~772.8 mm[20],在如此相对湿润且温度极低的条件下,生物结皮的发育特征与干热的沙漠地区是否一样,目前还不得而知.生物结皮作为地表特殊的结构层,其属于变水生物[21],在湿润时才能够进行新陈代谢活动[22].而青藏高原高黄河源区属于高原亚寒带半干旱区,仅有冷暖两季,冷季漫长,从每年10月初至次年4月之间地表处于冻结状态[23],降水主要以固态降雪为主,生物结皮呈现休眠状态.在半干旱低温环境条件下,生物结皮的理化性状如何,这一问题也尚不明晰. ...
... 生物结皮对土壤理化属性具有显著影响[1,3,14-15,28].关于生物结皮对土壤颗粒组成的影响,在不同的研究区域所得的研究结论存在一定分歧.有些研究结果表明,生物结皮的存在,能够细化土壤颗粒[28,34-35].也有研究表明,生物结皮对土壤颗粒组成并没有显著影响[36].本研究结果表明,由于生物结皮的发育可增加土壤中粉粒含量,5~20 cm土层中粉粒表现为深色藻结皮>浅色藻结皮>裸地,但差异并不显著.来自沙区的研究结果表明,生物结皮能够黏化土壤,捕捉土壤中细颗粒,使得表层土壤中黏粒和粉粒含量呈显著增加趋势.在黄土区的研究结果也表明,随着生物结皮的发育,结皮层土壤细颗相对增加[28].这些研究结果均表明生物结皮对土壤颗粒组成的影响集中在结皮层及其下0~5 cm左右,细颗粒呈现明显的表聚现象[28].而本研究结果发现由于结皮层的覆盖土壤粉粒在20~30 cm土层显著增加,这可能与该区域强烈的冻融过程及水热迁移有关.由于该区域早晚、四季之间温差较大,呈现明显的冻融循环作用[24-25],因此生物结皮层细颗粒可能由于冻融作用及水分迁移随之向下移动,聚集在其下20~30 cm土层.这种迁移作用可能随着水分含量以及发育年限的不同而发生相应的变化.与此同时,本研究结果表明,冻土区生物结皮层土壤容重也显著低于下层土壤,同时深色藻结皮不仅显著降低了自身容重,也影响到了其下0~20 cm土层土壤容重.而浅色藻结皮则对下层土壤容重影响不明显.本研究区海拔高于4 200 m,海拔高、风力大,与沙漠区域不同,生物结皮在形成过程中,并没有显著的捕捉细颗粒的作用,使结皮层土壤细化从而降低结皮层土壤容重.生物结皮在形成过程中,通过菌丝体、藻丝、假根或分泌的多糖来捆绑、黏结土壤颗粒,使得土壤形成许多团聚结构,土壤中孔隙增大,土壤容重降低[34],而深色藻结皮发育年限较长,厚度较厚,其孔隙度高,因此,容重较浅色藻结皮小,田间持水量显著高于浅色藻结皮及裸地. ...
... 生物结皮的形成,不仅可以改善土壤结构[28,37],也可显著影响表层土壤的养分累积[9,11,15,21].生物结皮中的固氮蓝藻的存在,可将大气中的氮固定从而输入到表层土壤中[38].同时生物结皮中的微生物能够进行固碳作用,可以增加表层土壤有机质含量[1,14,28],增加土壤肥力.虽然生物结皮对土壤养分积累效应已经形成一致结论,但其作用程度在不同区域还有所不同[14,28,40].本研究结果表明该区两种类型生物结皮均显著增加了结皮层及其下0~20 cm土层土壤有机质含量.藻类是生物结皮发育初期的先锋物种,它的形成能够向体外分泌以多糖为主的物质,从而形成生物结皮[41].同时,在给予充足的水分和光照条件下,生物结皮能够进行固碳作用,其固碳潜力主要取决于生物结皮的生物组成,研究已表明藓结皮的固碳能力高于藻结皮[42].该区域两种类型生物结皮,均以藻类结皮为主,因此其固碳能力可能没有显著差异.同时死亡的藻体又成为土壤有机物,为其他异氧微生物提供碳源[1],因此两种类型生物结皮均依靠显著增加了结皮层及其下0~20 cm土层土壤有机质含量.然而,两种生物结皮对土壤全氮含量的影响则有所不同.深色藻结皮的形成同时也提高了结皮层及其下0~20 cm土层土壤全氮含量,浅色藻结皮则仅提高了结皮层土层土壤全氮含量,对其下0~20 cm土层土壤全氮含量没有显著影响.生物结皮有机体在湿润状态下才能够进行生理代谢活动,进行相应的固氮作用,并且不同的物种固氮所需最佳水温条件不同[6].生物结皮在湿润后,其固氮活性强烈受制于水分[43].以上研究也表明,深色藻结皮的结皮层田间持水量显著高于浅色藻结皮,因此其固氮持续时间可能较长,故其固氮作用较强,向土壤中输入的氮素较多.氮素在水热交换作用下,向下层输入.而浅色藻结皮持水量较低,固氮作用较弱,因此其固定的氮素仅仅存留在结皮层.同时,生物结皮的存在降低了结皮层pH,但其对结皮层及下层土壤pH并没有显著影响.由于生物结皮某些藻类和地衣能够在湿润状态下进行生理活动,释放有机酸,使土壤pH降低[43],青藏高原黄河源区这一高寒地区气温常年低下,并且降水多以固态或暴雨的形式[44],并且在低温条件下,生物结皮有机体产生的有机酸较少也不易挥发释放出来,因此对土壤pH影响较小.高寒地区生物结皮对土壤养分影响过程及机理还需进一步研究. ...
The Distribution Patterns of Biological Soil Crust in Gurbantunggut Desert
3
2005
... 生物结皮是由生长在表层土壤的微生物、藻类、苔藓和地衣等个体微小的生物成分与土壤相互作用形成的复合层[1].其广泛分布于干旱、半干旱区和极地、亚极地等脆弱生态区[2-3].近年来,关于生物结皮生态功能的研究已详细展开,研究表明生物结皮的形成可以改善土壤理化性质,提高土壤稳定性,改善土壤肥力等,对干旱半干旱地区植被恢复与重建具有重要的生态学意义[3-4].目前关于生物结皮生态功能的研究已有很多,但这些研究主要集中在干旱半干旱的沙漠地区[5-14],例如美国科罗拉多沙漠及其西南部的莫哈维沙漠[6-7],以色列南部的内盖夫沙漠[8],阿根廷的蒙特沙漠[9]以及干旱半干旱的西班牙地中海地区[10].国内关于生物结皮的研究也主要集中在沙漠及干旱半干旱地区,如黄土丘陵沟壑区[11-12]、腾格尔沙漠地区[13-14]、西北及新疆沙漠区[15-16].我国青藏高原素有“世界屋脊”、“第三极”之称,其海拔高,气温低,辐射强烈,日照多,黄河源区也源自于此[17].黄河源区是我国青藏高原重要的水源涵养区,也是该区域最主要的畜牧业基地之一.源区内独特的气候条件及其丰富的自然生态资源为该区经济发展及生态平衡发挥了生态安全屏障的作用[17-18].然而,20世纪70年代以来,随着气候的逐渐干旱化和人口、牲畜数量的迅速增加,该区生态环境急剧恶化,大面积植被退化,20世纪末,植被退化面积占源区草地总面积1/3[19].2005年以来,随着三江源地区实施草地“生态保护和建设”过程的具体实施(围栏封育、退牧还草、沙漠化治理、湿地保护等)[19-20],生物结皮开始在该地区逐渐发育.笔者在野外实地考察过程中发现,生物结皮在黄河源区不同退化程度的草地斑块中均有较大面积的分布.黄河源区平均气温-5.38~4.14 ℃,年降水量262.2~772.8 mm[20],在如此相对湿润且温度极低的条件下,生物结皮的发育特征与干热的沙漠地区是否一样,目前还不得而知.生物结皮作为地表特殊的结构层,其属于变水生物[21],在湿润时才能够进行新陈代谢活动[22].而青藏高原高黄河源区属于高原亚寒带半干旱区,仅有冷暖两季,冷季漫长,从每年10月初至次年4月之间地表处于冻结状态[23],降水主要以固态降雪为主,生物结皮呈现休眠状态.在半干旱低温环境条件下,生物结皮的理化性状如何,这一问题也尚不明晰. ...
... 生物结皮可以在各种气候条件下广泛发育[1],国内外关于生物结皮的研究主要集中在荒漠地区及干旱半干旱的黄土高原等地区[4-16],关于青藏高原高寒冻土区生物结皮的发育特点鲜有报道.本研究结果表明冻土区生物结皮主要是以藻为主的生物结皮,结皮层厚度大约为6~22 mm.其在厚度上大于干旱的荒漠地区藻类结皮厚度(1~5 mm)[16].这与该地区特殊的生态环境条件密切相关,由于冻土区常年温度低,生物结皮活性较弱,呼吸等生理活性缓慢[4,21],其中积累的物质不易分解,因此,生物结皮层厚度较大.同时,生物结皮的厚度与生物结皮的发育年限密切相关[29].因此,该区域植被退化区域生物结皮(浅色藻结皮)较未退化区生物结皮(深色藻结皮)发育年限短,故其深色藻结皮厚度大于浅色藻结生物结皮小. ...
... [16].这与该地区特殊的生态环境条件密切相关,由于冻土区常年温度低,生物结皮活性较弱,呼吸等生理活性缓慢[4,21],其中积累的物质不易分解,因此,生物结皮层厚度较大.同时,生物结皮的厚度与生物结皮的发育年限密切相关[29].因此,该区域植被退化区域生物结皮(浅色藻结皮)较未退化区生物结皮(深色藻结皮)发育年限短,故其深色藻结皮厚度大于浅色藻结生物结皮小. ...
古尔班通古特沙漠生物结皮的分布特征
3
2005
... 生物结皮是由生长在表层土壤的微生物、藻类、苔藓和地衣等个体微小的生物成分与土壤相互作用形成的复合层[1].其广泛分布于干旱、半干旱区和极地、亚极地等脆弱生态区[2-3].近年来,关于生物结皮生态功能的研究已详细展开,研究表明生物结皮的形成可以改善土壤理化性质,提高土壤稳定性,改善土壤肥力等,对干旱半干旱地区植被恢复与重建具有重要的生态学意义[3-4].目前关于生物结皮生态功能的研究已有很多,但这些研究主要集中在干旱半干旱的沙漠地区[5-14],例如美国科罗拉多沙漠及其西南部的莫哈维沙漠[6-7],以色列南部的内盖夫沙漠[8],阿根廷的蒙特沙漠[9]以及干旱半干旱的西班牙地中海地区[10].国内关于生物结皮的研究也主要集中在沙漠及干旱半干旱地区,如黄土丘陵沟壑区[11-12]、腾格尔沙漠地区[13-14]、西北及新疆沙漠区[15-16].我国青藏高原素有“世界屋脊”、“第三极”之称,其海拔高,气温低,辐射强烈,日照多,黄河源区也源自于此[17].黄河源区是我国青藏高原重要的水源涵养区,也是该区域最主要的畜牧业基地之一.源区内独特的气候条件及其丰富的自然生态资源为该区经济发展及生态平衡发挥了生态安全屏障的作用[17-18].然而,20世纪70年代以来,随着气候的逐渐干旱化和人口、牲畜数量的迅速增加,该区生态环境急剧恶化,大面积植被退化,20世纪末,植被退化面积占源区草地总面积1/3[19].2005年以来,随着三江源地区实施草地“生态保护和建设”过程的具体实施(围栏封育、退牧还草、沙漠化治理、湿地保护等)[19-20],生物结皮开始在该地区逐渐发育.笔者在野外实地考察过程中发现,生物结皮在黄河源区不同退化程度的草地斑块中均有较大面积的分布.黄河源区平均气温-5.38~4.14 ℃,年降水量262.2~772.8 mm[20],在如此相对湿润且温度极低的条件下,生物结皮的发育特征与干热的沙漠地区是否一样,目前还不得而知.生物结皮作为地表特殊的结构层,其属于变水生物[21],在湿润时才能够进行新陈代谢活动[22].而青藏高原高黄河源区属于高原亚寒带半干旱区,仅有冷暖两季,冷季漫长,从每年10月初至次年4月之间地表处于冻结状态[23],降水主要以固态降雪为主,生物结皮呈现休眠状态.在半干旱低温环境条件下,生物结皮的理化性状如何,这一问题也尚不明晰. ...
... 生物结皮可以在各种气候条件下广泛发育[1],国内外关于生物结皮的研究主要集中在荒漠地区及干旱半干旱的黄土高原等地区[4-16],关于青藏高原高寒冻土区生物结皮的发育特点鲜有报道.本研究结果表明冻土区生物结皮主要是以藻为主的生物结皮,结皮层厚度大约为6~22 mm.其在厚度上大于干旱的荒漠地区藻类结皮厚度(1~5 mm)[16].这与该地区特殊的生态环境条件密切相关,由于冻土区常年温度低,生物结皮活性较弱,呼吸等生理活性缓慢[4,21],其中积累的物质不易分解,因此,生物结皮层厚度较大.同时,生物结皮的厚度与生物结皮的发育年限密切相关[29].因此,该区域植被退化区域生物结皮(浅色藻结皮)较未退化区生物结皮(深色藻结皮)发育年限短,故其深色藻结皮厚度大于浅色藻结生物结皮小. ...
... [16].这与该地区特殊的生态环境条件密切相关,由于冻土区常年温度低,生物结皮活性较弱,呼吸等生理活性缓慢[4,21],其中积累的物质不易分解,因此,生物结皮层厚度较大.同时,生物结皮的厚度与生物结皮的发育年限密切相关[29].因此,该区域植被退化区域生物结皮(浅色藻结皮)较未退化区生物结皮(深色藻结皮)发育年限短,故其深色藻结皮厚度大于浅色藻结生物结皮小. ...
Ecosystem water conservation changes in response to climate change in the source region of the Yellow River from 1981 to 2010
2
2016
... 生物结皮是由生长在表层土壤的微生物、藻类、苔藓和地衣等个体微小的生物成分与土壤相互作用形成的复合层[1].其广泛分布于干旱、半干旱区和极地、亚极地等脆弱生态区[2-3].近年来,关于生物结皮生态功能的研究已详细展开,研究表明生物结皮的形成可以改善土壤理化性质,提高土壤稳定性,改善土壤肥力等,对干旱半干旱地区植被恢复与重建具有重要的生态学意义[3-4].目前关于生物结皮生态功能的研究已有很多,但这些研究主要集中在干旱半干旱的沙漠地区[5-14],例如美国科罗拉多沙漠及其西南部的莫哈维沙漠[6-7],以色列南部的内盖夫沙漠[8],阿根廷的蒙特沙漠[9]以及干旱半干旱的西班牙地中海地区[10].国内关于生物结皮的研究也主要集中在沙漠及干旱半干旱地区,如黄土丘陵沟壑区[11-12]、腾格尔沙漠地区[13-14]、西北及新疆沙漠区[15-16].我国青藏高原素有“世界屋脊”、“第三极”之称,其海拔高,气温低,辐射强烈,日照多,黄河源区也源自于此[17].黄河源区是我国青藏高原重要的水源涵养区,也是该区域最主要的畜牧业基地之一.源区内独特的气候条件及其丰富的自然生态资源为该区经济发展及生态平衡发挥了生态安全屏障的作用[17-18].然而,20世纪70年代以来,随着气候的逐渐干旱化和人口、牲畜数量的迅速增加,该区生态环境急剧恶化,大面积植被退化,20世纪末,植被退化面积占源区草地总面积1/3[19].2005年以来,随着三江源地区实施草地“生态保护和建设”过程的具体实施(围栏封育、退牧还草、沙漠化治理、湿地保护等)[19-20],生物结皮开始在该地区逐渐发育.笔者在野外实地考察过程中发现,生物结皮在黄河源区不同退化程度的草地斑块中均有较大面积的分布.黄河源区平均气温-5.38~4.14 ℃,年降水量262.2~772.8 mm[20],在如此相对湿润且温度极低的条件下,生物结皮的发育特征与干热的沙漠地区是否一样,目前还不得而知.生物结皮作为地表特殊的结构层,其属于变水生物[21],在湿润时才能够进行新陈代谢活动[22].而青藏高原高黄河源区属于高原亚寒带半干旱区,仅有冷暖两季,冷季漫长,从每年10月初至次年4月之间地表处于冻结状态[23],降水主要以固态降雪为主,生物结皮呈现休眠状态.在半干旱低温环境条件下,生物结皮的理化性状如何,这一问题也尚不明晰. ...
... [17-18].然而,20世纪70年代以来,随着气候的逐渐干旱化和人口、牲畜数量的迅速增加,该区生态环境急剧恶化,大面积植被退化,20世纪末,植被退化面积占源区草地总面积1/3[19].2005年以来,随着三江源地区实施草地“生态保护和建设”过程的具体实施(围栏封育、退牧还草、沙漠化治理、湿地保护等)[19-20],生物结皮开始在该地区逐渐发育.笔者在野外实地考察过程中发现,生物结皮在黄河源区不同退化程度的草地斑块中均有较大面积的分布.黄河源区平均气温-5.38~4.14 ℃,年降水量262.2~772.8 mm[20],在如此相对湿润且温度极低的条件下,生物结皮的发育特征与干热的沙漠地区是否一样,目前还不得而知.生物结皮作为地表特殊的结构层,其属于变水生物[21],在湿润时才能够进行新陈代谢活动[22].而青藏高原高黄河源区属于高原亚寒带半干旱区,仅有冷暖两季,冷季漫长,从每年10月初至次年4月之间地表处于冻结状态[23],降水主要以固态降雪为主,生物结皮呈现休眠状态.在半干旱低温环境条件下,生物结皮的理化性状如何,这一问题也尚不明晰. ...
过去30年气候变化对黄河源区水源涵养量的影响
2
2016
... 生物结皮是由生长在表层土壤的微生物、藻类、苔藓和地衣等个体微小的生物成分与土壤相互作用形成的复合层[1].其广泛分布于干旱、半干旱区和极地、亚极地等脆弱生态区[2-3].近年来,关于生物结皮生态功能的研究已详细展开,研究表明生物结皮的形成可以改善土壤理化性质,提高土壤稳定性,改善土壤肥力等,对干旱半干旱地区植被恢复与重建具有重要的生态学意义[3-4].目前关于生物结皮生态功能的研究已有很多,但这些研究主要集中在干旱半干旱的沙漠地区[5-14],例如美国科罗拉多沙漠及其西南部的莫哈维沙漠[6-7],以色列南部的内盖夫沙漠[8],阿根廷的蒙特沙漠[9]以及干旱半干旱的西班牙地中海地区[10].国内关于生物结皮的研究也主要集中在沙漠及干旱半干旱地区,如黄土丘陵沟壑区[11-12]、腾格尔沙漠地区[13-14]、西北及新疆沙漠区[15-16].我国青藏高原素有“世界屋脊”、“第三极”之称,其海拔高,气温低,辐射强烈,日照多,黄河源区也源自于此[17].黄河源区是我国青藏高原重要的水源涵养区,也是该区域最主要的畜牧业基地之一.源区内独特的气候条件及其丰富的自然生态资源为该区经济发展及生态平衡发挥了生态安全屏障的作用[17-18].然而,20世纪70年代以来,随着气候的逐渐干旱化和人口、牲畜数量的迅速增加,该区生态环境急剧恶化,大面积植被退化,20世纪末,植被退化面积占源区草地总面积1/3[19].2005年以来,随着三江源地区实施草地“生态保护和建设”过程的具体实施(围栏封育、退牧还草、沙漠化治理、湿地保护等)[19-20],生物结皮开始在该地区逐渐发育.笔者在野外实地考察过程中发现,生物结皮在黄河源区不同退化程度的草地斑块中均有较大面积的分布.黄河源区平均气温-5.38~4.14 ℃,年降水量262.2~772.8 mm[20],在如此相对湿润且温度极低的条件下,生物结皮的发育特征与干热的沙漠地区是否一样,目前还不得而知.生物结皮作为地表特殊的结构层,其属于变水生物[21],在湿润时才能够进行新陈代谢活动[22].而青藏高原高黄河源区属于高原亚寒带半干旱区,仅有冷暖两季,冷季漫长,从每年10月初至次年4月之间地表处于冻结状态[23],降水主要以固态降雪为主,生物结皮呈现休眠状态.在半干旱低温环境条件下,生物结皮的理化性状如何,这一问题也尚不明晰. ...
... [17-18].然而,20世纪70年代以来,随着气候的逐渐干旱化和人口、牲畜数量的迅速增加,该区生态环境急剧恶化,大面积植被退化,20世纪末,植被退化面积占源区草地总面积1/3[19].2005年以来,随着三江源地区实施草地“生态保护和建设”过程的具体实施(围栏封育、退牧还草、沙漠化治理、湿地保护等)[19-20],生物结皮开始在该地区逐渐发育.笔者在野外实地考察过程中发现,生物结皮在黄河源区不同退化程度的草地斑块中均有较大面积的分布.黄河源区平均气温-5.38~4.14 ℃,年降水量262.2~772.8 mm[20],在如此相对湿润且温度极低的条件下,生物结皮的发育特征与干热的沙漠地区是否一样,目前还不得而知.生物结皮作为地表特殊的结构层,其属于变水生物[21],在湿润时才能够进行新陈代谢活动[22].而青藏高原高黄河源区属于高原亚寒带半干旱区,仅有冷暖两季,冷季漫长,从每年10月初至次年4月之间地表处于冻结状态[23],降水主要以固态降雪为主,生物结皮呈现休眠状态.在半干旱低温环境条件下,生物结皮的理化性状如何,这一问题也尚不明晰. ...
Spatiotemporal dynamics of grassland coverage in response to climate change from 1982 to 2012 in the Three Rivers Source Region
1
2017
... 生物结皮是由生长在表层土壤的微生物、藻类、苔藓和地衣等个体微小的生物成分与土壤相互作用形成的复合层[1].其广泛分布于干旱、半干旱区和极地、亚极地等脆弱生态区[2-3].近年来,关于生物结皮生态功能的研究已详细展开,研究表明生物结皮的形成可以改善土壤理化性质,提高土壤稳定性,改善土壤肥力等,对干旱半干旱地区植被恢复与重建具有重要的生态学意义[3-4].目前关于生物结皮生态功能的研究已有很多,但这些研究主要集中在干旱半干旱的沙漠地区[5-14],例如美国科罗拉多沙漠及其西南部的莫哈维沙漠[6-7],以色列南部的内盖夫沙漠[8],阿根廷的蒙特沙漠[9]以及干旱半干旱的西班牙地中海地区[10].国内关于生物结皮的研究也主要集中在沙漠及干旱半干旱地区,如黄土丘陵沟壑区[11-12]、腾格尔沙漠地区[13-14]、西北及新疆沙漠区[15-16].我国青藏高原素有“世界屋脊”、“第三极”之称,其海拔高,气温低,辐射强烈,日照多,黄河源区也源自于此[17].黄河源区是我国青藏高原重要的水源涵养区,也是该区域最主要的畜牧业基地之一.源区内独特的气候条件及其丰富的自然生态资源为该区经济发展及生态平衡发挥了生态安全屏障的作用[17-18].然而,20世纪70年代以来,随着气候的逐渐干旱化和人口、牲畜数量的迅速增加,该区生态环境急剧恶化,大面积植被退化,20世纪末,植被退化面积占源区草地总面积1/3[19].2005年以来,随着三江源地区实施草地“生态保护和建设”过程的具体实施(围栏封育、退牧还草、沙漠化治理、湿地保护等)[19-20],生物结皮开始在该地区逐渐发育.笔者在野外实地考察过程中发现,生物结皮在黄河源区不同退化程度的草地斑块中均有较大面积的分布.黄河源区平均气温-5.38~4.14 ℃,年降水量262.2~772.8 mm[20],在如此相对湿润且温度极低的条件下,生物结皮的发育特征与干热的沙漠地区是否一样,目前还不得而知.生物结皮作为地表特殊的结构层,其属于变水生物[21],在湿润时才能够进行新陈代谢活动[22].而青藏高原高黄河源区属于高原亚寒带半干旱区,仅有冷暖两季,冷季漫长,从每年10月初至次年4月之间地表处于冻结状态[23],降水主要以固态降雪为主,生物结皮呈现休眠状态.在半干旱低温环境条件下,生物结皮的理化性状如何,这一问题也尚不明晰. ...
三江源1982-2012年草地植被覆盖度动态及其对气候变化的响应
1
2017
... 生物结皮是由生长在表层土壤的微生物、藻类、苔藓和地衣等个体微小的生物成分与土壤相互作用形成的复合层[1].其广泛分布于干旱、半干旱区和极地、亚极地等脆弱生态区[2-3].近年来,关于生物结皮生态功能的研究已详细展开,研究表明生物结皮的形成可以改善土壤理化性质,提高土壤稳定性,改善土壤肥力等,对干旱半干旱地区植被恢复与重建具有重要的生态学意义[3-4].目前关于生物结皮生态功能的研究已有很多,但这些研究主要集中在干旱半干旱的沙漠地区[5-14],例如美国科罗拉多沙漠及其西南部的莫哈维沙漠[6-7],以色列南部的内盖夫沙漠[8],阿根廷的蒙特沙漠[9]以及干旱半干旱的西班牙地中海地区[10].国内关于生物结皮的研究也主要集中在沙漠及干旱半干旱地区,如黄土丘陵沟壑区[11-12]、腾格尔沙漠地区[13-14]、西北及新疆沙漠区[15-16].我国青藏高原素有“世界屋脊”、“第三极”之称,其海拔高,气温低,辐射强烈,日照多,黄河源区也源自于此[17].黄河源区是我国青藏高原重要的水源涵养区,也是该区域最主要的畜牧业基地之一.源区内独特的气候条件及其丰富的自然生态资源为该区经济发展及生态平衡发挥了生态安全屏障的作用[17-18].然而,20世纪70年代以来,随着气候的逐渐干旱化和人口、牲畜数量的迅速增加,该区生态环境急剧恶化,大面积植被退化,20世纪末,植被退化面积占源区草地总面积1/3[19].2005年以来,随着三江源地区实施草地“生态保护和建设”过程的具体实施(围栏封育、退牧还草、沙漠化治理、湿地保护等)[19-20],生物结皮开始在该地区逐渐发育.笔者在野外实地考察过程中发现,生物结皮在黄河源区不同退化程度的草地斑块中均有较大面积的分布.黄河源区平均气温-5.38~4.14 ℃,年降水量262.2~772.8 mm[20],在如此相对湿润且温度极低的条件下,生物结皮的发育特征与干热的沙漠地区是否一样,目前还不得而知.生物结皮作为地表特殊的结构层,其属于变水生物[21],在湿润时才能够进行新陈代谢活动[22].而青藏高原高黄河源区属于高原亚寒带半干旱区,仅有冷暖两季,冷季漫长,从每年10月初至次年4月之间地表处于冻结状态[23],降水主要以固态降雪为主,生物结皮呈现休眠状态.在半干旱低温环境条件下,生物结皮的理化性状如何,这一问题也尚不明晰. ...
Development and cause of aeolian desertification in alpine region: in case of Maduo County in Yellow River Source Area
2
2007
... 生物结皮是由生长在表层土壤的微生物、藻类、苔藓和地衣等个体微小的生物成分与土壤相互作用形成的复合层[1].其广泛分布于干旱、半干旱区和极地、亚极地等脆弱生态区[2-3].近年来,关于生物结皮生态功能的研究已详细展开,研究表明生物结皮的形成可以改善土壤理化性质,提高土壤稳定性,改善土壤肥力等,对干旱半干旱地区植被恢复与重建具有重要的生态学意义[3-4].目前关于生物结皮生态功能的研究已有很多,但这些研究主要集中在干旱半干旱的沙漠地区[5-14],例如美国科罗拉多沙漠及其西南部的莫哈维沙漠[6-7],以色列南部的内盖夫沙漠[8],阿根廷的蒙特沙漠[9]以及干旱半干旱的西班牙地中海地区[10].国内关于生物结皮的研究也主要集中在沙漠及干旱半干旱地区,如黄土丘陵沟壑区[11-12]、腾格尔沙漠地区[13-14]、西北及新疆沙漠区[15-16].我国青藏高原素有“世界屋脊”、“第三极”之称,其海拔高,气温低,辐射强烈,日照多,黄河源区也源自于此[17].黄河源区是我国青藏高原重要的水源涵养区,也是该区域最主要的畜牧业基地之一.源区内独特的气候条件及其丰富的自然生态资源为该区经济发展及生态平衡发挥了生态安全屏障的作用[17-18].然而,20世纪70年代以来,随着气候的逐渐干旱化和人口、牲畜数量的迅速增加,该区生态环境急剧恶化,大面积植被退化,20世纪末,植被退化面积占源区草地总面积1/3[19].2005年以来,随着三江源地区实施草地“生态保护和建设”过程的具体实施(围栏封育、退牧还草、沙漠化治理、湿地保护等)[19-20],生物结皮开始在该地区逐渐发育.笔者在野外实地考察过程中发现,生物结皮在黄河源区不同退化程度的草地斑块中均有较大面积的分布.黄河源区平均气温-5.38~4.14 ℃,年降水量262.2~772.8 mm[20],在如此相对湿润且温度极低的条件下,生物结皮的发育特征与干热的沙漠地区是否一样,目前还不得而知.生物结皮作为地表特殊的结构层,其属于变水生物[21],在湿润时才能够进行新陈代谢活动[22].而青藏高原高黄河源区属于高原亚寒带半干旱区,仅有冷暖两季,冷季漫长,从每年10月初至次年4月之间地表处于冻结状态[23],降水主要以固态降雪为主,生物结皮呈现休眠状态.在半干旱低温环境条件下,生物结皮的理化性状如何,这一问题也尚不明晰. ...
... [19-20],生物结皮开始在该地区逐渐发育.笔者在野外实地考察过程中发现,生物结皮在黄河源区不同退化程度的草地斑块中均有较大面积的分布.黄河源区平均气温-5.38~4.14 ℃,年降水量262.2~772.8 mm[20],在如此相对湿润且温度极低的条件下,生物结皮的发育特征与干热的沙漠地区是否一样,目前还不得而知.生物结皮作为地表特殊的结构层,其属于变水生物[21],在湿润时才能够进行新陈代谢活动[22].而青藏高原高黄河源区属于高原亚寒带半干旱区,仅有冷暖两季,冷季漫长,从每年10月初至次年4月之间地表处于冻结状态[23],降水主要以固态降雪为主,生物结皮呈现休眠状态.在半干旱低温环境条件下,生物结皮的理化性状如何,这一问题也尚不明晰. ...
高寒草甸地区沙漠化发展过程及成因——以黄河源区玛多县为例
2
2007
... 生物结皮是由生长在表层土壤的微生物、藻类、苔藓和地衣等个体微小的生物成分与土壤相互作用形成的复合层[1].其广泛分布于干旱、半干旱区和极地、亚极地等脆弱生态区[2-3].近年来,关于生物结皮生态功能的研究已详细展开,研究表明生物结皮的形成可以改善土壤理化性质,提高土壤稳定性,改善土壤肥力等,对干旱半干旱地区植被恢复与重建具有重要的生态学意义[3-4].目前关于生物结皮生态功能的研究已有很多,但这些研究主要集中在干旱半干旱的沙漠地区[5-14],例如美国科罗拉多沙漠及其西南部的莫哈维沙漠[6-7],以色列南部的内盖夫沙漠[8],阿根廷的蒙特沙漠[9]以及干旱半干旱的西班牙地中海地区[10].国内关于生物结皮的研究也主要集中在沙漠及干旱半干旱地区,如黄土丘陵沟壑区[11-12]、腾格尔沙漠地区[13-14]、西北及新疆沙漠区[15-16].我国青藏高原素有“世界屋脊”、“第三极”之称,其海拔高,气温低,辐射强烈,日照多,黄河源区也源自于此[17].黄河源区是我国青藏高原重要的水源涵养区,也是该区域最主要的畜牧业基地之一.源区内独特的气候条件及其丰富的自然生态资源为该区经济发展及生态平衡发挥了生态安全屏障的作用[17-18].然而,20世纪70年代以来,随着气候的逐渐干旱化和人口、牲畜数量的迅速增加,该区生态环境急剧恶化,大面积植被退化,20世纪末,植被退化面积占源区草地总面积1/3[19].2005年以来,随着三江源地区实施草地“生态保护和建设”过程的具体实施(围栏封育、退牧还草、沙漠化治理、湿地保护等)[19-20],生物结皮开始在该地区逐渐发育.笔者在野外实地考察过程中发现,生物结皮在黄河源区不同退化程度的草地斑块中均有较大面积的分布.黄河源区平均气温-5.38~4.14 ℃,年降水量262.2~772.8 mm[20],在如此相对湿润且温度极低的条件下,生物结皮的发育特征与干热的沙漠地区是否一样,目前还不得而知.生物结皮作为地表特殊的结构层,其属于变水生物[21],在湿润时才能够进行新陈代谢活动[22].而青藏高原高黄河源区属于高原亚寒带半干旱区,仅有冷暖两季,冷季漫长,从每年10月初至次年4月之间地表处于冻结状态[23],降水主要以固态降雪为主,生物结皮呈现休眠状态.在半干旱低温环境条件下,生物结皮的理化性状如何,这一问题也尚不明晰. ...
... [19-20],生物结皮开始在该地区逐渐发育.笔者在野外实地考察过程中发现,生物结皮在黄河源区不同退化程度的草地斑块中均有较大面积的分布.黄河源区平均气温-5.38~4.14 ℃,年降水量262.2~772.8 mm[20],在如此相对湿润且温度极低的条件下,生物结皮的发育特征与干热的沙漠地区是否一样,目前还不得而知.生物结皮作为地表特殊的结构层,其属于变水生物[21],在湿润时才能够进行新陈代谢活动[22].而青藏高原高黄河源区属于高原亚寒带半干旱区,仅有冷暖两季,冷季漫长,从每年10月初至次年4月之间地表处于冻结状态[23],降水主要以固态降雪为主,生物结皮呈现休眠状态.在半干旱低温环境条件下,生物结皮的理化性状如何,这一问题也尚不明晰. ...
Simulation and evaluation of forage height in alpine meadow grassland in headwaters of Yellow River
2
2018
... 生物结皮是由生长在表层土壤的微生物、藻类、苔藓和地衣等个体微小的生物成分与土壤相互作用形成的复合层[1].其广泛分布于干旱、半干旱区和极地、亚极地等脆弱生态区[2-3].近年来,关于生物结皮生态功能的研究已详细展开,研究表明生物结皮的形成可以改善土壤理化性质,提高土壤稳定性,改善土壤肥力等,对干旱半干旱地区植被恢复与重建具有重要的生态学意义[3-4].目前关于生物结皮生态功能的研究已有很多,但这些研究主要集中在干旱半干旱的沙漠地区[5-14],例如美国科罗拉多沙漠及其西南部的莫哈维沙漠[6-7],以色列南部的内盖夫沙漠[8],阿根廷的蒙特沙漠[9]以及干旱半干旱的西班牙地中海地区[10].国内关于生物结皮的研究也主要集中在沙漠及干旱半干旱地区,如黄土丘陵沟壑区[11-12]、腾格尔沙漠地区[13-14]、西北及新疆沙漠区[15-16].我国青藏高原素有“世界屋脊”、“第三极”之称,其海拔高,气温低,辐射强烈,日照多,黄河源区也源自于此[17].黄河源区是我国青藏高原重要的水源涵养区,也是该区域最主要的畜牧业基地之一.源区内独特的气候条件及其丰富的自然生态资源为该区经济发展及生态平衡发挥了生态安全屏障的作用[17-18].然而,20世纪70年代以来,随着气候的逐渐干旱化和人口、牲畜数量的迅速增加,该区生态环境急剧恶化,大面积植被退化,20世纪末,植被退化面积占源区草地总面积1/3[19].2005年以来,随着三江源地区实施草地“生态保护和建设”过程的具体实施(围栏封育、退牧还草、沙漠化治理、湿地保护等)[19-20],生物结皮开始在该地区逐渐发育.笔者在野外实地考察过程中发现,生物结皮在黄河源区不同退化程度的草地斑块中均有较大面积的分布.黄河源区平均气温-5.38~4.14 ℃,年降水量262.2~772.8 mm[20],在如此相对湿润且温度极低的条件下,生物结皮的发育特征与干热的沙漠地区是否一样,目前还不得而知.生物结皮作为地表特殊的结构层,其属于变水生物[21],在湿润时才能够进行新陈代谢活动[22].而青藏高原高黄河源区属于高原亚寒带半干旱区,仅有冷暖两季,冷季漫长,从每年10月初至次年4月之间地表处于冻结状态[23],降水主要以固态降雪为主,生物结皮呈现休眠状态.在半干旱低温环境条件下,生物结皮的理化性状如何,这一问题也尚不明晰. ...
... [20],在如此相对湿润且温度极低的条件下,生物结皮的发育特征与干热的沙漠地区是否一样,目前还不得而知.生物结皮作为地表特殊的结构层,其属于变水生物[21],在湿润时才能够进行新陈代谢活动[22].而青藏高原高黄河源区属于高原亚寒带半干旱区,仅有冷暖两季,冷季漫长,从每年10月初至次年4月之间地表处于冻结状态[23],降水主要以固态降雪为主,生物结皮呈现休眠状态.在半干旱低温环境条件下,生物结皮的理化性状如何,这一问题也尚不明晰. ...
黄河源区高寒草甸群落高度遥感模拟与评价
2
2018
... 生物结皮是由生长在表层土壤的微生物、藻类、苔藓和地衣等个体微小的生物成分与土壤相互作用形成的复合层[1].其广泛分布于干旱、半干旱区和极地、亚极地等脆弱生态区[2-3].近年来,关于生物结皮生态功能的研究已详细展开,研究表明生物结皮的形成可以改善土壤理化性质,提高土壤稳定性,改善土壤肥力等,对干旱半干旱地区植被恢复与重建具有重要的生态学意义[3-4].目前关于生物结皮生态功能的研究已有很多,但这些研究主要集中在干旱半干旱的沙漠地区[5-14],例如美国科罗拉多沙漠及其西南部的莫哈维沙漠[6-7],以色列南部的内盖夫沙漠[8],阿根廷的蒙特沙漠[9]以及干旱半干旱的西班牙地中海地区[10].国内关于生物结皮的研究也主要集中在沙漠及干旱半干旱地区,如黄土丘陵沟壑区[11-12]、腾格尔沙漠地区[13-14]、西北及新疆沙漠区[15-16].我国青藏高原素有“世界屋脊”、“第三极”之称,其海拔高,气温低,辐射强烈,日照多,黄河源区也源自于此[17].黄河源区是我国青藏高原重要的水源涵养区,也是该区域最主要的畜牧业基地之一.源区内独特的气候条件及其丰富的自然生态资源为该区经济发展及生态平衡发挥了生态安全屏障的作用[17-18].然而,20世纪70年代以来,随着气候的逐渐干旱化和人口、牲畜数量的迅速增加,该区生态环境急剧恶化,大面积植被退化,20世纪末,植被退化面积占源区草地总面积1/3[19].2005年以来,随着三江源地区实施草地“生态保护和建设”过程的具体实施(围栏封育、退牧还草、沙漠化治理、湿地保护等)[19-20],生物结皮开始在该地区逐渐发育.笔者在野外实地考察过程中发现,生物结皮在黄河源区不同退化程度的草地斑块中均有较大面积的分布.黄河源区平均气温-5.38~4.14 ℃,年降水量262.2~772.8 mm[20],在如此相对湿润且温度极低的条件下,生物结皮的发育特征与干热的沙漠地区是否一样,目前还不得而知.生物结皮作为地表特殊的结构层,其属于变水生物[21],在湿润时才能够进行新陈代谢活动[22].而青藏高原高黄河源区属于高原亚寒带半干旱区,仅有冷暖两季,冷季漫长,从每年10月初至次年4月之间地表处于冻结状态[23],降水主要以固态降雪为主,生物结皮呈现休眠状态.在半干旱低温环境条件下,生物结皮的理化性状如何,这一问题也尚不明晰. ...
... [20],在如此相对湿润且温度极低的条件下,生物结皮的发育特征与干热的沙漠地区是否一样,目前还不得而知.生物结皮作为地表特殊的结构层,其属于变水生物[21],在湿润时才能够进行新陈代谢活动[22].而青藏高原高黄河源区属于高原亚寒带半干旱区,仅有冷暖两季,冷季漫长,从每年10月初至次年4月之间地表处于冻结状态[23],降水主要以固态降雪为主,生物结皮呈现休眠状态.在半干旱低温环境条件下,生物结皮的理化性状如何,这一问题也尚不明晰. ...
3
1996
... 生物结皮是由生长在表层土壤的微生物、藻类、苔藓和地衣等个体微小的生物成分与土壤相互作用形成的复合层[1].其广泛分布于干旱、半干旱区和极地、亚极地等脆弱生态区[2-3].近年来,关于生物结皮生态功能的研究已详细展开,研究表明生物结皮的形成可以改善土壤理化性质,提高土壤稳定性,改善土壤肥力等,对干旱半干旱地区植被恢复与重建具有重要的生态学意义[3-4].目前关于生物结皮生态功能的研究已有很多,但这些研究主要集中在干旱半干旱的沙漠地区[5-14],例如美国科罗拉多沙漠及其西南部的莫哈维沙漠[6-7],以色列南部的内盖夫沙漠[8],阿根廷的蒙特沙漠[9]以及干旱半干旱的西班牙地中海地区[10].国内关于生物结皮的研究也主要集中在沙漠及干旱半干旱地区,如黄土丘陵沟壑区[11-12]、腾格尔沙漠地区[13-14]、西北及新疆沙漠区[15-16].我国青藏高原素有“世界屋脊”、“第三极”之称,其海拔高,气温低,辐射强烈,日照多,黄河源区也源自于此[17].黄河源区是我国青藏高原重要的水源涵养区,也是该区域最主要的畜牧业基地之一.源区内独特的气候条件及其丰富的自然生态资源为该区经济发展及生态平衡发挥了生态安全屏障的作用[17-18].然而,20世纪70年代以来,随着气候的逐渐干旱化和人口、牲畜数量的迅速增加,该区生态环境急剧恶化,大面积植被退化,20世纪末,植被退化面积占源区草地总面积1/3[19].2005年以来,随着三江源地区实施草地“生态保护和建设”过程的具体实施(围栏封育、退牧还草、沙漠化治理、湿地保护等)[19-20],生物结皮开始在该地区逐渐发育.笔者在野外实地考察过程中发现,生物结皮在黄河源区不同退化程度的草地斑块中均有较大面积的分布.黄河源区平均气温-5.38~4.14 ℃,年降水量262.2~772.8 mm[20],在如此相对湿润且温度极低的条件下,生物结皮的发育特征与干热的沙漠地区是否一样,目前还不得而知.生物结皮作为地表特殊的结构层,其属于变水生物[21],在湿润时才能够进行新陈代谢活动[22].而青藏高原高黄河源区属于高原亚寒带半干旱区,仅有冷暖两季,冷季漫长,从每年10月初至次年4月之间地表处于冻结状态[23],降水主要以固态降雪为主,生物结皮呈现休眠状态.在半干旱低温环境条件下,生物结皮的理化性状如何,这一问题也尚不明晰. ...
... 生物结皮可以在各种气候条件下广泛发育[1],国内外关于生物结皮的研究主要集中在荒漠地区及干旱半干旱的黄土高原等地区[4-16],关于青藏高原高寒冻土区生物结皮的发育特点鲜有报道.本研究结果表明冻土区生物结皮主要是以藻为主的生物结皮,结皮层厚度大约为6~22 mm.其在厚度上大于干旱的荒漠地区藻类结皮厚度(1~5 mm)[16].这与该地区特殊的生态环境条件密切相关,由于冻土区常年温度低,生物结皮活性较弱,呼吸等生理活性缓慢[4,21],其中积累的物质不易分解,因此,生物结皮层厚度较大.同时,生物结皮的厚度与生物结皮的发育年限密切相关[29].因此,该区域植被退化区域生物结皮(浅色藻结皮)较未退化区生物结皮(深色藻结皮)发育年限短,故其深色藻结皮厚度大于浅色藻结生物结皮小. ...
... 生物结皮的形成,不仅可以改善土壤结构[28,37],也可显著影响表层土壤的养分累积[9,11,15,21].生物结皮中的固氮蓝藻的存在,可将大气中的氮固定从而输入到表层土壤中[38].同时生物结皮中的微生物能够进行固碳作用,可以增加表层土壤有机质含量[1,14,28],增加土壤肥力.虽然生物结皮对土壤养分积累效应已经形成一致结论,但其作用程度在不同区域还有所不同[14,28,40].本研究结果表明该区两种类型生物结皮均显著增加了结皮层及其下0~20 cm土层土壤有机质含量.藻类是生物结皮发育初期的先锋物种,它的形成能够向体外分泌以多糖为主的物质,从而形成生物结皮[41].同时,在给予充足的水分和光照条件下,生物结皮能够进行固碳作用,其固碳潜力主要取决于生物结皮的生物组成,研究已表明藓结皮的固碳能力高于藻结皮[42].该区域两种类型生物结皮,均以藻类结皮为主,因此其固碳能力可能没有显著差异.同时死亡的藻体又成为土壤有机物,为其他异氧微生物提供碳源[1],因此两种类型生物结皮均依靠显著增加了结皮层及其下0~20 cm土层土壤有机质含量.然而,两种生物结皮对土壤全氮含量的影响则有所不同.深色藻结皮的形成同时也提高了结皮层及其下0~20 cm土层土壤全氮含量,浅色藻结皮则仅提高了结皮层土层土壤全氮含量,对其下0~20 cm土层土壤全氮含量没有显著影响.生物结皮有机体在湿润状态下才能够进行生理代谢活动,进行相应的固氮作用,并且不同的物种固氮所需最佳水温条件不同[6].生物结皮在湿润后,其固氮活性强烈受制于水分[43].以上研究也表明,深色藻结皮的结皮层田间持水量显著高于浅色藻结皮,因此其固氮持续时间可能较长,故其固氮作用较强,向土壤中输入的氮素较多.氮素在水热交换作用下,向下层输入.而浅色藻结皮持水量较低,固氮作用较弱,因此其固定的氮素仅仅存留在结皮层.同时,生物结皮的存在降低了结皮层pH,但其对结皮层及下层土壤pH并没有显著影响.由于生物结皮某些藻类和地衣能够在湿润状态下进行生理活动,释放有机酸,使土壤pH降低[43],青藏高原黄河源区这一高寒地区气温常年低下,并且降水多以固态或暴雨的形式[44],并且在低温条件下,生物结皮有机体产生的有机酸较少也不易挥发释放出来,因此对土壤pH影响较小.高寒地区生物结皮对土壤养分影响过程及机理还需进一步研究. ...
Response of desert biological soil crusts to alterations in precipitation frequency
1
2004
... 生物结皮是由生长在表层土壤的微生物、藻类、苔藓和地衣等个体微小的生物成分与土壤相互作用形成的复合层[1].其广泛分布于干旱、半干旱区和极地、亚极地等脆弱生态区[2-3].近年来,关于生物结皮生态功能的研究已详细展开,研究表明生物结皮的形成可以改善土壤理化性质,提高土壤稳定性,改善土壤肥力等,对干旱半干旱地区植被恢复与重建具有重要的生态学意义[3-4].目前关于生物结皮生态功能的研究已有很多,但这些研究主要集中在干旱半干旱的沙漠地区[5-14],例如美国科罗拉多沙漠及其西南部的莫哈维沙漠[6-7],以色列南部的内盖夫沙漠[8],阿根廷的蒙特沙漠[9]以及干旱半干旱的西班牙地中海地区[10].国内关于生物结皮的研究也主要集中在沙漠及干旱半干旱地区,如黄土丘陵沟壑区[11-12]、腾格尔沙漠地区[13-14]、西北及新疆沙漠区[15-16].我国青藏高原素有“世界屋脊”、“第三极”之称,其海拔高,气温低,辐射强烈,日照多,黄河源区也源自于此[17].黄河源区是我国青藏高原重要的水源涵养区,也是该区域最主要的畜牧业基地之一.源区内独特的气候条件及其丰富的自然生态资源为该区经济发展及生态平衡发挥了生态安全屏障的作用[17-18].然而,20世纪70年代以来,随着气候的逐渐干旱化和人口、牲畜数量的迅速增加,该区生态环境急剧恶化,大面积植被退化,20世纪末,植被退化面积占源区草地总面积1/3[19].2005年以来,随着三江源地区实施草地“生态保护和建设”过程的具体实施(围栏封育、退牧还草、沙漠化治理、湿地保护等)[19-20],生物结皮开始在该地区逐渐发育.笔者在野外实地考察过程中发现,生物结皮在黄河源区不同退化程度的草地斑块中均有较大面积的分布.黄河源区平均气温-5.38~4.14 ℃,年降水量262.2~772.8 mm[20],在如此相对湿润且温度极低的条件下,生物结皮的发育特征与干热的沙漠地区是否一样,目前还不得而知.生物结皮作为地表特殊的结构层,其属于变水生物[21],在湿润时才能够进行新陈代谢活动[22].而青藏高原高黄河源区属于高原亚寒带半干旱区,仅有冷暖两季,冷季漫长,从每年10月初至次年4月之间地表处于冻结状态[23],降水主要以固态降雪为主,生物结皮呈现休眠状态.在半干旱低温环境条件下,生物结皮的理化性状如何,这一问题也尚不明晰. ...
Temporal and spatial characteristics of permafrost active layer and freezing-thawing process of seasonal permafrost in the source area of the Yellow River
1
2014
... 生物结皮是由生长在表层土壤的微生物、藻类、苔藓和地衣等个体微小的生物成分与土壤相互作用形成的复合层[1].其广泛分布于干旱、半干旱区和极地、亚极地等脆弱生态区[2-3].近年来,关于生物结皮生态功能的研究已详细展开,研究表明生物结皮的形成可以改善土壤理化性质,提高土壤稳定性,改善土壤肥力等,对干旱半干旱地区植被恢复与重建具有重要的生态学意义[3-4].目前关于生物结皮生态功能的研究已有很多,但这些研究主要集中在干旱半干旱的沙漠地区[5-14],例如美国科罗拉多沙漠及其西南部的莫哈维沙漠[6-7],以色列南部的内盖夫沙漠[8],阿根廷的蒙特沙漠[9]以及干旱半干旱的西班牙地中海地区[10].国内关于生物结皮的研究也主要集中在沙漠及干旱半干旱地区,如黄土丘陵沟壑区[11-12]、腾格尔沙漠地区[13-14]、西北及新疆沙漠区[15-16].我国青藏高原素有“世界屋脊”、“第三极”之称,其海拔高,气温低,辐射强烈,日照多,黄河源区也源自于此[17].黄河源区是我国青藏高原重要的水源涵养区,也是该区域最主要的畜牧业基地之一.源区内独特的气候条件及其丰富的自然生态资源为该区经济发展及生态平衡发挥了生态安全屏障的作用[17-18].然而,20世纪70年代以来,随着气候的逐渐干旱化和人口、牲畜数量的迅速增加,该区生态环境急剧恶化,大面积植被退化,20世纪末,植被退化面积占源区草地总面积1/3[19].2005年以来,随着三江源地区实施草地“生态保护和建设”过程的具体实施(围栏封育、退牧还草、沙漠化治理、湿地保护等)[19-20],生物结皮开始在该地区逐渐发育.笔者在野外实地考察过程中发现,生物结皮在黄河源区不同退化程度的草地斑块中均有较大面积的分布.黄河源区平均气温-5.38~4.14 ℃,年降水量262.2~772.8 mm[20],在如此相对湿润且温度极低的条件下,生物结皮的发育特征与干热的沙漠地区是否一样,目前还不得而知.生物结皮作为地表特殊的结构层,其属于变水生物[21],在湿润时才能够进行新陈代谢活动[22].而青藏高原高黄河源区属于高原亚寒带半干旱区,仅有冷暖两季,冷季漫长,从每年10月初至次年4月之间地表处于冻结状态[23],降水主要以固态降雪为主,生物结皮呈现休眠状态.在半干旱低温环境条件下,生物结皮的理化性状如何,这一问题也尚不明晰. ...
黄河源区多年冻土活动层和季节冻土冻融过程时空特征
1
2014
... 生物结皮是由生长在表层土壤的微生物、藻类、苔藓和地衣等个体微小的生物成分与土壤相互作用形成的复合层[1].其广泛分布于干旱、半干旱区和极地、亚极地等脆弱生态区[2-3].近年来,关于生物结皮生态功能的研究已详细展开,研究表明生物结皮的形成可以改善土壤理化性质,提高土壤稳定性,改善土壤肥力等,对干旱半干旱地区植被恢复与重建具有重要的生态学意义[3-4].目前关于生物结皮生态功能的研究已有很多,但这些研究主要集中在干旱半干旱的沙漠地区[5-14],例如美国科罗拉多沙漠及其西南部的莫哈维沙漠[6-7],以色列南部的内盖夫沙漠[8],阿根廷的蒙特沙漠[9]以及干旱半干旱的西班牙地中海地区[10].国内关于生物结皮的研究也主要集中在沙漠及干旱半干旱地区,如黄土丘陵沟壑区[11-12]、腾格尔沙漠地区[13-14]、西北及新疆沙漠区[15-16].我国青藏高原素有“世界屋脊”、“第三极”之称,其海拔高,气温低,辐射强烈,日照多,黄河源区也源自于此[17].黄河源区是我国青藏高原重要的水源涵养区,也是该区域最主要的畜牧业基地之一.源区内独特的气候条件及其丰富的自然生态资源为该区经济发展及生态平衡发挥了生态安全屏障的作用[17-18].然而,20世纪70年代以来,随着气候的逐渐干旱化和人口、牲畜数量的迅速增加,该区生态环境急剧恶化,大面积植被退化,20世纪末,植被退化面积占源区草地总面积1/3[19].2005年以来,随着三江源地区实施草地“生态保护和建设”过程的具体实施(围栏封育、退牧还草、沙漠化治理、湿地保护等)[19-20],生物结皮开始在该地区逐渐发育.笔者在野外实地考察过程中发现,生物结皮在黄河源区不同退化程度的草地斑块中均有较大面积的分布.黄河源区平均气温-5.38~4.14 ℃,年降水量262.2~772.8 mm[20],在如此相对湿润且温度极低的条件下,生物结皮的发育特征与干热的沙漠地区是否一样,目前还不得而知.生物结皮作为地表特殊的结构层,其属于变水生物[21],在湿润时才能够进行新陈代谢活动[22].而青藏高原高黄河源区属于高原亚寒带半干旱区,仅有冷暖两季,冷季漫长,从每年10月初至次年4月之间地表处于冻结状态[23],降水主要以固态降雪为主,生物结皮呈现休眠状态.在半干旱低温环境条件下,生物结皮的理化性状如何,这一问题也尚不明晰. ...
Research on the environmental effect caused by climate change in the source region of the Yellow River
2
2013
... 黄河源区玛多县隶属果洛藏族自治州,地理位置为95°30′~103°30′ E,32°30′~30°30′ N,位于青海省南部,巴颜喀拉山北麓,土地面积12.2×104 km2.该地属于平原地区,平均海拔4 200 m.植被类型以草地为主,约占88%.区域气候类型属于青藏高原亚寒带半干旱半湿润型气候[24-25],昼夜温差大,年平均气温-1.6 ℃;年内降水多集中在5—9月,年内累积降水量407~582 mm;年累积蒸发量为484~584 mm[25].该地区特有的地理位置和地形、地貌、水文、干寒的气候条件决定了该区为季节冻土区,并镶嵌着不连续多年冻土、零星高温多年冻土和季节冻土[26-27].我们的采样观测场地位于玛多县野马滩,位于38°34′ N,98°00′ E,海拔4 320 m,属于季节冻土区,研究场地景观如图1所示. ...
... 生物结皮对土壤理化属性具有显著影响[1,3,14-15,28].关于生物结皮对土壤颗粒组成的影响,在不同的研究区域所得的研究结论存在一定分歧.有些研究结果表明,生物结皮的存在,能够细化土壤颗粒[28,34-35].也有研究表明,生物结皮对土壤颗粒组成并没有显著影响[36].本研究结果表明,由于生物结皮的发育可增加土壤中粉粒含量,5~20 cm土层中粉粒表现为深色藻结皮>浅色藻结皮>裸地,但差异并不显著.来自沙区的研究结果表明,生物结皮能够黏化土壤,捕捉土壤中细颗粒,使得表层土壤中黏粒和粉粒含量呈显著增加趋势.在黄土区的研究结果也表明,随着生物结皮的发育,结皮层土壤细颗相对增加[28].这些研究结果均表明生物结皮对土壤颗粒组成的影响集中在结皮层及其下0~5 cm左右,细颗粒呈现明显的表聚现象[28].而本研究结果发现由于结皮层的覆盖土壤粉粒在20~30 cm土层显著增加,这可能与该区域强烈的冻融过程及水热迁移有关.由于该区域早晚、四季之间温差较大,呈现明显的冻融循环作用[24-25],因此生物结皮层细颗粒可能由于冻融作用及水分迁移随之向下移动,聚集在其下20~30 cm土层.这种迁移作用可能随着水分含量以及发育年限的不同而发生相应的变化.与此同时,本研究结果表明,冻土区生物结皮层土壤容重也显著低于下层土壤,同时深色藻结皮不仅显著降低了自身容重,也影响到了其下0~20 cm土层土壤容重.而浅色藻结皮则对下层土壤容重影响不明显.本研究区海拔高于4 200 m,海拔高、风力大,与沙漠区域不同,生物结皮在形成过程中,并没有显著的捕捉细颗粒的作用,使结皮层土壤细化从而降低结皮层土壤容重.生物结皮在形成过程中,通过菌丝体、藻丝、假根或分泌的多糖来捆绑、黏结土壤颗粒,使得土壤形成许多团聚结构,土壤中孔隙增大,土壤容重降低[34],而深色藻结皮发育年限较长,厚度较厚,其孔隙度高,因此,容重较浅色藻结皮小,田间持水量显著高于浅色藻结皮及裸地. ...
黄河源区气候变化的环境效应研究
2
2013
... 黄河源区玛多县隶属果洛藏族自治州,地理位置为95°30′~103°30′ E,32°30′~30°30′ N,位于青海省南部,巴颜喀拉山北麓,土地面积12.2×104 km2.该地属于平原地区,平均海拔4 200 m.植被类型以草地为主,约占88%.区域气候类型属于青藏高原亚寒带半干旱半湿润型气候[24-25],昼夜温差大,年平均气温-1.6 ℃;年内降水多集中在5—9月,年内累积降水量407~582 mm;年累积蒸发量为484~584 mm[25].该地区特有的地理位置和地形、地貌、水文、干寒的气候条件决定了该区为季节冻土区,并镶嵌着不连续多年冻土、零星高温多年冻土和季节冻土[26-27].我们的采样观测场地位于玛多县野马滩,位于38°34′ N,98°00′ E,海拔4 320 m,属于季节冻土区,研究场地景观如图1所示. ...
... 生物结皮对土壤理化属性具有显著影响[1,3,14-15,28].关于生物结皮对土壤颗粒组成的影响,在不同的研究区域所得的研究结论存在一定分歧.有些研究结果表明,生物结皮的存在,能够细化土壤颗粒[28,34-35].也有研究表明,生物结皮对土壤颗粒组成并没有显著影响[36].本研究结果表明,由于生物结皮的发育可增加土壤中粉粒含量,5~20 cm土层中粉粒表现为深色藻结皮>浅色藻结皮>裸地,但差异并不显著.来自沙区的研究结果表明,生物结皮能够黏化土壤,捕捉土壤中细颗粒,使得表层土壤中黏粒和粉粒含量呈显著增加趋势.在黄土区的研究结果也表明,随着生物结皮的发育,结皮层土壤细颗相对增加[28].这些研究结果均表明生物结皮对土壤颗粒组成的影响集中在结皮层及其下0~5 cm左右,细颗粒呈现明显的表聚现象[28].而本研究结果发现由于结皮层的覆盖土壤粉粒在20~30 cm土层显著增加,这可能与该区域强烈的冻融过程及水热迁移有关.由于该区域早晚、四季之间温差较大,呈现明显的冻融循环作用[24-25],因此生物结皮层细颗粒可能由于冻融作用及水分迁移随之向下移动,聚集在其下20~30 cm土层.这种迁移作用可能随着水分含量以及发育年限的不同而发生相应的变化.与此同时,本研究结果表明,冻土区生物结皮层土壤容重也显著低于下层土壤,同时深色藻结皮不仅显著降低了自身容重,也影响到了其下0~20 cm土层土壤容重.而浅色藻结皮则对下层土壤容重影响不明显.本研究区海拔高于4 200 m,海拔高、风力大,与沙漠区域不同,生物结皮在形成过程中,并没有显著的捕捉细颗粒的作用,使结皮层土壤细化从而降低结皮层土壤容重.生物结皮在形成过程中,通过菌丝体、藻丝、假根或分泌的多糖来捆绑、黏结土壤颗粒,使得土壤形成许多团聚结构,土壤中孔隙增大,土壤容重降低[34],而深色藻结皮发育年限较长,厚度较厚,其孔隙度高,因此,容重较浅色藻结皮小,田间持水量显著高于浅色藻结皮及裸地. ...
Study on the variation characteristics of reference crop evapotranspiration in recent 40 years in source region of Yellow River
3
2013
... 黄河源区玛多县隶属果洛藏族自治州,地理位置为95°30′~103°30′ E,32°30′~30°30′ N,位于青海省南部,巴颜喀拉山北麓,土地面积12.2×104 km2.该地属于平原地区,平均海拔4 200 m.植被类型以草地为主,约占88%.区域气候类型属于青藏高原亚寒带半干旱半湿润型气候[24-25],昼夜温差大,年平均气温-1.6 ℃;年内降水多集中在5—9月,年内累积降水量407~582 mm;年累积蒸发量为484~584 mm[25].该地区特有的地理位置和地形、地貌、水文、干寒的气候条件决定了该区为季节冻土区,并镶嵌着不连续多年冻土、零星高温多年冻土和季节冻土[26-27].我们的采样观测场地位于玛多县野马滩,位于38°34′ N,98°00′ E,海拔4 320 m,属于季节冻土区,研究场地景观如图1所示. ...
... [25].该地区特有的地理位置和地形、地貌、水文、干寒的气候条件决定了该区为季节冻土区,并镶嵌着不连续多年冻土、零星高温多年冻土和季节冻土[26-27].我们的采样观测场地位于玛多县野马滩,位于38°34′ N,98°00′ E,海拔4 320 m,属于季节冻土区,研究场地景观如图1所示. ...
... 生物结皮对土壤理化属性具有显著影响[1,3,14-15,28].关于生物结皮对土壤颗粒组成的影响,在不同的研究区域所得的研究结论存在一定分歧.有些研究结果表明,生物结皮的存在,能够细化土壤颗粒[28,34-35].也有研究表明,生物结皮对土壤颗粒组成并没有显著影响[36].本研究结果表明,由于生物结皮的发育可增加土壤中粉粒含量,5~20 cm土层中粉粒表现为深色藻结皮>浅色藻结皮>裸地,但差异并不显著.来自沙区的研究结果表明,生物结皮能够黏化土壤,捕捉土壤中细颗粒,使得表层土壤中黏粒和粉粒含量呈显著增加趋势.在黄土区的研究结果也表明,随着生物结皮的发育,结皮层土壤细颗相对增加[28].这些研究结果均表明生物结皮对土壤颗粒组成的影响集中在结皮层及其下0~5 cm左右,细颗粒呈现明显的表聚现象[28].而本研究结果发现由于结皮层的覆盖土壤粉粒在20~30 cm土层显著增加,这可能与该区域强烈的冻融过程及水热迁移有关.由于该区域早晚、四季之间温差较大,呈现明显的冻融循环作用[24-25],因此生物结皮层细颗粒可能由于冻融作用及水分迁移随之向下移动,聚集在其下20~30 cm土层.这种迁移作用可能随着水分含量以及发育年限的不同而发生相应的变化.与此同时,本研究结果表明,冻土区生物结皮层土壤容重也显著低于下层土壤,同时深色藻结皮不仅显著降低了自身容重,也影响到了其下0~20 cm土层土壤容重.而浅色藻结皮则对下层土壤容重影响不明显.本研究区海拔高于4 200 m,海拔高、风力大,与沙漠区域不同,生物结皮在形成过程中,并没有显著的捕捉细颗粒的作用,使结皮层土壤细化从而降低结皮层土壤容重.生物结皮在形成过程中,通过菌丝体、藻丝、假根或分泌的多糖来捆绑、黏结土壤颗粒,使得土壤形成许多团聚结构,土壤中孔隙增大,土壤容重降低[34],而深色藻结皮发育年限较长,厚度较厚,其孔隙度高,因此,容重较浅色藻结皮小,田间持水量显著高于浅色藻结皮及裸地. ...
黄河源区近40年参考作物蒸散量变化特征研究
3
2013
... 黄河源区玛多县隶属果洛藏族自治州,地理位置为95°30′~103°30′ E,32°30′~30°30′ N,位于青海省南部,巴颜喀拉山北麓,土地面积12.2×104 km2.该地属于平原地区,平均海拔4 200 m.植被类型以草地为主,约占88%.区域气候类型属于青藏高原亚寒带半干旱半湿润型气候[24-25],昼夜温差大,年平均气温-1.6 ℃;年内降水多集中在5—9月,年内累积降水量407~582 mm;年累积蒸发量为484~584 mm[25].该地区特有的地理位置和地形、地貌、水文、干寒的气候条件决定了该区为季节冻土区,并镶嵌着不连续多年冻土、零星高温多年冻土和季节冻土[26-27].我们的采样观测场地位于玛多县野马滩,位于38°34′ N,98°00′ E,海拔4 320 m,属于季节冻土区,研究场地景观如图1所示. ...
... [25].该地区特有的地理位置和地形、地貌、水文、干寒的气候条件决定了该区为季节冻土区,并镶嵌着不连续多年冻土、零星高温多年冻土和季节冻土[26-27].我们的采样观测场地位于玛多县野马滩,位于38°34′ N,98°00′ E,海拔4 320 m,属于季节冻土区,研究场地景观如图1所示. ...
... 生物结皮对土壤理化属性具有显著影响[1,3,14-15,28].关于生物结皮对土壤颗粒组成的影响,在不同的研究区域所得的研究结论存在一定分歧.有些研究结果表明,生物结皮的存在,能够细化土壤颗粒[28,34-35].也有研究表明,生物结皮对土壤颗粒组成并没有显著影响[36].本研究结果表明,由于生物结皮的发育可增加土壤中粉粒含量,5~20 cm土层中粉粒表现为深色藻结皮>浅色藻结皮>裸地,但差异并不显著.来自沙区的研究结果表明,生物结皮能够黏化土壤,捕捉土壤中细颗粒,使得表层土壤中黏粒和粉粒含量呈显著增加趋势.在黄土区的研究结果也表明,随着生物结皮的发育,结皮层土壤细颗相对增加[28].这些研究结果均表明生物结皮对土壤颗粒组成的影响集中在结皮层及其下0~5 cm左右,细颗粒呈现明显的表聚现象[28].而本研究结果发现由于结皮层的覆盖土壤粉粒在20~30 cm土层显著增加,这可能与该区域强烈的冻融过程及水热迁移有关.由于该区域早晚、四季之间温差较大,呈现明显的冻融循环作用[24-25],因此生物结皮层细颗粒可能由于冻融作用及水分迁移随之向下移动,聚集在其下20~30 cm土层.这种迁移作用可能随着水分含量以及发育年限的不同而发生相应的变化.与此同时,本研究结果表明,冻土区生物结皮层土壤容重也显著低于下层土壤,同时深色藻结皮不仅显著降低了自身容重,也影响到了其下0~20 cm土层土壤容重.而浅色藻结皮则对下层土壤容重影响不明显.本研究区海拔高于4 200 m,海拔高、风力大,与沙漠区域不同,生物结皮在形成过程中,并没有显著的捕捉细颗粒的作用,使结皮层土壤细化从而降低结皮层土壤容重.生物结皮在形成过程中,通过菌丝体、藻丝、假根或分泌的多糖来捆绑、黏结土壤颗粒,使得土壤形成许多团聚结构,土壤中孔隙增大,土壤容重降低[34],而深色藻结皮发育年限较长,厚度较厚,其孔隙度高,因此,容重较浅色藻结皮小,田间持水量显著高于浅色藻结皮及裸地. ...
Thermal regime of warm-dry permafrost in relation to ground surface temperature in the Source Areas of Yangtze and Yellow rivers on the Qinghai-Tibet Plateau, SW China
1
2018
... 黄河源区玛多县隶属果洛藏族自治州,地理位置为95°30′~103°30′ E,32°30′~30°30′ N,位于青海省南部,巴颜喀拉山北麓,土地面积12.2×104 km2.该地属于平原地区,平均海拔4 200 m.植被类型以草地为主,约占88%.区域气候类型属于青藏高原亚寒带半干旱半湿润型气候[24-25],昼夜温差大,年平均气温-1.6 ℃;年内降水多集中在5—9月,年内累积降水量407~582 mm;年累积蒸发量为484~584 mm[25].该地区特有的地理位置和地形、地貌、水文、干寒的气候条件决定了该区为季节冻土区,并镶嵌着不连续多年冻土、零星高温多年冻土和季节冻土[26-27].我们的采样观测场地位于玛多县野马滩,位于38°34′ N,98°00′ E,海拔4 320 m,属于季节冻土区,研究场地景观如图1所示. ...
Spatial distribution characteristics of biological soil crusts and its impact factors and modeling on slope scale in small watershed of northern Shaanxi
1
2015
... 黄河源区玛多县隶属果洛藏族自治州,地理位置为95°30′~103°30′ E,32°30′~30°30′ N,位于青海省南部,巴颜喀拉山北麓,土地面积12.2×104 km2.该地属于平原地区,平均海拔4 200 m.植被类型以草地为主,约占88%.区域气候类型属于青藏高原亚寒带半干旱半湿润型气候[24-25],昼夜温差大,年平均气温-1.6 ℃;年内降水多集中在5—9月,年内累积降水量407~582 mm;年累积蒸发量为484~584 mm[25].该地区特有的地理位置和地形、地貌、水文、干寒的气候条件决定了该区为季节冻土区,并镶嵌着不连续多年冻土、零星高温多年冻土和季节冻土[26-27].我们的采样观测场地位于玛多县野马滩,位于38°34′ N,98°00′ E,海拔4 320 m,属于季节冻土区,研究场地景观如图1所示. ...
陕北小流域坡面尺度生物结皮空间分布特征及其影响因子与建模
1
2015
... 黄河源区玛多县隶属果洛藏族自治州,地理位置为95°30′~103°30′ E,32°30′~30°30′ N,位于青海省南部,巴颜喀拉山北麓,土地面积12.2×104 km2.该地属于平原地区,平均海拔4 200 m.植被类型以草地为主,约占88%.区域气候类型属于青藏高原亚寒带半干旱半湿润型气候[24-25],昼夜温差大,年平均气温-1.6 ℃;年内降水多集中在5—9月,年内累积降水量407~582 mm;年累积蒸发量为484~584 mm[25].该地区特有的地理位置和地形、地貌、水文、干寒的气候条件决定了该区为季节冻土区,并镶嵌着不连续多年冻土、零星高温多年冻土和季节冻土[26-27].我们的采样观测场地位于玛多县野马滩,位于38°34′ N,98°00′ E,海拔4 320 m,属于季节冻土区,研究场地景观如图1所示. ...
Impact of biological soil crust on soil physical properties in the Hilly Loess Plateau region, China
8
2012
... 田间持水量:将原装生物结皮放在具孔小筛上(孔径为1 mm),然后把结皮连同小筛一起,放在盛薄层的托盘上,当结皮充分饱和时,将结皮和小筛取出来,放置在定性滤纸(5层),吸水5 min后,进行结皮湿重的称量,后再将生物结皮的烘干,并称量得到其干重,计算质量含水量,以此作为相对田间持水量[28]. ...
... 生物结皮对土壤理化属性具有显著影响[1,3,14-15,28].关于生物结皮对土壤颗粒组成的影响,在不同的研究区域所得的研究结论存在一定分歧.有些研究结果表明,生物结皮的存在,能够细化土壤颗粒[28,34-35].也有研究表明,生物结皮对土壤颗粒组成并没有显著影响[36].本研究结果表明,由于生物结皮的发育可增加土壤中粉粒含量,5~20 cm土层中粉粒表现为深色藻结皮>浅色藻结皮>裸地,但差异并不显著.来自沙区的研究结果表明,生物结皮能够黏化土壤,捕捉土壤中细颗粒,使得表层土壤中黏粒和粉粒含量呈显著增加趋势.在黄土区的研究结果也表明,随着生物结皮的发育,结皮层土壤细颗相对增加[28].这些研究结果均表明生物结皮对土壤颗粒组成的影响集中在结皮层及其下0~5 cm左右,细颗粒呈现明显的表聚现象[28].而本研究结果发现由于结皮层的覆盖土壤粉粒在20~30 cm土层显著增加,这可能与该区域强烈的冻融过程及水热迁移有关.由于该区域早晚、四季之间温差较大,呈现明显的冻融循环作用[24-25],因此生物结皮层细颗粒可能由于冻融作用及水分迁移随之向下移动,聚集在其下20~30 cm土层.这种迁移作用可能随着水分含量以及发育年限的不同而发生相应的变化.与此同时,本研究结果表明,冻土区生物结皮层土壤容重也显著低于下层土壤,同时深色藻结皮不仅显著降低了自身容重,也影响到了其下0~20 cm土层土壤容重.而浅色藻结皮则对下层土壤容重影响不明显.本研究区海拔高于4 200 m,海拔高、风力大,与沙漠区域不同,生物结皮在形成过程中,并没有显著的捕捉细颗粒的作用,使结皮层土壤细化从而降低结皮层土壤容重.生物结皮在形成过程中,通过菌丝体、藻丝、假根或分泌的多糖来捆绑、黏结土壤颗粒,使得土壤形成许多团聚结构,土壤中孔隙增大,土壤容重降低[34],而深色藻结皮发育年限较长,厚度较厚,其孔隙度高,因此,容重较浅色藻结皮小,田间持水量显著高于浅色藻结皮及裸地. ...
... [28,34-35].也有研究表明,生物结皮对土壤颗粒组成并没有显著影响[36].本研究结果表明,由于生物结皮的发育可增加土壤中粉粒含量,5~20 cm土层中粉粒表现为深色藻结皮>浅色藻结皮>裸地,但差异并不显著.来自沙区的研究结果表明,生物结皮能够黏化土壤,捕捉土壤中细颗粒,使得表层土壤中黏粒和粉粒含量呈显著增加趋势.在黄土区的研究结果也表明,随着生物结皮的发育,结皮层土壤细颗相对增加[28].这些研究结果均表明生物结皮对土壤颗粒组成的影响集中在结皮层及其下0~5 cm左右,细颗粒呈现明显的表聚现象[28].而本研究结果发现由于结皮层的覆盖土壤粉粒在20~30 cm土层显著增加,这可能与该区域强烈的冻融过程及水热迁移有关.由于该区域早晚、四季之间温差较大,呈现明显的冻融循环作用[24-25],因此生物结皮层细颗粒可能由于冻融作用及水分迁移随之向下移动,聚集在其下20~30 cm土层.这种迁移作用可能随着水分含量以及发育年限的不同而发生相应的变化.与此同时,本研究结果表明,冻土区生物结皮层土壤容重也显著低于下层土壤,同时深色藻结皮不仅显著降低了自身容重,也影响到了其下0~20 cm土层土壤容重.而浅色藻结皮则对下层土壤容重影响不明显.本研究区海拔高于4 200 m,海拔高、风力大,与沙漠区域不同,生物结皮在形成过程中,并没有显著的捕捉细颗粒的作用,使结皮层土壤细化从而降低结皮层土壤容重.生物结皮在形成过程中,通过菌丝体、藻丝、假根或分泌的多糖来捆绑、黏结土壤颗粒,使得土壤形成许多团聚结构,土壤中孔隙增大,土壤容重降低[34],而深色藻结皮发育年限较长,厚度较厚,其孔隙度高,因此,容重较浅色藻结皮小,田间持水量显著高于浅色藻结皮及裸地. ...
... [28].这些研究结果均表明生物结皮对土壤颗粒组成的影响集中在结皮层及其下0~5 cm左右,细颗粒呈现明显的表聚现象[28].而本研究结果发现由于结皮层的覆盖土壤粉粒在20~30 cm土层显著增加,这可能与该区域强烈的冻融过程及水热迁移有关.由于该区域早晚、四季之间温差较大,呈现明显的冻融循环作用[24-25],因此生物结皮层细颗粒可能由于冻融作用及水分迁移随之向下移动,聚集在其下20~30 cm土层.这种迁移作用可能随着水分含量以及发育年限的不同而发生相应的变化.与此同时,本研究结果表明,冻土区生物结皮层土壤容重也显著低于下层土壤,同时深色藻结皮不仅显著降低了自身容重,也影响到了其下0~20 cm土层土壤容重.而浅色藻结皮则对下层土壤容重影响不明显.本研究区海拔高于4 200 m,海拔高、风力大,与沙漠区域不同,生物结皮在形成过程中,并没有显著的捕捉细颗粒的作用,使结皮层土壤细化从而降低结皮层土壤容重.生物结皮在形成过程中,通过菌丝体、藻丝、假根或分泌的多糖来捆绑、黏结土壤颗粒,使得土壤形成许多团聚结构,土壤中孔隙增大,土壤容重降低[34],而深色藻结皮发育年限较长,厚度较厚,其孔隙度高,因此,容重较浅色藻结皮小,田间持水量显著高于浅色藻结皮及裸地. ...
... [28].而本研究结果发现由于结皮层的覆盖土壤粉粒在20~30 cm土层显著增加,这可能与该区域强烈的冻融过程及水热迁移有关.由于该区域早晚、四季之间温差较大,呈现明显的冻融循环作用[24-25],因此生物结皮层细颗粒可能由于冻融作用及水分迁移随之向下移动,聚集在其下20~30 cm土层.这种迁移作用可能随着水分含量以及发育年限的不同而发生相应的变化.与此同时,本研究结果表明,冻土区生物结皮层土壤容重也显著低于下层土壤,同时深色藻结皮不仅显著降低了自身容重,也影响到了其下0~20 cm土层土壤容重.而浅色藻结皮则对下层土壤容重影响不明显.本研究区海拔高于4 200 m,海拔高、风力大,与沙漠区域不同,生物结皮在形成过程中,并没有显著的捕捉细颗粒的作用,使结皮层土壤细化从而降低结皮层土壤容重.生物结皮在形成过程中,通过菌丝体、藻丝、假根或分泌的多糖来捆绑、黏结土壤颗粒,使得土壤形成许多团聚结构,土壤中孔隙增大,土壤容重降低[34],而深色藻结皮发育年限较长,厚度较厚,其孔隙度高,因此,容重较浅色藻结皮小,田间持水量显著高于浅色藻结皮及裸地. ...
... 生物结皮的形成,不仅可以改善土壤结构[28,37],也可显著影响表层土壤的养分累积[9,11,15,21].生物结皮中的固氮蓝藻的存在,可将大气中的氮固定从而输入到表层土壤中[38].同时生物结皮中的微生物能够进行固碳作用,可以增加表层土壤有机质含量[1,14,28],增加土壤肥力.虽然生物结皮对土壤养分积累效应已经形成一致结论,但其作用程度在不同区域还有所不同[14,28,40].本研究结果表明该区两种类型生物结皮均显著增加了结皮层及其下0~20 cm土层土壤有机质含量.藻类是生物结皮发育初期的先锋物种,它的形成能够向体外分泌以多糖为主的物质,从而形成生物结皮[41].同时,在给予充足的水分和光照条件下,生物结皮能够进行固碳作用,其固碳潜力主要取决于生物结皮的生物组成,研究已表明藓结皮的固碳能力高于藻结皮[42].该区域两种类型生物结皮,均以藻类结皮为主,因此其固碳能力可能没有显著差异.同时死亡的藻体又成为土壤有机物,为其他异氧微生物提供碳源[1],因此两种类型生物结皮均依靠显著增加了结皮层及其下0~20 cm土层土壤有机质含量.然而,两种生物结皮对土壤全氮含量的影响则有所不同.深色藻结皮的形成同时也提高了结皮层及其下0~20 cm土层土壤全氮含量,浅色藻结皮则仅提高了结皮层土层土壤全氮含量,对其下0~20 cm土层土壤全氮含量没有显著影响.生物结皮有机体在湿润状态下才能够进行生理代谢活动,进行相应的固氮作用,并且不同的物种固氮所需最佳水温条件不同[6].生物结皮在湿润后,其固氮活性强烈受制于水分[43].以上研究也表明,深色藻结皮的结皮层田间持水量显著高于浅色藻结皮,因此其固氮持续时间可能较长,故其固氮作用较强,向土壤中输入的氮素较多.氮素在水热交换作用下,向下层输入.而浅色藻结皮持水量较低,固氮作用较弱,因此其固定的氮素仅仅存留在结皮层.同时,生物结皮的存在降低了结皮层pH,但其对结皮层及下层土壤pH并没有显著影响.由于生物结皮某些藻类和地衣能够在湿润状态下进行生理活动,释放有机酸,使土壤pH降低[43],青藏高原黄河源区这一高寒地区气温常年低下,并且降水多以固态或暴雨的形式[44],并且在低温条件下,生物结皮有机体产生的有机酸较少也不易挥发释放出来,因此对土壤pH影响较小.高寒地区生物结皮对土壤养分影响过程及机理还需进一步研究. ...
... ,28],增加土壤肥力.虽然生物结皮对土壤养分积累效应已经形成一致结论,但其作用程度在不同区域还有所不同[14,28,40].本研究结果表明该区两种类型生物结皮均显著增加了结皮层及其下0~20 cm土层土壤有机质含量.藻类是生物结皮发育初期的先锋物种,它的形成能够向体外分泌以多糖为主的物质,从而形成生物结皮[41].同时,在给予充足的水分和光照条件下,生物结皮能够进行固碳作用,其固碳潜力主要取决于生物结皮的生物组成,研究已表明藓结皮的固碳能力高于藻结皮[42].该区域两种类型生物结皮,均以藻类结皮为主,因此其固碳能力可能没有显著差异.同时死亡的藻体又成为土壤有机物,为其他异氧微生物提供碳源[1],因此两种类型生物结皮均依靠显著增加了结皮层及其下0~20 cm土层土壤有机质含量.然而,两种生物结皮对土壤全氮含量的影响则有所不同.深色藻结皮的形成同时也提高了结皮层及其下0~20 cm土层土壤全氮含量,浅色藻结皮则仅提高了结皮层土层土壤全氮含量,对其下0~20 cm土层土壤全氮含量没有显著影响.生物结皮有机体在湿润状态下才能够进行生理代谢活动,进行相应的固氮作用,并且不同的物种固氮所需最佳水温条件不同[6].生物结皮在湿润后,其固氮活性强烈受制于水分[43].以上研究也表明,深色藻结皮的结皮层田间持水量显著高于浅色藻结皮,因此其固氮持续时间可能较长,故其固氮作用较强,向土壤中输入的氮素较多.氮素在水热交换作用下,向下层输入.而浅色藻结皮持水量较低,固氮作用较弱,因此其固定的氮素仅仅存留在结皮层.同时,生物结皮的存在降低了结皮层pH,但其对结皮层及下层土壤pH并没有显著影响.由于生物结皮某些藻类和地衣能够在湿润状态下进行生理活动,释放有机酸,使土壤pH降低[43],青藏高原黄河源区这一高寒地区气温常年低下,并且降水多以固态或暴雨的形式[44],并且在低温条件下,生物结皮有机体产生的有机酸较少也不易挥发释放出来,因此对土壤pH影响较小.高寒地区生物结皮对土壤养分影响过程及机理还需进一步研究. ...
... ,28,40].本研究结果表明该区两种类型生物结皮均显著增加了结皮层及其下0~20 cm土层土壤有机质含量.藻类是生物结皮发育初期的先锋物种,它的形成能够向体外分泌以多糖为主的物质,从而形成生物结皮[41].同时,在给予充足的水分和光照条件下,生物结皮能够进行固碳作用,其固碳潜力主要取决于生物结皮的生物组成,研究已表明藓结皮的固碳能力高于藻结皮[42].该区域两种类型生物结皮,均以藻类结皮为主,因此其固碳能力可能没有显著差异.同时死亡的藻体又成为土壤有机物,为其他异氧微生物提供碳源[1],因此两种类型生物结皮均依靠显著增加了结皮层及其下0~20 cm土层土壤有机质含量.然而,两种生物结皮对土壤全氮含量的影响则有所不同.深色藻结皮的形成同时也提高了结皮层及其下0~20 cm土层土壤全氮含量,浅色藻结皮则仅提高了结皮层土层土壤全氮含量,对其下0~20 cm土层土壤全氮含量没有显著影响.生物结皮有机体在湿润状态下才能够进行生理代谢活动,进行相应的固氮作用,并且不同的物种固氮所需最佳水温条件不同[6].生物结皮在湿润后,其固氮活性强烈受制于水分[43].以上研究也表明,深色藻结皮的结皮层田间持水量显著高于浅色藻结皮,因此其固氮持续时间可能较长,故其固氮作用较强,向土壤中输入的氮素较多.氮素在水热交换作用下,向下层输入.而浅色藻结皮持水量较低,固氮作用较弱,因此其固定的氮素仅仅存留在结皮层.同时,生物结皮的存在降低了结皮层pH,但其对结皮层及下层土壤pH并没有显著影响.由于生物结皮某些藻类和地衣能够在湿润状态下进行生理活动,释放有机酸,使土壤pH降低[43],青藏高原黄河源区这一高寒地区气温常年低下,并且降水多以固态或暴雨的形式[44],并且在低温条件下,生物结皮有机体产生的有机酸较少也不易挥发释放出来,因此对土壤pH影响较小.高寒地区生物结皮对土壤养分影响过程及机理还需进一步研究. ...
高丽倩, 赵允格, 秦宁强,
8
2012
... 田间持水量:将原装生物结皮放在具孔小筛上(孔径为1 mm),然后把结皮连同小筛一起,放在盛薄层的托盘上,当结皮充分饱和时,将结皮和小筛取出来,放置在定性滤纸(5层),吸水5 min后,进行结皮湿重的称量,后再将生物结皮的烘干,并称量得到其干重,计算质量含水量,以此作为相对田间持水量[28]. ...
... 生物结皮对土壤理化属性具有显著影响[1,3,14-15,28].关于生物结皮对土壤颗粒组成的影响,在不同的研究区域所得的研究结论存在一定分歧.有些研究结果表明,生物结皮的存在,能够细化土壤颗粒[28,34-35].也有研究表明,生物结皮对土壤颗粒组成并没有显著影响[36].本研究结果表明,由于生物结皮的发育可增加土壤中粉粒含量,5~20 cm土层中粉粒表现为深色藻结皮>浅色藻结皮>裸地,但差异并不显著.来自沙区的研究结果表明,生物结皮能够黏化土壤,捕捉土壤中细颗粒,使得表层土壤中黏粒和粉粒含量呈显著增加趋势.在黄土区的研究结果也表明,随着生物结皮的发育,结皮层土壤细颗相对增加[28].这些研究结果均表明生物结皮对土壤颗粒组成的影响集中在结皮层及其下0~5 cm左右,细颗粒呈现明显的表聚现象[28].而本研究结果发现由于结皮层的覆盖土壤粉粒在20~30 cm土层显著增加,这可能与该区域强烈的冻融过程及水热迁移有关.由于该区域早晚、四季之间温差较大,呈现明显的冻融循环作用[24-25],因此生物结皮层细颗粒可能由于冻融作用及水分迁移随之向下移动,聚集在其下20~30 cm土层.这种迁移作用可能随着水分含量以及发育年限的不同而发生相应的变化.与此同时,本研究结果表明,冻土区生物结皮层土壤容重也显著低于下层土壤,同时深色藻结皮不仅显著降低了自身容重,也影响到了其下0~20 cm土层土壤容重.而浅色藻结皮则对下层土壤容重影响不明显.本研究区海拔高于4 200 m,海拔高、风力大,与沙漠区域不同,生物结皮在形成过程中,并没有显著的捕捉细颗粒的作用,使结皮层土壤细化从而降低结皮层土壤容重.生物结皮在形成过程中,通过菌丝体、藻丝、假根或分泌的多糖来捆绑、黏结土壤颗粒,使得土壤形成许多团聚结构,土壤中孔隙增大,土壤容重降低[34],而深色藻结皮发育年限较长,厚度较厚,其孔隙度高,因此,容重较浅色藻结皮小,田间持水量显著高于浅色藻结皮及裸地. ...
... [28,34-35].也有研究表明,生物结皮对土壤颗粒组成并没有显著影响[36].本研究结果表明,由于生物结皮的发育可增加土壤中粉粒含量,5~20 cm土层中粉粒表现为深色藻结皮>浅色藻结皮>裸地,但差异并不显著.来自沙区的研究结果表明,生物结皮能够黏化土壤,捕捉土壤中细颗粒,使得表层土壤中黏粒和粉粒含量呈显著增加趋势.在黄土区的研究结果也表明,随着生物结皮的发育,结皮层土壤细颗相对增加[28].这些研究结果均表明生物结皮对土壤颗粒组成的影响集中在结皮层及其下0~5 cm左右,细颗粒呈现明显的表聚现象[28].而本研究结果发现由于结皮层的覆盖土壤粉粒在20~30 cm土层显著增加,这可能与该区域强烈的冻融过程及水热迁移有关.由于该区域早晚、四季之间温差较大,呈现明显的冻融循环作用[24-25],因此生物结皮层细颗粒可能由于冻融作用及水分迁移随之向下移动,聚集在其下20~30 cm土层.这种迁移作用可能随着水分含量以及发育年限的不同而发生相应的变化.与此同时,本研究结果表明,冻土区生物结皮层土壤容重也显著低于下层土壤,同时深色藻结皮不仅显著降低了自身容重,也影响到了其下0~20 cm土层土壤容重.而浅色藻结皮则对下层土壤容重影响不明显.本研究区海拔高于4 200 m,海拔高、风力大,与沙漠区域不同,生物结皮在形成过程中,并没有显著的捕捉细颗粒的作用,使结皮层土壤细化从而降低结皮层土壤容重.生物结皮在形成过程中,通过菌丝体、藻丝、假根或分泌的多糖来捆绑、黏结土壤颗粒,使得土壤形成许多团聚结构,土壤中孔隙增大,土壤容重降低[34],而深色藻结皮发育年限较长,厚度较厚,其孔隙度高,因此,容重较浅色藻结皮小,田间持水量显著高于浅色藻结皮及裸地. ...
... [28].这些研究结果均表明生物结皮对土壤颗粒组成的影响集中在结皮层及其下0~5 cm左右,细颗粒呈现明显的表聚现象[28].而本研究结果发现由于结皮层的覆盖土壤粉粒在20~30 cm土层显著增加,这可能与该区域强烈的冻融过程及水热迁移有关.由于该区域早晚、四季之间温差较大,呈现明显的冻融循环作用[24-25],因此生物结皮层细颗粒可能由于冻融作用及水分迁移随之向下移动,聚集在其下20~30 cm土层.这种迁移作用可能随着水分含量以及发育年限的不同而发生相应的变化.与此同时,本研究结果表明,冻土区生物结皮层土壤容重也显著低于下层土壤,同时深色藻结皮不仅显著降低了自身容重,也影响到了其下0~20 cm土层土壤容重.而浅色藻结皮则对下层土壤容重影响不明显.本研究区海拔高于4 200 m,海拔高、风力大,与沙漠区域不同,生物结皮在形成过程中,并没有显著的捕捉细颗粒的作用,使结皮层土壤细化从而降低结皮层土壤容重.生物结皮在形成过程中,通过菌丝体、藻丝、假根或分泌的多糖来捆绑、黏结土壤颗粒,使得土壤形成许多团聚结构,土壤中孔隙增大,土壤容重降低[34],而深色藻结皮发育年限较长,厚度较厚,其孔隙度高,因此,容重较浅色藻结皮小,田间持水量显著高于浅色藻结皮及裸地. ...
... [28].而本研究结果发现由于结皮层的覆盖土壤粉粒在20~30 cm土层显著增加,这可能与该区域强烈的冻融过程及水热迁移有关.由于该区域早晚、四季之间温差较大,呈现明显的冻融循环作用[24-25],因此生物结皮层细颗粒可能由于冻融作用及水分迁移随之向下移动,聚集在其下20~30 cm土层.这种迁移作用可能随着水分含量以及发育年限的不同而发生相应的变化.与此同时,本研究结果表明,冻土区生物结皮层土壤容重也显著低于下层土壤,同时深色藻结皮不仅显著降低了自身容重,也影响到了其下0~20 cm土层土壤容重.而浅色藻结皮则对下层土壤容重影响不明显.本研究区海拔高于4 200 m,海拔高、风力大,与沙漠区域不同,生物结皮在形成过程中,并没有显著的捕捉细颗粒的作用,使结皮层土壤细化从而降低结皮层土壤容重.生物结皮在形成过程中,通过菌丝体、藻丝、假根或分泌的多糖来捆绑、黏结土壤颗粒,使得土壤形成许多团聚结构,土壤中孔隙增大,土壤容重降低[34],而深色藻结皮发育年限较长,厚度较厚,其孔隙度高,因此,容重较浅色藻结皮小,田间持水量显著高于浅色藻结皮及裸地. ...
... 生物结皮的形成,不仅可以改善土壤结构[28,37],也可显著影响表层土壤的养分累积[9,11,15,21].生物结皮中的固氮蓝藻的存在,可将大气中的氮固定从而输入到表层土壤中[38].同时生物结皮中的微生物能够进行固碳作用,可以增加表层土壤有机质含量[1,14,28],增加土壤肥力.虽然生物结皮对土壤养分积累效应已经形成一致结论,但其作用程度在不同区域还有所不同[14,28,40].本研究结果表明该区两种类型生物结皮均显著增加了结皮层及其下0~20 cm土层土壤有机质含量.藻类是生物结皮发育初期的先锋物种,它的形成能够向体外分泌以多糖为主的物质,从而形成生物结皮[41].同时,在给予充足的水分和光照条件下,生物结皮能够进行固碳作用,其固碳潜力主要取决于生物结皮的生物组成,研究已表明藓结皮的固碳能力高于藻结皮[42].该区域两种类型生物结皮,均以藻类结皮为主,因此其固碳能力可能没有显著差异.同时死亡的藻体又成为土壤有机物,为其他异氧微生物提供碳源[1],因此两种类型生物结皮均依靠显著增加了结皮层及其下0~20 cm土层土壤有机质含量.然而,两种生物结皮对土壤全氮含量的影响则有所不同.深色藻结皮的形成同时也提高了结皮层及其下0~20 cm土层土壤全氮含量,浅色藻结皮则仅提高了结皮层土层土壤全氮含量,对其下0~20 cm土层土壤全氮含量没有显著影响.生物结皮有机体在湿润状态下才能够进行生理代谢活动,进行相应的固氮作用,并且不同的物种固氮所需最佳水温条件不同[6].生物结皮在湿润后,其固氮活性强烈受制于水分[43].以上研究也表明,深色藻结皮的结皮层田间持水量显著高于浅色藻结皮,因此其固氮持续时间可能较长,故其固氮作用较强,向土壤中输入的氮素较多.氮素在水热交换作用下,向下层输入.而浅色藻结皮持水量较低,固氮作用较弱,因此其固定的氮素仅仅存留在结皮层.同时,生物结皮的存在降低了结皮层pH,但其对结皮层及下层土壤pH并没有显著影响.由于生物结皮某些藻类和地衣能够在湿润状态下进行生理活动,释放有机酸,使土壤pH降低[43],青藏高原黄河源区这一高寒地区气温常年低下,并且降水多以固态或暴雨的形式[44],并且在低温条件下,生物结皮有机体产生的有机酸较少也不易挥发释放出来,因此对土壤pH影响较小.高寒地区生物结皮对土壤养分影响过程及机理还需进一步研究. ...
... ,28],增加土壤肥力.虽然生物结皮对土壤养分积累效应已经形成一致结论,但其作用程度在不同区域还有所不同[14,28,40].本研究结果表明该区两种类型生物结皮均显著增加了结皮层及其下0~20 cm土层土壤有机质含量.藻类是生物结皮发育初期的先锋物种,它的形成能够向体外分泌以多糖为主的物质,从而形成生物结皮[41].同时,在给予充足的水分和光照条件下,生物结皮能够进行固碳作用,其固碳潜力主要取决于生物结皮的生物组成,研究已表明藓结皮的固碳能力高于藻结皮[42].该区域两种类型生物结皮,均以藻类结皮为主,因此其固碳能力可能没有显著差异.同时死亡的藻体又成为土壤有机物,为其他异氧微生物提供碳源[1],因此两种类型生物结皮均依靠显著增加了结皮层及其下0~20 cm土层土壤有机质含量.然而,两种生物结皮对土壤全氮含量的影响则有所不同.深色藻结皮的形成同时也提高了结皮层及其下0~20 cm土层土壤全氮含量,浅色藻结皮则仅提高了结皮层土层土壤全氮含量,对其下0~20 cm土层土壤全氮含量没有显著影响.生物结皮有机体在湿润状态下才能够进行生理代谢活动,进行相应的固氮作用,并且不同的物种固氮所需最佳水温条件不同[6].生物结皮在湿润后,其固氮活性强烈受制于水分[43].以上研究也表明,深色藻结皮的结皮层田间持水量显著高于浅色藻结皮,因此其固氮持续时间可能较长,故其固氮作用较强,向土壤中输入的氮素较多.氮素在水热交换作用下,向下层输入.而浅色藻结皮持水量较低,固氮作用较弱,因此其固定的氮素仅仅存留在结皮层.同时,生物结皮的存在降低了结皮层pH,但其对结皮层及下层土壤pH并没有显著影响.由于生物结皮某些藻类和地衣能够在湿润状态下进行生理活动,释放有机酸,使土壤pH降低[43],青藏高原黄河源区这一高寒地区气温常年低下,并且降水多以固态或暴雨的形式[44],并且在低温条件下,生物结皮有机体产生的有机酸较少也不易挥发释放出来,因此对土壤pH影响较小.高寒地区生物结皮对土壤养分影响过程及机理还需进一步研究. ...
... ,28,40].本研究结果表明该区两种类型生物结皮均显著增加了结皮层及其下0~20 cm土层土壤有机质含量.藻类是生物结皮发育初期的先锋物种,它的形成能够向体外分泌以多糖为主的物质,从而形成生物结皮[41].同时,在给予充足的水分和光照条件下,生物结皮能够进行固碳作用,其固碳潜力主要取决于生物结皮的生物组成,研究已表明藓结皮的固碳能力高于藻结皮[42].该区域两种类型生物结皮,均以藻类结皮为主,因此其固碳能力可能没有显著差异.同时死亡的藻体又成为土壤有机物,为其他异氧微生物提供碳源[1],因此两种类型生物结皮均依靠显著增加了结皮层及其下0~20 cm土层土壤有机质含量.然而,两种生物结皮对土壤全氮含量的影响则有所不同.深色藻结皮的形成同时也提高了结皮层及其下0~20 cm土层土壤全氮含量,浅色藻结皮则仅提高了结皮层土层土壤全氮含量,对其下0~20 cm土层土壤全氮含量没有显著影响.生物结皮有机体在湿润状态下才能够进行生理代谢活动,进行相应的固氮作用,并且不同的物种固氮所需最佳水温条件不同[6].生物结皮在湿润后,其固氮活性强烈受制于水分[43].以上研究也表明,深色藻结皮的结皮层田间持水量显著高于浅色藻结皮,因此其固氮持续时间可能较长,故其固氮作用较强,向土壤中输入的氮素较多.氮素在水热交换作用下,向下层输入.而浅色藻结皮持水量较低,固氮作用较弱,因此其固定的氮素仅仅存留在结皮层.同时,生物结皮的存在降低了结皮层pH,但其对结皮层及下层土壤pH并没有显著影响.由于生物结皮某些藻类和地衣能够在湿润状态下进行生理活动,释放有机酸,使土壤pH降低[43],青藏高原黄河源区这一高寒地区气温常年低下,并且降水多以固态或暴雨的形式[44],并且在低温条件下,生物结皮有机体产生的有机酸较少也不易挥发释放出来,因此对土壤pH影响较小.高寒地区生物结皮对土壤养分影响过程及机理还需进一步研究. ...
Determination the bulk density of biological soil crust by the method of coatings
2
2001
... 容重:结皮层容重与生物结皮层下土壤容重的测定采取的方法分别是涂膜法[29]和环刀法. ...
... 生物结皮可以在各种气候条件下广泛发育[1],国内外关于生物结皮的研究主要集中在荒漠地区及干旱半干旱的黄土高原等地区[4-16],关于青藏高原高寒冻土区生物结皮的发育特点鲜有报道.本研究结果表明冻土区生物结皮主要是以藻为主的生物结皮,结皮层厚度大约为6~22 mm.其在厚度上大于干旱的荒漠地区藻类结皮厚度(1~5 mm)[16].这与该地区特殊的生态环境条件密切相关,由于冻土区常年温度低,生物结皮活性较弱,呼吸等生理活性缓慢[4,21],其中积累的物质不易分解,因此,生物结皮层厚度较大.同时,生物结皮的厚度与生物结皮的发育年限密切相关[29].因此,该区域植被退化区域生物结皮(浅色藻结皮)较未退化区生物结皮(深色藻结皮)发育年限短,故其深色藻结皮厚度大于浅色藻结生物结皮小. ...
涂膜法测定黄土结皮容重
2
2001
... 容重:结皮层容重与生物结皮层下土壤容重的测定采取的方法分别是涂膜法[29]和环刀法. ...
... 生物结皮可以在各种气候条件下广泛发育[1],国内外关于生物结皮的研究主要集中在荒漠地区及干旱半干旱的黄土高原等地区[4-16],关于青藏高原高寒冻土区生物结皮的发育特点鲜有报道.本研究结果表明冻土区生物结皮主要是以藻为主的生物结皮,结皮层厚度大约为6~22 mm.其在厚度上大于干旱的荒漠地区藻类结皮厚度(1~5 mm)[16].这与该地区特殊的生态环境条件密切相关,由于冻土区常年温度低,生物结皮活性较弱,呼吸等生理活性缓慢[4,21],其中积累的物质不易分解,因此,生物结皮层厚度较大.同时,生物结皮的厚度与生物结皮的发育年限密切相关[29].因此,该区域植被退化区域生物结皮(浅色藻结皮)较未退化区生物结皮(深色藻结皮)发育年限短,故其深色藻结皮厚度大于浅色藻结生物结皮小. ...
Nitrogen-Tatal
1
2
... 全氮(TN)用开氏法测定[30],有机质(SOC)使用重铬酸钾氧化法测定[31],pH值用电极法测定[32-33]. ...
Total carbon, organic carbon, and organic matter
1
1996
... 全氮(TN)用开氏法测定[30],有机质(SOC)使用重铬酸钾氧化法测定[31],pH值用电极法测定[32-33]. ...
Long-term fencing effects on plant diversity and soil properties in China
1
2014
... 全氮(TN)用开氏法测定[30],有机质(SOC)使用重铬酸钾氧化法测定[31],pH值用电极法测定[32-33]. ...
Land-use conversion and changing soil carbon stocks in China’s grain-for-green program: a synthesis
1
2014
... 全氮(TN)用开氏法测定[30],有机质(SOC)使用重铬酸钾氧化法测定[31],pH值用电极法测定[32-33]. ...
Effects of biological soil crust on soil physiochemical properties in water-wind erosion crisscross region, northern Shaanxi Province, China
2
2007
... 生物结皮对土壤理化属性具有显著影响[1,3,14-15,28].关于生物结皮对土壤颗粒组成的影响,在不同的研究区域所得的研究结论存在一定分歧.有些研究结果表明,生物结皮的存在,能够细化土壤颗粒[28,34-35].也有研究表明,生物结皮对土壤颗粒组成并没有显著影响[36].本研究结果表明,由于生物结皮的发育可增加土壤中粉粒含量,5~20 cm土层中粉粒表现为深色藻结皮>浅色藻结皮>裸地,但差异并不显著.来自沙区的研究结果表明,生物结皮能够黏化土壤,捕捉土壤中细颗粒,使得表层土壤中黏粒和粉粒含量呈显著增加趋势.在黄土区的研究结果也表明,随着生物结皮的发育,结皮层土壤细颗相对增加[28].这些研究结果均表明生物结皮对土壤颗粒组成的影响集中在结皮层及其下0~5 cm左右,细颗粒呈现明显的表聚现象[28].而本研究结果发现由于结皮层的覆盖土壤粉粒在20~30 cm土层显著增加,这可能与该区域强烈的冻融过程及水热迁移有关.由于该区域早晚、四季之间温差较大,呈现明显的冻融循环作用[24-25],因此生物结皮层细颗粒可能由于冻融作用及水分迁移随之向下移动,聚集在其下20~30 cm土层.这种迁移作用可能随着水分含量以及发育年限的不同而发生相应的变化.与此同时,本研究结果表明,冻土区生物结皮层土壤容重也显著低于下层土壤,同时深色藻结皮不仅显著降低了自身容重,也影响到了其下0~20 cm土层土壤容重.而浅色藻结皮则对下层土壤容重影响不明显.本研究区海拔高于4 200 m,海拔高、风力大,与沙漠区域不同,生物结皮在形成过程中,并没有显著的捕捉细颗粒的作用,使结皮层土壤细化从而降低结皮层土壤容重.生物结皮在形成过程中,通过菌丝体、藻丝、假根或分泌的多糖来捆绑、黏结土壤颗粒,使得土壤形成许多团聚结构,土壤中孔隙增大,土壤容重降低[34],而深色藻结皮发育年限较长,厚度较厚,其孔隙度高,因此,容重较浅色藻结皮小,田间持水量显著高于浅色藻结皮及裸地. ...
... [34],而深色藻结皮发育年限较长,厚度较厚,其孔隙度高,因此,容重较浅色藻结皮小,田间持水量显著高于浅色藻结皮及裸地. ...
陕北水蚀风蚀交错区两种生物结皮对土壤理化性质的影响
2
2007
... 生物结皮对土壤理化属性具有显著影响[1,3,14-15,28].关于生物结皮对土壤颗粒组成的影响,在不同的研究区域所得的研究结论存在一定分歧.有些研究结果表明,生物结皮的存在,能够细化土壤颗粒[28,34-35].也有研究表明,生物结皮对土壤颗粒组成并没有显著影响[36].本研究结果表明,由于生物结皮的发育可增加土壤中粉粒含量,5~20 cm土层中粉粒表现为深色藻结皮>浅色藻结皮>裸地,但差异并不显著.来自沙区的研究结果表明,生物结皮能够黏化土壤,捕捉土壤中细颗粒,使得表层土壤中黏粒和粉粒含量呈显著增加趋势.在黄土区的研究结果也表明,随着生物结皮的发育,结皮层土壤细颗相对增加[28].这些研究结果均表明生物结皮对土壤颗粒组成的影响集中在结皮层及其下0~5 cm左右,细颗粒呈现明显的表聚现象[28].而本研究结果发现由于结皮层的覆盖土壤粉粒在20~30 cm土层显著增加,这可能与该区域强烈的冻融过程及水热迁移有关.由于该区域早晚、四季之间温差较大,呈现明显的冻融循环作用[24-25],因此生物结皮层细颗粒可能由于冻融作用及水分迁移随之向下移动,聚集在其下20~30 cm土层.这种迁移作用可能随着水分含量以及发育年限的不同而发生相应的变化.与此同时,本研究结果表明,冻土区生物结皮层土壤容重也显著低于下层土壤,同时深色藻结皮不仅显著降低了自身容重,也影响到了其下0~20 cm土层土壤容重.而浅色藻结皮则对下层土壤容重影响不明显.本研究区海拔高于4 200 m,海拔高、风力大,与沙漠区域不同,生物结皮在形成过程中,并没有显著的捕捉细颗粒的作用,使结皮层土壤细化从而降低结皮层土壤容重.生物结皮在形成过程中,通过菌丝体、藻丝、假根或分泌的多糖来捆绑、黏结土壤颗粒,使得土壤形成许多团聚结构,土壤中孔隙增大,土壤容重降低[34],而深色藻结皮发育年限较长,厚度较厚,其孔隙度高,因此,容重较浅色藻结皮小,田间持水量显著高于浅色藻结皮及裸地. ...
... [34],而深色藻结皮发育年限较长,厚度较厚,其孔隙度高,因此,容重较浅色藻结皮小,田间持水量显著高于浅色藻结皮及裸地. ...
Study on the soil and water conservation function of microbiltic soil crusts on loess area of northern Shaanxi Province
1
2009
... 生物结皮对土壤理化属性具有显著影响[1,3,14-15,28].关于生物结皮对土壤颗粒组成的影响,在不同的研究区域所得的研究结论存在一定分歧.有些研究结果表明,生物结皮的存在,能够细化土壤颗粒[28,34-35].也有研究表明,生物结皮对土壤颗粒组成并没有显著影响[36].本研究结果表明,由于生物结皮的发育可增加土壤中粉粒含量,5~20 cm土层中粉粒表现为深色藻结皮>浅色藻结皮>裸地,但差异并不显著.来自沙区的研究结果表明,生物结皮能够黏化土壤,捕捉土壤中细颗粒,使得表层土壤中黏粒和粉粒含量呈显著增加趋势.在黄土区的研究结果也表明,随着生物结皮的发育,结皮层土壤细颗相对增加[28].这些研究结果均表明生物结皮对土壤颗粒组成的影响集中在结皮层及其下0~5 cm左右,细颗粒呈现明显的表聚现象[28].而本研究结果发现由于结皮层的覆盖土壤粉粒在20~30 cm土层显著增加,这可能与该区域强烈的冻融过程及水热迁移有关.由于该区域早晚、四季之间温差较大,呈现明显的冻融循环作用[24-25],因此生物结皮层细颗粒可能由于冻融作用及水分迁移随之向下移动,聚集在其下20~30 cm土层.这种迁移作用可能随着水分含量以及发育年限的不同而发生相应的变化.与此同时,本研究结果表明,冻土区生物结皮层土壤容重也显著低于下层土壤,同时深色藻结皮不仅显著降低了自身容重,也影响到了其下0~20 cm土层土壤容重.而浅色藻结皮则对下层土壤容重影响不明显.本研究区海拔高于4 200 m,海拔高、风力大,与沙漠区域不同,生物结皮在形成过程中,并没有显著的捕捉细颗粒的作用,使结皮层土壤细化从而降低结皮层土壤容重.生物结皮在形成过程中,通过菌丝体、藻丝、假根或分泌的多糖来捆绑、黏结土壤颗粒,使得土壤形成许多团聚结构,土壤中孔隙增大,土壤容重降低[34],而深色藻结皮发育年限较长,厚度较厚,其孔隙度高,因此,容重较浅色藻结皮小,田间持水量显著高于浅色藻结皮及裸地. ...
陕北黄土区生物土壤结皮水土保持功能研究
1
2009
... 生物结皮对土壤理化属性具有显著影响[1,3,14-15,28].关于生物结皮对土壤颗粒组成的影响,在不同的研究区域所得的研究结论存在一定分歧.有些研究结果表明,生物结皮的存在,能够细化土壤颗粒[28,34-35].也有研究表明,生物结皮对土壤颗粒组成并没有显著影响[36].本研究结果表明,由于生物结皮的发育可增加土壤中粉粒含量,5~20 cm土层中粉粒表现为深色藻结皮>浅色藻结皮>裸地,但差异并不显著.来自沙区的研究结果表明,生物结皮能够黏化土壤,捕捉土壤中细颗粒,使得表层土壤中黏粒和粉粒含量呈显著增加趋势.在黄土区的研究结果也表明,随着生物结皮的发育,结皮层土壤细颗相对增加[28].这些研究结果均表明生物结皮对土壤颗粒组成的影响集中在结皮层及其下0~5 cm左右,细颗粒呈现明显的表聚现象[28].而本研究结果发现由于结皮层的覆盖土壤粉粒在20~30 cm土层显著增加,这可能与该区域强烈的冻融过程及水热迁移有关.由于该区域早晚、四季之间温差较大,呈现明显的冻融循环作用[24-25],因此生物结皮层细颗粒可能由于冻融作用及水分迁移随之向下移动,聚集在其下20~30 cm土层.这种迁移作用可能随着水分含量以及发育年限的不同而发生相应的变化.与此同时,本研究结果表明,冻土区生物结皮层土壤容重也显著低于下层土壤,同时深色藻结皮不仅显著降低了自身容重,也影响到了其下0~20 cm土层土壤容重.而浅色藻结皮则对下层土壤容重影响不明显.本研究区海拔高于4 200 m,海拔高、风力大,与沙漠区域不同,生物结皮在形成过程中,并没有显著的捕捉细颗粒的作用,使结皮层土壤细化从而降低结皮层土壤容重.生物结皮在形成过程中,通过菌丝体、藻丝、假根或分泌的多糖来捆绑、黏结土壤颗粒,使得土壤形成许多团聚结构,土壤中孔隙增大,土壤容重降低[34],而深色藻结皮发育年限较长,厚度较厚,其孔隙度高,因此,容重较浅色藻结皮小,田间持水量显著高于浅色藻结皮及裸地. ...
Biological soil crust development and its topsoil properties in the process of dune stabilization, Inner Mongolia, China
1
2008
... 生物结皮对土壤理化属性具有显著影响[1,3,14-15,28].关于生物结皮对土壤颗粒组成的影响,在不同的研究区域所得的研究结论存在一定分歧.有些研究结果表明,生物结皮的存在,能够细化土壤颗粒[28,34-35].也有研究表明,生物结皮对土壤颗粒组成并没有显著影响[36].本研究结果表明,由于生物结皮的发育可增加土壤中粉粒含量,5~20 cm土层中粉粒表现为深色藻结皮>浅色藻结皮>裸地,但差异并不显著.来自沙区的研究结果表明,生物结皮能够黏化土壤,捕捉土壤中细颗粒,使得表层土壤中黏粒和粉粒含量呈显著增加趋势.在黄土区的研究结果也表明,随着生物结皮的发育,结皮层土壤细颗相对增加[28].这些研究结果均表明生物结皮对土壤颗粒组成的影响集中在结皮层及其下0~5 cm左右,细颗粒呈现明显的表聚现象[28].而本研究结果发现由于结皮层的覆盖土壤粉粒在20~30 cm土层显著增加,这可能与该区域强烈的冻融过程及水热迁移有关.由于该区域早晚、四季之间温差较大,呈现明显的冻融循环作用[24-25],因此生物结皮层细颗粒可能由于冻融作用及水分迁移随之向下移动,聚集在其下20~30 cm土层.这种迁移作用可能随着水分含量以及发育年限的不同而发生相应的变化.与此同时,本研究结果表明,冻土区生物结皮层土壤容重也显著低于下层土壤,同时深色藻结皮不仅显著降低了自身容重,也影响到了其下0~20 cm土层土壤容重.而浅色藻结皮则对下层土壤容重影响不明显.本研究区海拔高于4 200 m,海拔高、风力大,与沙漠区域不同,生物结皮在形成过程中,并没有显著的捕捉细颗粒的作用,使结皮层土壤细化从而降低结皮层土壤容重.生物结皮在形成过程中,通过菌丝体、藻丝、假根或分泌的多糖来捆绑、黏结土壤颗粒,使得土壤形成许多团聚结构,土壤中孔隙增大,土壤容重降低[34],而深色藻结皮发育年限较长,厚度较厚,其孔隙度高,因此,容重较浅色藻结皮小,田间持水量显著高于浅色藻结皮及裸地. ...
1
2003
... 生物结皮的形成,不仅可以改善土壤结构[28,37],也可显著影响表层土壤的养分累积[9,11,15,21].生物结皮中的固氮蓝藻的存在,可将大气中的氮固定从而输入到表层土壤中[38].同时生物结皮中的微生物能够进行固碳作用,可以增加表层土壤有机质含量[1,14,28],增加土壤肥力.虽然生物结皮对土壤养分积累效应已经形成一致结论,但其作用程度在不同区域还有所不同[14,28,40].本研究结果表明该区两种类型生物结皮均显著增加了结皮层及其下0~20 cm土层土壤有机质含量.藻类是生物结皮发育初期的先锋物种,它的形成能够向体外分泌以多糖为主的物质,从而形成生物结皮[41].同时,在给予充足的水分和光照条件下,生物结皮能够进行固碳作用,其固碳潜力主要取决于生物结皮的生物组成,研究已表明藓结皮的固碳能力高于藻结皮[42].该区域两种类型生物结皮,均以藻类结皮为主,因此其固碳能力可能没有显著差异.同时死亡的藻体又成为土壤有机物,为其他异氧微生物提供碳源[1],因此两种类型生物结皮均依靠显著增加了结皮层及其下0~20 cm土层土壤有机质含量.然而,两种生物结皮对土壤全氮含量的影响则有所不同.深色藻结皮的形成同时也提高了结皮层及其下0~20 cm土层土壤全氮含量,浅色藻结皮则仅提高了结皮层土层土壤全氮含量,对其下0~20 cm土层土壤全氮含量没有显著影响.生物结皮有机体在湿润状态下才能够进行生理代谢活动,进行相应的固氮作用,并且不同的物种固氮所需最佳水温条件不同[6].生物结皮在湿润后,其固氮活性强烈受制于水分[43].以上研究也表明,深色藻结皮的结皮层田间持水量显著高于浅色藻结皮,因此其固氮持续时间可能较长,故其固氮作用较强,向土壤中输入的氮素较多.氮素在水热交换作用下,向下层输入.而浅色藻结皮持水量较低,固氮作用较弱,因此其固定的氮素仅仅存留在结皮层.同时,生物结皮的存在降低了结皮层pH,但其对结皮层及下层土壤pH并没有显著影响.由于生物结皮某些藻类和地衣能够在湿润状态下进行生理活动,释放有机酸,使土壤pH降低[43],青藏高原黄河源区这一高寒地区气温常年低下,并且降水多以固态或暴雨的形式[44],并且在低温条件下,生物结皮有机体产生的有机酸较少也不易挥发释放出来,因此对土壤pH影响较小.高寒地区生物结皮对土壤养分影响过程及机理还需进一步研究. ...
Nitrogen fixation in biological soil crusts from southeast Utah, USA
1
2002
... 生物结皮的形成,不仅可以改善土壤结构[28,37],也可显著影响表层土壤的养分累积[9,11,15,21].生物结皮中的固氮蓝藻的存在,可将大气中的氮固定从而输入到表层土壤中[38].同时生物结皮中的微生物能够进行固碳作用,可以增加表层土壤有机质含量[1,14,28],增加土壤肥力.虽然生物结皮对土壤养分积累效应已经形成一致结论,但其作用程度在不同区域还有所不同[14,28,40].本研究结果表明该区两种类型生物结皮均显著增加了结皮层及其下0~20 cm土层土壤有机质含量.藻类是生物结皮发育初期的先锋物种,它的形成能够向体外分泌以多糖为主的物质,从而形成生物结皮[41].同时,在给予充足的水分和光照条件下,生物结皮能够进行固碳作用,其固碳潜力主要取决于生物结皮的生物组成,研究已表明藓结皮的固碳能力高于藻结皮[42].该区域两种类型生物结皮,均以藻类结皮为主,因此其固碳能力可能没有显著差异.同时死亡的藻体又成为土壤有机物,为其他异氧微生物提供碳源[1],因此两种类型生物结皮均依靠显著增加了结皮层及其下0~20 cm土层土壤有机质含量.然而,两种生物结皮对土壤全氮含量的影响则有所不同.深色藻结皮的形成同时也提高了结皮层及其下0~20 cm土层土壤全氮含量,浅色藻结皮则仅提高了结皮层土层土壤全氮含量,对其下0~20 cm土层土壤全氮含量没有显著影响.生物结皮有机体在湿润状态下才能够进行生理代谢活动,进行相应的固氮作用,并且不同的物种固氮所需最佳水温条件不同[6].生物结皮在湿润后,其固氮活性强烈受制于水分[43].以上研究也表明,深色藻结皮的结皮层田间持水量显著高于浅色藻结皮,因此其固氮持续时间可能较长,故其固氮作用较强,向土壤中输入的氮素较多.氮素在水热交换作用下,向下层输入.而浅色藻结皮持水量较低,固氮作用较弱,因此其固定的氮素仅仅存留在结皮层.同时,生物结皮的存在降低了结皮层pH,但其对结皮层及下层土壤pH并没有显著影响.由于生物结皮某些藻类和地衣能够在湿润状态下进行生理活动,释放有机酸,使土壤pH降低[43],青藏高原黄河源区这一高寒地区气温常年低下,并且降水多以固态或暴雨的形式[44],并且在低温条件下,生物结皮有机体产生的有机酸较少也不易挥发释放出来,因此对土壤pH影响较小.高寒地区生物结皮对土壤养分影响过程及机理还需进一步研究. ...
Physico-chemical properties of soil microbiotic crusts on Erdos Plateau
0
2004
Influence of cryptogamic soil crusts on accumulation of soil organic matter in Gurbantunggut Desert, northern Xinjiang, China
1
2005
... 生物结皮的形成,不仅可以改善土壤结构[28,37],也可显著影响表层土壤的养分累积[9,11,15,21].生物结皮中的固氮蓝藻的存在,可将大气中的氮固定从而输入到表层土壤中[38].同时生物结皮中的微生物能够进行固碳作用,可以增加表层土壤有机质含量[1,14,28],增加土壤肥力.虽然生物结皮对土壤养分积累效应已经形成一致结论,但其作用程度在不同区域还有所不同[14,28,40].本研究结果表明该区两种类型生物结皮均显著增加了结皮层及其下0~20 cm土层土壤有机质含量.藻类是生物结皮发育初期的先锋物种,它的形成能够向体外分泌以多糖为主的物质,从而形成生物结皮[41].同时,在给予充足的水分和光照条件下,生物结皮能够进行固碳作用,其固碳潜力主要取决于生物结皮的生物组成,研究已表明藓结皮的固碳能力高于藻结皮[42].该区域两种类型生物结皮,均以藻类结皮为主,因此其固碳能力可能没有显著差异.同时死亡的藻体又成为土壤有机物,为其他异氧微生物提供碳源[1],因此两种类型生物结皮均依靠显著增加了结皮层及其下0~20 cm土层土壤有机质含量.然而,两种生物结皮对土壤全氮含量的影响则有所不同.深色藻结皮的形成同时也提高了结皮层及其下0~20 cm土层土壤全氮含量,浅色藻结皮则仅提高了结皮层土层土壤全氮含量,对其下0~20 cm土层土壤全氮含量没有显著影响.生物结皮有机体在湿润状态下才能够进行生理代谢活动,进行相应的固氮作用,并且不同的物种固氮所需最佳水温条件不同[6].生物结皮在湿润后,其固氮活性强烈受制于水分[43].以上研究也表明,深色藻结皮的结皮层田间持水量显著高于浅色藻结皮,因此其固氮持续时间可能较长,故其固氮作用较强,向土壤中输入的氮素较多.氮素在水热交换作用下,向下层输入.而浅色藻结皮持水量较低,固氮作用较弱,因此其固定的氮素仅仅存留在结皮层.同时,生物结皮的存在降低了结皮层pH,但其对结皮层及下层土壤pH并没有显著影响.由于生物结皮某些藻类和地衣能够在湿润状态下进行生理活动,释放有机酸,使土壤pH降低[43],青藏高原黄河源区这一高寒地区气温常年低下,并且降水多以固态或暴雨的形式[44],并且在低温条件下,生物结皮有机体产生的有机酸较少也不易挥发释放出来,因此对土壤pH影响较小.高寒地区生物结皮对土壤养分影响过程及机理还需进一步研究. ...
生物结皮影响下的土壤有机质分异特征
1
2005
... 生物结皮的形成,不仅可以改善土壤结构[28,37],也可显著影响表层土壤的养分累积[9,11,15,21].生物结皮中的固氮蓝藻的存在,可将大气中的氮固定从而输入到表层土壤中[38].同时生物结皮中的微生物能够进行固碳作用,可以增加表层土壤有机质含量[1,14,28],增加土壤肥力.虽然生物结皮对土壤养分积累效应已经形成一致结论,但其作用程度在不同区域还有所不同[14,28,40].本研究结果表明该区两种类型生物结皮均显著增加了结皮层及其下0~20 cm土层土壤有机质含量.藻类是生物结皮发育初期的先锋物种,它的形成能够向体外分泌以多糖为主的物质,从而形成生物结皮[41].同时,在给予充足的水分和光照条件下,生物结皮能够进行固碳作用,其固碳潜力主要取决于生物结皮的生物组成,研究已表明藓结皮的固碳能力高于藻结皮[42].该区域两种类型生物结皮,均以藻类结皮为主,因此其固碳能力可能没有显著差异.同时死亡的藻体又成为土壤有机物,为其他异氧微生物提供碳源[1],因此两种类型生物结皮均依靠显著增加了结皮层及其下0~20 cm土层土壤有机质含量.然而,两种生物结皮对土壤全氮含量的影响则有所不同.深色藻结皮的形成同时也提高了结皮层及其下0~20 cm土层土壤全氮含量,浅色藻结皮则仅提高了结皮层土层土壤全氮含量,对其下0~20 cm土层土壤全氮含量没有显著影响.生物结皮有机体在湿润状态下才能够进行生理代谢活动,进行相应的固氮作用,并且不同的物种固氮所需最佳水温条件不同[6].生物结皮在湿润后,其固氮活性强烈受制于水分[43].以上研究也表明,深色藻结皮的结皮层田间持水量显著高于浅色藻结皮,因此其固氮持续时间可能较长,故其固氮作用较强,向土壤中输入的氮素较多.氮素在水热交换作用下,向下层输入.而浅色藻结皮持水量较低,固氮作用较弱,因此其固定的氮素仅仅存留在结皮层.同时,生物结皮的存在降低了结皮层pH,但其对结皮层及下层土壤pH并没有显著影响.由于生物结皮某些藻类和地衣能够在湿润状态下进行生理活动,释放有机酸,使土壤pH降低[43],青藏高原黄河源区这一高寒地区气温常年低下,并且降水多以固态或暴雨的形式[44],并且在低温条件下,生物结皮有机体产生的有机酸较少也不易挥发释放出来,因此对土壤pH影响较小.高寒地区生物结皮对土壤养分影响过程及机理还需进一步研究. ...
Response of biocrusts photosynthesis environmental factors: apossible explanation of the spatial distribution of biocrusts in the Hilly Loess Plateau region of China
1
2010
... 生物结皮的形成,不仅可以改善土壤结构[28,37],也可显著影响表层土壤的养分累积[9,11,15,21].生物结皮中的固氮蓝藻的存在,可将大气中的氮固定从而输入到表层土壤中[38].同时生物结皮中的微生物能够进行固碳作用,可以增加表层土壤有机质含量[1,14,28],增加土壤肥力.虽然生物结皮对土壤养分积累效应已经形成一致结论,但其作用程度在不同区域还有所不同[14,28,40].本研究结果表明该区两种类型生物结皮均显著增加了结皮层及其下0~20 cm土层土壤有机质含量.藻类是生物结皮发育初期的先锋物种,它的形成能够向体外分泌以多糖为主的物质,从而形成生物结皮[41].同时,在给予充足的水分和光照条件下,生物结皮能够进行固碳作用,其固碳潜力主要取决于生物结皮的生物组成,研究已表明藓结皮的固碳能力高于藻结皮[42].该区域两种类型生物结皮,均以藻类结皮为主,因此其固碳能力可能没有显著差异.同时死亡的藻体又成为土壤有机物,为其他异氧微生物提供碳源[1],因此两种类型生物结皮均依靠显著增加了结皮层及其下0~20 cm土层土壤有机质含量.然而,两种生物结皮对土壤全氮含量的影响则有所不同.深色藻结皮的形成同时也提高了结皮层及其下0~20 cm土层土壤全氮含量,浅色藻结皮则仅提高了结皮层土层土壤全氮含量,对其下0~20 cm土层土壤全氮含量没有显著影响.生物结皮有机体在湿润状态下才能够进行生理代谢活动,进行相应的固氮作用,并且不同的物种固氮所需最佳水温条件不同[6].生物结皮在湿润后,其固氮活性强烈受制于水分[43].以上研究也表明,深色藻结皮的结皮层田间持水量显著高于浅色藻结皮,因此其固氮持续时间可能较长,故其固氮作用较强,向土壤中输入的氮素较多.氮素在水热交换作用下,向下层输入.而浅色藻结皮持水量较低,固氮作用较弱,因此其固定的氮素仅仅存留在结皮层.同时,生物结皮的存在降低了结皮层pH,但其对结皮层及下层土壤pH并没有显著影响.由于生物结皮某些藻类和地衣能够在湿润状态下进行生理活动,释放有机酸,使土壤pH降低[43],青藏高原黄河源区这一高寒地区气温常年低下,并且降水多以固态或暴雨的形式[44],并且在低温条件下,生物结皮有机体产生的有机酸较少也不易挥发释放出来,因此对土壤pH影响较小.高寒地区生物结皮对土壤养分影响过程及机理还需进一步研究. ...
生物结皮光合作用对光温水的响应及其对生物结皮空间分布格局的解译
1
2010
... 生物结皮的形成,不仅可以改善土壤结构[28,37],也可显著影响表层土壤的养分累积[9,11,15,21].生物结皮中的固氮蓝藻的存在,可将大气中的氮固定从而输入到表层土壤中[38].同时生物结皮中的微生物能够进行固碳作用,可以增加表层土壤有机质含量[1,14,28],增加土壤肥力.虽然生物结皮对土壤养分积累效应已经形成一致结论,但其作用程度在不同区域还有所不同[14,28,40].本研究结果表明该区两种类型生物结皮均显著增加了结皮层及其下0~20 cm土层土壤有机质含量.藻类是生物结皮发育初期的先锋物种,它的形成能够向体外分泌以多糖为主的物质,从而形成生物结皮[41].同时,在给予充足的水分和光照条件下,生物结皮能够进行固碳作用,其固碳潜力主要取决于生物结皮的生物组成,研究已表明藓结皮的固碳能力高于藻结皮[42].该区域两种类型生物结皮,均以藻类结皮为主,因此其固碳能力可能没有显著差异.同时死亡的藻体又成为土壤有机物,为其他异氧微生物提供碳源[1],因此两种类型生物结皮均依靠显著增加了结皮层及其下0~20 cm土层土壤有机质含量.然而,两种生物结皮对土壤全氮含量的影响则有所不同.深色藻结皮的形成同时也提高了结皮层及其下0~20 cm土层土壤全氮含量,浅色藻结皮则仅提高了结皮层土层土壤全氮含量,对其下0~20 cm土层土壤全氮含量没有显著影响.生物结皮有机体在湿润状态下才能够进行生理代谢活动,进行相应的固氮作用,并且不同的物种固氮所需最佳水温条件不同[6].生物结皮在湿润后,其固氮活性强烈受制于水分[43].以上研究也表明,深色藻结皮的结皮层田间持水量显著高于浅色藻结皮,因此其固氮持续时间可能较长,故其固氮作用较强,向土壤中输入的氮素较多.氮素在水热交换作用下,向下层输入.而浅色藻结皮持水量较低,固氮作用较弱,因此其固定的氮素仅仅存留在结皮层.同时,生物结皮的存在降低了结皮层pH,但其对结皮层及下层土壤pH并没有显著影响.由于生物结皮某些藻类和地衣能够在湿润状态下进行生理活动,释放有机酸,使土壤pH降低[43],青藏高原黄河源区这一高寒地区气温常年低下,并且降水多以固态或暴雨的形式[44],并且在低温条件下,生物结皮有机体产生的有机酸较少也不易挥发释放出来,因此对土壤pH影响较小.高寒地区生物结皮对土壤养分影响过程及机理还需进一步研究. ...
The nitrogenase activity of biological soil crusts and their responses to environmental factors
1
2011
... 生物结皮的形成,不仅可以改善土壤结构[28,37],也可显著影响表层土壤的养分累积[9,11,15,21].生物结皮中的固氮蓝藻的存在,可将大气中的氮固定从而输入到表层土壤中[38].同时生物结皮中的微生物能够进行固碳作用,可以增加表层土壤有机质含量[1,14,28],增加土壤肥力.虽然生物结皮对土壤养分积累效应已经形成一致结论,但其作用程度在不同区域还有所不同[14,28,40].本研究结果表明该区两种类型生物结皮均显著增加了结皮层及其下0~20 cm土层土壤有机质含量.藻类是生物结皮发育初期的先锋物种,它的形成能够向体外分泌以多糖为主的物质,从而形成生物结皮[41].同时,在给予充足的水分和光照条件下,生物结皮能够进行固碳作用,其固碳潜力主要取决于生物结皮的生物组成,研究已表明藓结皮的固碳能力高于藻结皮[42].该区域两种类型生物结皮,均以藻类结皮为主,因此其固碳能力可能没有显著差异.同时死亡的藻体又成为土壤有机物,为其他异氧微生物提供碳源[1],因此两种类型生物结皮均依靠显著增加了结皮层及其下0~20 cm土层土壤有机质含量.然而,两种生物结皮对土壤全氮含量的影响则有所不同.深色藻结皮的形成同时也提高了结皮层及其下0~20 cm土层土壤全氮含量,浅色藻结皮则仅提高了结皮层土层土壤全氮含量,对其下0~20 cm土层土壤全氮含量没有显著影响.生物结皮有机体在湿润状态下才能够进行生理代谢活动,进行相应的固氮作用,并且不同的物种固氮所需最佳水温条件不同[6].生物结皮在湿润后,其固氮活性强烈受制于水分[43].以上研究也表明,深色藻结皮的结皮层田间持水量显著高于浅色藻结皮,因此其固氮持续时间可能较长,故其固氮作用较强,向土壤中输入的氮素较多.氮素在水热交换作用下,向下层输入.而浅色藻结皮持水量较低,固氮作用较弱,因此其固定的氮素仅仅存留在结皮层.同时,生物结皮的存在降低了结皮层pH,但其对结皮层及下层土壤pH并没有显著影响.由于生物结皮某些藻类和地衣能够在湿润状态下进行生理活动,释放有机酸,使土壤pH降低[43],青藏高原黄河源区这一高寒地区气温常年低下,并且降水多以固态或暴雨的形式[44],并且在低温条件下,生物结皮有机体产生的有机酸较少也不易挥发释放出来,因此对土壤pH影响较小.高寒地区生物结皮对土壤养分影响过程及机理还需进一步研究. ...
不同类型生物结皮固氮活性对环境因子的响应研究
1
2011
... 生物结皮的形成,不仅可以改善土壤结构[28,37],也可显著影响表层土壤的养分累积[9,11,15,21].生物结皮中的固氮蓝藻的存在,可将大气中的氮固定从而输入到表层土壤中[38].同时生物结皮中的微生物能够进行固碳作用,可以增加表层土壤有机质含量[1,14,28],增加土壤肥力.虽然生物结皮对土壤养分积累效应已经形成一致结论,但其作用程度在不同区域还有所不同[14,28,40].本研究结果表明该区两种类型生物结皮均显著增加了结皮层及其下0~20 cm土层土壤有机质含量.藻类是生物结皮发育初期的先锋物种,它的形成能够向体外分泌以多糖为主的物质,从而形成生物结皮[41].同时,在给予充足的水分和光照条件下,生物结皮能够进行固碳作用,其固碳潜力主要取决于生物结皮的生物组成,研究已表明藓结皮的固碳能力高于藻结皮[42].该区域两种类型生物结皮,均以藻类结皮为主,因此其固碳能力可能没有显著差异.同时死亡的藻体又成为土壤有机物,为其他异氧微生物提供碳源[1],因此两种类型生物结皮均依靠显著增加了结皮层及其下0~20 cm土层土壤有机质含量.然而,两种生物结皮对土壤全氮含量的影响则有所不同.深色藻结皮的形成同时也提高了结皮层及其下0~20 cm土层土壤全氮含量,浅色藻结皮则仅提高了结皮层土层土壤全氮含量,对其下0~20 cm土层土壤全氮含量没有显著影响.生物结皮有机体在湿润状态下才能够进行生理代谢活动,进行相应的固氮作用,并且不同的物种固氮所需最佳水温条件不同[6].生物结皮在湿润后,其固氮活性强烈受制于水分[43].以上研究也表明,深色藻结皮的结皮层田间持水量显著高于浅色藻结皮,因此其固氮持续时间可能较长,故其固氮作用较强,向土壤中输入的氮素较多.氮素在水热交换作用下,向下层输入.而浅色藻结皮持水量较低,固氮作用较弱,因此其固定的氮素仅仅存留在结皮层.同时,生物结皮的存在降低了结皮层pH,但其对结皮层及下层土壤pH并没有显著影响.由于生物结皮某些藻类和地衣能够在湿润状态下进行生理活动,释放有机酸,使土壤pH降低[43],青藏高原黄河源区这一高寒地区气温常年低下,并且降水多以固态或暴雨的形式[44],并且在低温条件下,生物结皮有机体产生的有机酸较少也不易挥发释放出来,因此对土壤pH影响较小.高寒地区生物结皮对土壤养分影响过程及机理还需进一步研究. ...
The chemical ecology of cyanobacteria
2
2012
... 生物结皮的形成,不仅可以改善土壤结构[28,37],也可显著影响表层土壤的养分累积[9,11,15,21].生物结皮中的固氮蓝藻的存在,可将大气中的氮固定从而输入到表层土壤中[38].同时生物结皮中的微生物能够进行固碳作用,可以增加表层土壤有机质含量[1,14,28],增加土壤肥力.虽然生物结皮对土壤养分积累效应已经形成一致结论,但其作用程度在不同区域还有所不同[14,28,40].本研究结果表明该区两种类型生物结皮均显著增加了结皮层及其下0~20 cm土层土壤有机质含量.藻类是生物结皮发育初期的先锋物种,它的形成能够向体外分泌以多糖为主的物质,从而形成生物结皮[41].同时,在给予充足的水分和光照条件下,生物结皮能够进行固碳作用,其固碳潜力主要取决于生物结皮的生物组成,研究已表明藓结皮的固碳能力高于藻结皮[42].该区域两种类型生物结皮,均以藻类结皮为主,因此其固碳能力可能没有显著差异.同时死亡的藻体又成为土壤有机物,为其他异氧微生物提供碳源[1],因此两种类型生物结皮均依靠显著增加了结皮层及其下0~20 cm土层土壤有机质含量.然而,两种生物结皮对土壤全氮含量的影响则有所不同.深色藻结皮的形成同时也提高了结皮层及其下0~20 cm土层土壤全氮含量,浅色藻结皮则仅提高了结皮层土层土壤全氮含量,对其下0~20 cm土层土壤全氮含量没有显著影响.生物结皮有机体在湿润状态下才能够进行生理代谢活动,进行相应的固氮作用,并且不同的物种固氮所需最佳水温条件不同[6].生物结皮在湿润后,其固氮活性强烈受制于水分[43].以上研究也表明,深色藻结皮的结皮层田间持水量显著高于浅色藻结皮,因此其固氮持续时间可能较长,故其固氮作用较强,向土壤中输入的氮素较多.氮素在水热交换作用下,向下层输入.而浅色藻结皮持水量较低,固氮作用较弱,因此其固定的氮素仅仅存留在结皮层.同时,生物结皮的存在降低了结皮层pH,但其对结皮层及下层土壤pH并没有显著影响.由于生物结皮某些藻类和地衣能够在湿润状态下进行生理活动,释放有机酸,使土壤pH降低[43],青藏高原黄河源区这一高寒地区气温常年低下,并且降水多以固态或暴雨的形式[44],并且在低温条件下,生物结皮有机体产生的有机酸较少也不易挥发释放出来,因此对土壤pH影响较小.高寒地区生物结皮对土壤养分影响过程及机理还需进一步研究. ...
... [43],青藏高原黄河源区这一高寒地区气温常年低下,并且降水多以固态或暴雨的形式[44],并且在低温条件下,生物结皮有机体产生的有机酸较少也不易挥发释放出来,因此对土壤pH影响较小.高寒地区生物结皮对土壤养分影响过程及机理还需进一步研究. ...
Features and degradation of frozen ground in the sources area of the Yellow River, China
1
2010
... 生物结皮的形成,不仅可以改善土壤结构[28,37],也可显著影响表层土壤的养分累积[9,11,15,21].生物结皮中的固氮蓝藻的存在,可将大气中的氮固定从而输入到表层土壤中[38].同时生物结皮中的微生物能够进行固碳作用,可以增加表层土壤有机质含量[1,14,28],增加土壤肥力.虽然生物结皮对土壤养分积累效应已经形成一致结论,但其作用程度在不同区域还有所不同[14,28,40].本研究结果表明该区两种类型生物结皮均显著增加了结皮层及其下0~20 cm土层土壤有机质含量.藻类是生物结皮发育初期的先锋物种,它的形成能够向体外分泌以多糖为主的物质,从而形成生物结皮[41].同时,在给予充足的水分和光照条件下,生物结皮能够进行固碳作用,其固碳潜力主要取决于生物结皮的生物组成,研究已表明藓结皮的固碳能力高于藻结皮[42].该区域两种类型生物结皮,均以藻类结皮为主,因此其固碳能力可能没有显著差异.同时死亡的藻体又成为土壤有机物,为其他异氧微生物提供碳源[1],因此两种类型生物结皮均依靠显著增加了结皮层及其下0~20 cm土层土壤有机质含量.然而,两种生物结皮对土壤全氮含量的影响则有所不同.深色藻结皮的形成同时也提高了结皮层及其下0~20 cm土层土壤全氮含量,浅色藻结皮则仅提高了结皮层土层土壤全氮含量,对其下0~20 cm土层土壤全氮含量没有显著影响.生物结皮有机体在湿润状态下才能够进行生理代谢活动,进行相应的固氮作用,并且不同的物种固氮所需最佳水温条件不同[6].生物结皮在湿润后,其固氮活性强烈受制于水分[43].以上研究也表明,深色藻结皮的结皮层田间持水量显著高于浅色藻结皮,因此其固氮持续时间可能较长,故其固氮作用较强,向土壤中输入的氮素较多.氮素在水热交换作用下,向下层输入.而浅色藻结皮持水量较低,固氮作用较弱,因此其固定的氮素仅仅存留在结皮层.同时,生物结皮的存在降低了结皮层pH,但其对结皮层及下层土壤pH并没有显著影响.由于生物结皮某些藻类和地衣能够在湿润状态下进行生理活动,释放有机酸,使土壤pH降低[43],青藏高原黄河源区这一高寒地区气温常年低下,并且降水多以固态或暴雨的形式[44],并且在低温条件下,生物结皮有机体产生的有机酸较少也不易挥发释放出来,因此对土壤pH影响较小.高寒地区生物结皮对土壤养分影响过程及机理还需进一步研究. ...
黄河源区冻土特征及退化趋势
1
2010
... 生物结皮的形成,不仅可以改善土壤结构[28,37],也可显著影响表层土壤的养分累积[9,11,15,21].生物结皮中的固氮蓝藻的存在,可将大气中的氮固定从而输入到表层土壤中[38].同时生物结皮中的微生物能够进行固碳作用,可以增加表层土壤有机质含量[1,14,28],增加土壤肥力.虽然生物结皮对土壤养分积累效应已经形成一致结论,但其作用程度在不同区域还有所不同[14,28,40].本研究结果表明该区两种类型生物结皮均显著增加了结皮层及其下0~20 cm土层土壤有机质含量.藻类是生物结皮发育初期的先锋物种,它的形成能够向体外分泌以多糖为主的物质,从而形成生物结皮[41].同时,在给予充足的水分和光照条件下,生物结皮能够进行固碳作用,其固碳潜力主要取决于生物结皮的生物组成,研究已表明藓结皮的固碳能力高于藻结皮[42].该区域两种类型生物结皮,均以藻类结皮为主,因此其固碳能力可能没有显著差异.同时死亡的藻体又成为土壤有机物,为其他异氧微生物提供碳源[1],因此两种类型生物结皮均依靠显著增加了结皮层及其下0~20 cm土层土壤有机质含量.然而,两种生物结皮对土壤全氮含量的影响则有所不同.深色藻结皮的形成同时也提高了结皮层及其下0~20 cm土层土壤全氮含量,浅色藻结皮则仅提高了结皮层土层土壤全氮含量,对其下0~20 cm土层土壤全氮含量没有显著影响.生物结皮有机体在湿润状态下才能够进行生理代谢活动,进行相应的固氮作用,并且不同的物种固氮所需最佳水温条件不同[6].生物结皮在湿润后,其固氮活性强烈受制于水分[43].以上研究也表明,深色藻结皮的结皮层田间持水量显著高于浅色藻结皮,因此其固氮持续时间可能较长,故其固氮作用较强,向土壤中输入的氮素较多.氮素在水热交换作用下,向下层输入.而浅色藻结皮持水量较低,固氮作用较弱,因此其固定的氮素仅仅存留在结皮层.同时,生物结皮的存在降低了结皮层pH,但其对结皮层及下层土壤pH并没有显著影响.由于生物结皮某些藻类和地衣能够在湿润状态下进行生理活动,释放有机酸,使土壤pH降低[43],青藏高原黄河源区这一高寒地区气温常年低下,并且降水多以固态或暴雨的形式[44],并且在低温条件下,生物结皮有机体产生的有机酸较少也不易挥发释放出来,因此对土壤pH影响较小.高寒地区生物结皮对土壤养分影响过程及机理还需进一步研究. ...


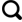




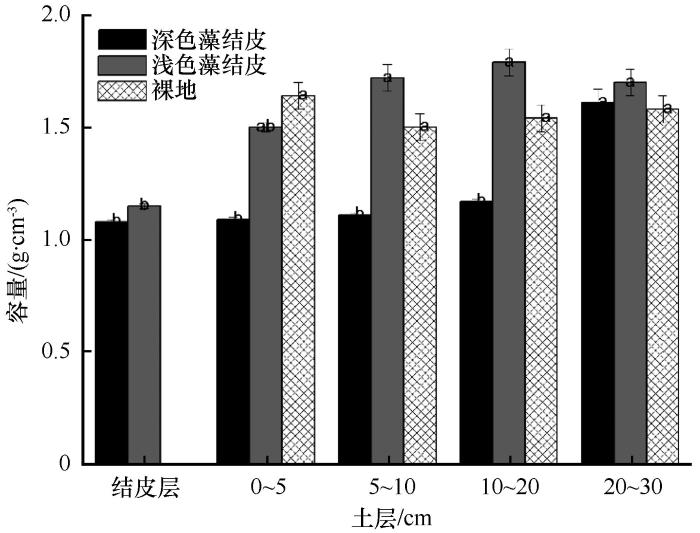
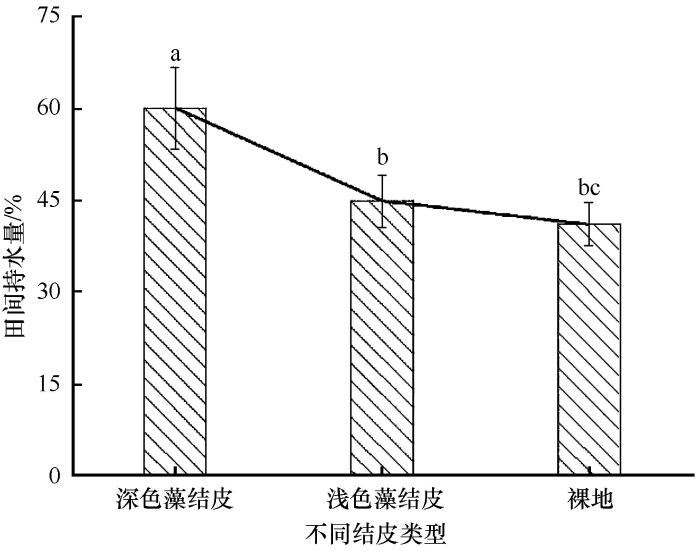

 甘公网安备 62010202000676号
甘公网安备 62010202000676号
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}