The toxicology of mercury and its chemical compounds
3
2006
... 汞是一种全球性污染物,由于元素汞具有较高蒸汽压,在大气中的停留时间长(0.5~2年),是唯一能以气态形式进行远距离传输的重金属元素[1].人类活动排放的汞污染物可以通过大气环流迁移到地球偏远地区,并通过干、湿沉降进入陆地和水生生态系统,最终富集于冰川、植被、水体和土壤等地表环境之中[2-3].当今人为活动使大气中的总汞浓度已超出自然水平的4.5倍[4],大气沉降到地表环境之中的无机汞可以通过生物和非生物甲基化作用转化为毒性更大的甲基汞[5-7].甲基汞具有很强的神经毒性,它在生态系统食物链中具有显著的生物放大和生物积累特性[1,8].2013年,联合国环境规划署通过了旨在减排汞污染物的国际公约—《关于汞的水俣公约》,以遏制人为汞污染向环境排放输入,协议的主要目标之一是最大程度上降低与甲基汞接触相关的健康风险. ...
... [1,8].2013年,联合国环境规划署通过了旨在减排汞污染物的国际公约—《关于汞的水俣公约》,以遏制人为汞污染向环境排放输入,协议的主要目标之一是最大程度上降低与甲基汞接触相关的健康风险. ...
... 不仅如此,气候变化会刺激冻土、融雪和湿地环境微生物汞甲基化,同时冻土融化的增强、降水的改变以及融雪导致地表水流量的增加,促使甲基汞向下游生态系统输送[22-25].冰川、冰盖和海冰的减少也会对冰冻圈汞生物地球化学循环产生重大影响[26-27].以上事实都表明,冰川、冻土、积雪和海冰等冰冻圈介质融化将成为陆地和海洋生态系统汞的一个重要来源,越来越多的证据亦指出冰冻圈消融向下游生态系统输送的汞总量巨大,这无疑会加剧甲基汞暴露风险[24-25,28-31].甲基汞可以在生物体内积累,进而对高营养级生物甚至人类产生潜在危害[1,8].已有研究报道北极生物体内有高水平的汞含量(如鸟类和鱼类),Franklin海湾冰下鳕鱼的总汞浓度为0.38 μg·g-1,其中80%为甲基汞[32].有研究报道北极地区居民每周从海洋食品摄入汞约846 μg,远超过世界卫生组织限定的阈值[33]. ...
Long-range atmospheric transport of mercury to ecosystems, and the importance of anthropogenic emissions: a critical review and evaluation of the published evidence
1
1997
... 汞是一种全球性污染物,由于元素汞具有较高蒸汽压,在大气中的停留时间长(0.5~2年),是唯一能以气态形式进行远距离传输的重金属元素[1].人类活动排放的汞污染物可以通过大气环流迁移到地球偏远地区,并通过干、湿沉降进入陆地和水生生态系统,最终富集于冰川、植被、水体和土壤等地表环境之中[2-3].当今人为活动使大气中的总汞浓度已超出自然水平的4.5倍[4],大气沉降到地表环境之中的无机汞可以通过生物和非生物甲基化作用转化为毒性更大的甲基汞[5-7].甲基汞具有很强的神经毒性,它在生态系统食物链中具有显著的生物放大和生物积累特性[1,8].2013年,联合国环境规划署通过了旨在减排汞污染物的国际公约—《关于汞的水俣公约》,以遏制人为汞污染向环境排放输入,协议的主要目标之一是最大程度上降低与甲基汞接触相关的健康风险. ...
The case for atmospheric mercury contamination in remote areas
2
1998
... 汞是一种全球性污染物,由于元素汞具有较高蒸汽压,在大气中的停留时间长(0.5~2年),是唯一能以气态形式进行远距离传输的重金属元素[1].人类活动排放的汞污染物可以通过大气环流迁移到地球偏远地区,并通过干、湿沉降进入陆地和水生生态系统,最终富集于冰川、植被、水体和土壤等地表环境之中[2-3].当今人为活动使大气中的总汞浓度已超出自然水平的4.5倍[4],大气沉降到地表环境之中的无机汞可以通过生物和非生物甲基化作用转化为毒性更大的甲基汞[5-7].甲基汞具有很强的神经毒性,它在生态系统食物链中具有显著的生物放大和生物积累特性[1,8].2013年,联合国环境规划署通过了旨在减排汞污染物的国际公约—《关于汞的水俣公约》,以遏制人为汞污染向环境排放输入,协议的主要目标之一是最大程度上降低与甲基汞接触相关的健康风险. ...
... 冰冻圈甲基汞富集作用同样对生活在该区域的人类健康构成重大威胁.北极地区居住着土著居民(如因纽特人),他们依赖于捕获鱼类和海洋哺乳动物为食,通过饮食摄入是当地居民暴露甲基汞的主要途径[3,155-157].北极因纽特人原住民由于常年捕食海洋生物,人体内的甲基汞含量都远超过了毒理学阈值[104,158].由于甲基汞对成人和儿童的毒性效应有所不同,处于发育中的儿童中枢神经系统比成年人对甲基汞更为敏感,而孕妇长期处于低剂量甲基汞暴露将会对婴儿的智力发育产生严重影响[33].研究表明,北极一些地区50%以上的母亲和育龄妇女的血液中的汞含量已超过美国环境保护署建议的标准浓度(5.8 μg·L-1),且在格陵兰岛部分地区近乎90%的育龄妇女血液中的汞含量都超标,这些都表明我们迫切需要重视冰冻圈甲基汞暴露风险[33,157-158]. ...
Arctic Monitoring and Assessment Programme 2018: technical background assessment for the 2018 Global Mercury Assessment
2
2018
... 汞是一种全球性污染物,由于元素汞具有较高蒸汽压,在大气中的停留时间长(0.5~2年),是唯一能以气态形式进行远距离传输的重金属元素[1].人类活动排放的汞污染物可以通过大气环流迁移到地球偏远地区,并通过干、湿沉降进入陆地和水生生态系统,最终富集于冰川、植被、水体和土壤等地表环境之中[2-3].当今人为活动使大气中的总汞浓度已超出自然水平的4.5倍[4],大气沉降到地表环境之中的无机汞可以通过生物和非生物甲基化作用转化为毒性更大的甲基汞[5-7].甲基汞具有很强的神经毒性,它在生态系统食物链中具有显著的生物放大和生物积累特性[1,8].2013年,联合国环境规划署通过了旨在减排汞污染物的国际公约—《关于汞的水俣公约》,以遏制人为汞污染向环境排放输入,协议的主要目标之一是最大程度上降低与甲基汞接触相关的健康风险. ...
... 冰冻圈在地球上覆盖范围超过9.6×107 km2,包括冰盖、山地冰川、多年冻土、积雪、海冰、冰架等,它们主要分布在南北极和青藏高原(“第三极”)[9-10].在人类活动深刻影响自然环境的背景下,远离人类活动区的冰冻圈亦受到可全球传输的污染物影响.人类活动释放的汞经长距离传输进入偏远地区,封存于冰冻圈,使冰川、冻土等冰冻圈要素成为汞的临时储库[11-17].Schuster等[15]估算了整个北半球冻土区的汞储量约为(1 656±962) Gg,Mu等[17]估算出青藏高原活动层储存汞约16.6 Gg.沉积于北极冰川中的汞总量约为(2 415±22) Gg,Huang等[18-19]估算出中国西部冰川区每年冰尘中汞总量为34.3 kg.然而,随着全球气候持续变暖,冰冻圈要素快速退缩变化,这将导致储存于冰冻圈中的污染物发生“二次释放”,将对生态环境和人体健康产生不利影响[4,9].据估计,格陵兰冰盖西南部融水径流中汞通量约为3~42.4 kg·a-1[20].Schaefer等[21]模拟了RCP8.5排放情景下冻土退化导致Yukon流域汞输出将增加一倍(约为11 Mg·a-1).Dastoord等[18]估算出亚北极地区(60° N以北)河流向北冰洋输出溶解汞和颗粒汞总量约为(41±4) Mg·a-1,主要来源于季节性积雪和多年冻土区. ...
Organomercurials. Their formation and pathways in the environment
2
2010
... 汞是一种全球性污染物,由于元素汞具有较高蒸汽压,在大气中的停留时间长(0.5~2年),是唯一能以气态形式进行远距离传输的重金属元素[1].人类活动排放的汞污染物可以通过大气环流迁移到地球偏远地区,并通过干、湿沉降进入陆地和水生生态系统,最终富集于冰川、植被、水体和土壤等地表环境之中[2-3].当今人为活动使大气中的总汞浓度已超出自然水平的4.5倍[4],大气沉降到地表环境之中的无机汞可以通过生物和非生物甲基化作用转化为毒性更大的甲基汞[5-7].甲基汞具有很强的神经毒性,它在生态系统食物链中具有显著的生物放大和生物积累特性[1,8].2013年,联合国环境规划署通过了旨在减排汞污染物的国际公约—《关于汞的水俣公约》,以遏制人为汞污染向环境排放输入,协议的主要目标之一是最大程度上降低与甲基汞接触相关的健康风险. ...
... 汞的甲基化包括生物和非生物两种途径[5,37-40].在自然条件下汞的非生物甲基化效率非常低,通常认为环境中的甲基汞是由微生物主导产生[38-40].Jensen等[40]首次通过实验验证了沉积物中的甲基汞主要是由微生物甲基化产生的,主要的汞甲基化微生物包括硫酸盐还原菌(Sulphate reduction bacteria, SRB)、铁还原菌(Iron-reducing bacteria, FeRB)和产甲烷菌(Methanogenus)等.在过去很长一段时间内,对汞甲基化的遗传机制并不清楚.直到2013年美国橡树岭国家实验室研究人员报道了hgcA和hgcB两个基因簇,可以直接调控微生物汞甲基化,从而揭示了汞的微生物甲基化遗传机制[41].其中hgcA编码类咕啉蛋白,hgcB编码铁还原蛋白,这两种蛋白分别在甲基运输以及电子提供中起着重要的作用[41-42].研究表明,在所有已被证实的汞甲基化细菌中都存在这两个基因簇,hgcAB基因的发现成为可靠的甲基化微生物的分子标记物,可以预测汞甲基化过程的存在[41-43].目前使用这一分子探针已识别到30个门类的汞甲基化细菌,Gilmour等[43-44]还在许多新环境鉴定出了汞甲基化细菌以及除硫酸盐还原菌等常见菌种之外的许多细菌.随着对微生物甲基化分子机制理解的深入,可以根据环境中hgcAB基因的含量进行甲基汞风险预测,已有证据表明土壤中hgcA丰度与甲基汞浓度呈显著正相关[45]. ...
Kinetics of methylmercury production revisited
0
2018
Mercury methylation by bacteria
1
1986
... 汞是一种全球性污染物,由于元素汞具有较高蒸汽压,在大气中的停留时间长(0.5~2年),是唯一能以气态形式进行远距离传输的重金属元素[1].人类活动排放的汞污染物可以通过大气环流迁移到地球偏远地区,并通过干、湿沉降进入陆地和水生生态系统,最终富集于冰川、植被、水体和土壤等地表环境之中[2-3].当今人为活动使大气中的总汞浓度已超出自然水平的4.5倍[4],大气沉降到地表环境之中的无机汞可以通过生物和非生物甲基化作用转化为毒性更大的甲基汞[5-7].甲基汞具有很强的神经毒性,它在生态系统食物链中具有显著的生物放大和生物积累特性[1,8].2013年,联合国环境规划署通过了旨在减排汞污染物的国际公约—《关于汞的水俣公约》,以遏制人为汞污染向环境排放输入,协议的主要目标之一是最大程度上降低与甲基汞接触相关的健康风险. ...
The chemical cycle and bioaccumulation of mercury
2
1998
... 汞是一种全球性污染物,由于元素汞具有较高蒸汽压,在大气中的停留时间长(0.5~2年),是唯一能以气态形式进行远距离传输的重金属元素[1].人类活动排放的汞污染物可以通过大气环流迁移到地球偏远地区,并通过干、湿沉降进入陆地和水生生态系统,最终富集于冰川、植被、水体和土壤等地表环境之中[2-3].当今人为活动使大气中的总汞浓度已超出自然水平的4.5倍[4],大气沉降到地表环境之中的无机汞可以通过生物和非生物甲基化作用转化为毒性更大的甲基汞[5-7].甲基汞具有很强的神经毒性,它在生态系统食物链中具有显著的生物放大和生物积累特性[1,8].2013年,联合国环境规划署通过了旨在减排汞污染物的国际公约—《关于汞的水俣公约》,以遏制人为汞污染向环境排放输入,协议的主要目标之一是最大程度上降低与甲基汞接触相关的健康风险. ...
... 不仅如此,气候变化会刺激冻土、融雪和湿地环境微生物汞甲基化,同时冻土融化的增强、降水的改变以及融雪导致地表水流量的增加,促使甲基汞向下游生态系统输送[22-25].冰川、冰盖和海冰的减少也会对冰冻圈汞生物地球化学循环产生重大影响[26-27].以上事实都表明,冰川、冻土、积雪和海冰等冰冻圈介质融化将成为陆地和海洋生态系统汞的一个重要来源,越来越多的证据亦指出冰冻圈消融向下游生态系统输送的汞总量巨大,这无疑会加剧甲基汞暴露风险[24-25,28-31].甲基汞可以在生物体内积累,进而对高营养级生物甚至人类产生潜在危害[1,8].已有研究报道北极生物体内有高水平的汞含量(如鸟类和鱼类),Franklin海湾冰下鳕鱼的总汞浓度为0.38 μg·g-1,其中80%为甲基汞[32].有研究报道北极地区居民每周从海洋食品摄入汞约846 μg,远超过世界卫生组织限定的阈值[33]. ...
3
2014
... 冰冻圈在地球上覆盖范围超过9.6×107 km2,包括冰盖、山地冰川、多年冻土、积雪、海冰、冰架等,它们主要分布在南北极和青藏高原(“第三极”)[9-10].在人类活动深刻影响自然环境的背景下,远离人类活动区的冰冻圈亦受到可全球传输的污染物影响.人类活动释放的汞经长距离传输进入偏远地区,封存于冰冻圈,使冰川、冻土等冰冻圈要素成为汞的临时储库[11-17].Schuster等[15]估算了整个北半球冻土区的汞储量约为(1 656±962) Gg,Mu等[17]估算出青藏高原活动层储存汞约16.6 Gg.沉积于北极冰川中的汞总量约为(2 415±22) Gg,Huang等[18-19]估算出中国西部冰川区每年冰尘中汞总量为34.3 kg.然而,随着全球气候持续变暖,冰冻圈要素快速退缩变化,这将导致储存于冰冻圈中的污染物发生“二次释放”,将对生态环境和人体健康产生不利影响[4,9].据估计,格陵兰冰盖西南部融水径流中汞通量约为3~42.4 kg·a-1[20].Schaefer等[21]模拟了RCP8.5排放情景下冻土退化导致Yukon流域汞输出将增加一倍(约为11 Mg·a-1).Dastoord等[18]估算出亚北极地区(60° N以北)河流向北冰洋输出溶解汞和颗粒汞总量约为(41±4) Mg·a-1,主要来源于季节性积雪和多年冻土区. ...
... ,9].据估计,格陵兰冰盖西南部融水径流中汞通量约为3~42.4 kg·a-1[20].Schaefer等[21]模拟了RCP8.5排放情景下冻土退化导致Yukon流域汞输出将增加一倍(约为11 Mg·a-1).Dastoord等[18]估算出亚北极地区(60° N以北)河流向北冰洋输出溶解汞和颗粒汞总量约为(41±4) Mg·a-1,主要来源于季节性积雪和多年冻土区. ...
... 冰川对气候变化响应非常敏感,由于气候变暖,冰川正在以前所未有的速度退缩[9].冰川消融不仅给下游生态系统带来水资源,而且还可能释放出在冰川中存储了几个世纪的汞[20].在消融初期,雪冰中的汞以脉冲式释放到冰川径流,近90%的汞会被释放进入融水[76].Sun等[77-78]估算出喜马拉雅地区融水径流汞输出通量为337 kg·a-1,青藏高原冬克玛底流域中汞输出总量约为747.4 g·a-1,亚北极Yukon流域汞输出总量约为4 400 kg·a-1[79-80].Zolkos等[81]对六条北极主要河流进行全年采样调查,估算出总汞输出通量为20 Mg·a-1,Dastoor等[18]估算了北极冰川补给流域总汞年产率为(0.03~3.9 g·km2·a-1).尽管青藏高原河流输出总量较低,但山地冰川(如扎当冰川等)的汞输出通量要高于大部分河流,表明青藏高原山地冰川补给流域具有更高的汞释放效率[30-31,78].在未来全球升温1.5 ℃情景下,到21世纪末青藏高原及周边地区冰川储量将减少2 169~3 217 km3,预估计在未来约11~16吨汞污染物将从冰川释放到下游地区(冰川中汞平均浓度为5.6 ng·L-1)[82-84].富含湿地的冰川流域通常有着更高的总汞和甲基汞含量,生物群落中的汞含量亦较高[85].阿拉斯加东南部冰河湾下游集水区,存在湿地的溪流溶解汞的比例大得多(0.1~3.4 ng·L-1),而裸露的冰川径流溶解汞含量仅为0.1~0.9 ng·L-1[85].湿地可以提供丰富的溶解性有机物,同时这种厌氧环境还有利于使汞甲基化的硫酸盐还原菌生存,可以增强微生物的活性和汞甲基化[85-87].汞污染物进入湿地之后被汞甲基化微生物转化为毒性更强的甲基汞再次进入径流,对下游地区人类健康和生态系统存在潜在威胁[88-91].因此,冰川流域生物群落中的甲基汞浓度与流域湿地覆盖率之间存在密切关系,冰川流域湿地可能成为冰川径流中甲基汞净来源,并将其输送到下游生态系统[85,88,92-95]. ...
2
2017
... 冰冻圈在地球上覆盖范围超过9.6×107 km2,包括冰盖、山地冰川、多年冻土、积雪、海冰、冰架等,它们主要分布在南北极和青藏高原(“第三极”)[9-10].在人类活动深刻影响自然环境的背景下,远离人类活动区的冰冻圈亦受到可全球传输的污染物影响.人类活动释放的汞经长距离传输进入偏远地区,封存于冰冻圈,使冰川、冻土等冰冻圈要素成为汞的临时储库[11-17].Schuster等[15]估算了整个北半球冻土区的汞储量约为(1 656±962) Gg,Mu等[17]估算出青藏高原活动层储存汞约16.6 Gg.沉积于北极冰川中的汞总量约为(2 415±22) Gg,Huang等[18-19]估算出中国西部冰川区每年冰尘中汞总量为34.3 kg.然而,随着全球气候持续变暖,冰冻圈要素快速退缩变化,这将导致储存于冰冻圈中的污染物发生“二次释放”,将对生态环境和人体健康产生不利影响[4,9].据估计,格陵兰冰盖西南部融水径流中汞通量约为3~42.4 kg·a-1[20].Schaefer等[21]模拟了RCP8.5排放情景下冻土退化导致Yukon流域汞输出将增加一倍(约为11 Mg·a-1).Dastoord等[18]估算出亚北极地区(60° N以北)河流向北冰洋输出溶解汞和颗粒汞总量约为(41±4) Mg·a-1,主要来源于季节性积雪和多年冻土区. ...
... 冰川作为冰冻圈重要组成因素,全球山地冰川(不包含格陵兰冰盖和南极冰盖)覆盖面积约为17×104 km2,共约11×104条冰川,主要分布在亚北极和青藏高原等地[10].早期Jitaru等[70]报道了欧洲阿尔卑斯山雪冰中溶解性甲基汞的含量(0.3~4.0 ng·L-1).近年来科研人员对我国青藏高原冰川也开展了研究工作,Huang等[19]对中国西部冰川上冰尘甲基汞含量进行测试分析,测得甲基汞平均含量为(1.0±0.4) ng·g-1,并初步估算出每年冰尘中的甲基汞总量约为(0.7±0.3) kg.微生物群落在汞的生物地球化学转化过程中非常活跃,冰川微生物活动可能发生汞微生物甲基化[71-74].大气汞沉降到青藏高原冰川之后在冰尘中积累,由于冰尘洞会发生季节性融化,这里便成为各种微生物群落的栖息地[75].前人研究指出我国西部冰川冰尘甲基汞与总汞的比值较高,说明冰尘表面可能是冰川汞甲基化的潜在位点[19].Zhang等[56]对青藏高原冰川开展了汞微生物甲基化初步研究,结合老虎沟12号冰川末端6个不同类型样品的汞形态和宏基因组学分析,发现在含大量碎屑和粉尘的尘埃层中汞浓度和汞甲基化基因(hgcA)丰度最高.不过该研究中汞甲基化细菌的未分类序列占比很高,微生物群落组成仍不明晰,尚未能全面地阐明汞甲基化的微生物学机制,需要进一步深入研究. ...
2
2017
... 冰冻圈在地球上覆盖范围超过9.6×107 km2,包括冰盖、山地冰川、多年冻土、积雪、海冰、冰架等,它们主要分布在南北极和青藏高原(“第三极”)[9-10].在人类活动深刻影响自然环境的背景下,远离人类活动区的冰冻圈亦受到可全球传输的污染物影响.人类活动释放的汞经长距离传输进入偏远地区,封存于冰冻圈,使冰川、冻土等冰冻圈要素成为汞的临时储库[11-17].Schuster等[15]估算了整个北半球冻土区的汞储量约为(1 656±962) Gg,Mu等[17]估算出青藏高原活动层储存汞约16.6 Gg.沉积于北极冰川中的汞总量约为(2 415±22) Gg,Huang等[18-19]估算出中国西部冰川区每年冰尘中汞总量为34.3 kg.然而,随着全球气候持续变暖,冰冻圈要素快速退缩变化,这将导致储存于冰冻圈中的污染物发生“二次释放”,将对生态环境和人体健康产生不利影响[4,9].据估计,格陵兰冰盖西南部融水径流中汞通量约为3~42.4 kg·a-1[20].Schaefer等[21]模拟了RCP8.5排放情景下冻土退化导致Yukon流域汞输出将增加一倍(约为11 Mg·a-1).Dastoord等[18]估算出亚北极地区(60° N以北)河流向北冰洋输出溶解汞和颗粒汞总量约为(41±4) Mg·a-1,主要来源于季节性积雪和多年冻土区. ...
... 冰川作为冰冻圈重要组成因素,全球山地冰川(不包含格陵兰冰盖和南极冰盖)覆盖面积约为17×104 km2,共约11×104条冰川,主要分布在亚北极和青藏高原等地[10].早期Jitaru等[70]报道了欧洲阿尔卑斯山雪冰中溶解性甲基汞的含量(0.3~4.0 ng·L-1).近年来科研人员对我国青藏高原冰川也开展了研究工作,Huang等[19]对中国西部冰川上冰尘甲基汞含量进行测试分析,测得甲基汞平均含量为(1.0±0.4) ng·g-1,并初步估算出每年冰尘中的甲基汞总量约为(0.7±0.3) kg.微生物群落在汞的生物地球化学转化过程中非常活跃,冰川微生物活动可能发生汞微生物甲基化[71-74].大气汞沉降到青藏高原冰川之后在冰尘中积累,由于冰尘洞会发生季节性融化,这里便成为各种微生物群落的栖息地[75].前人研究指出我国西部冰川冰尘甲基汞与总汞的比值较高,说明冰尘表面可能是冰川汞甲基化的潜在位点[19].Zhang等[56]对青藏高原冰川开展了汞微生物甲基化初步研究,结合老虎沟12号冰川末端6个不同类型样品的汞形态和宏基因组学分析,发现在含大量碎屑和粉尘的尘埃层中汞浓度和汞甲基化基因(hgcA)丰度最高.不过该研究中汞甲基化细菌的未分类序列占比很高,微生物群落组成仍不明晰,尚未能全面地阐明汞甲基化的微生物学机制,需要进一步深入研究. ...
summary for policy-makers
1
2021
... 冰冻圈在地球上覆盖范围超过9.6×107 km2,包括冰盖、山地冰川、多年冻土、积雪、海冰、冰架等,它们主要分布在南北极和青藏高原(“第三极”)[9-10].在人类活动深刻影响自然环境的背景下,远离人类活动区的冰冻圈亦受到可全球传输的污染物影响.人类活动释放的汞经长距离传输进入偏远地区,封存于冰冻圈,使冰川、冻土等冰冻圈要素成为汞的临时储库[11-17].Schuster等[15]估算了整个北半球冻土区的汞储量约为(1 656±962) Gg,Mu等[17]估算出青藏高原活动层储存汞约16.6 Gg.沉积于北极冰川中的汞总量约为(2 415±22) Gg,Huang等[18-19]估算出中国西部冰川区每年冰尘中汞总量为34.3 kg.然而,随着全球气候持续变暖,冰冻圈要素快速退缩变化,这将导致储存于冰冻圈中的污染物发生“二次释放”,将对生态环境和人体健康产生不利影响[4,9].据估计,格陵兰冰盖西南部融水径流中汞通量约为3~42.4 kg·a-1[20].Schaefer等[21]模拟了RCP8.5排放情景下冻土退化导致Yukon流域汞输出将增加一倍(约为11 Mg·a-1).Dastoord等[18]估算出亚北极地区(60° N以北)河流向北冰洋输出溶解汞和颗粒汞总量约为(41±4) Mg·a-1,主要来源于季节性积雪和多年冻土区. ...
Atmospheric transport of mercury to the Tibetan Plateau
0
2007
Mercury in a Greenland ice sheet: evidence of recent input by man
0
1971
Atmospheric mercury depositional chronology reconstructed from lake sediments and ice core in the Himalayas and Tibetan Plateau
0
2016
Permafrost Stores a Globally Significant Amount of Mercury
1
2018
... 冰冻圈在地球上覆盖范围超过9.6×107 km2,包括冰盖、山地冰川、多年冻土、积雪、海冰、冰架等,它们主要分布在南北极和青藏高原(“第三极”)[9-10].在人类活动深刻影响自然环境的背景下,远离人类活动区的冰冻圈亦受到可全球传输的污染物影响.人类活动释放的汞经长距离传输进入偏远地区,封存于冰冻圈,使冰川、冻土等冰冻圈要素成为汞的临时储库[11-17].Schuster等[15]估算了整个北半球冻土区的汞储量约为(1 656±962) Gg,Mu等[17]估算出青藏高原活动层储存汞约16.6 Gg.沉积于北极冰川中的汞总量约为(2 415±22) Gg,Huang等[18-19]估算出中国西部冰川区每年冰尘中汞总量为34.3 kg.然而,随着全球气候持续变暖,冰冻圈要素快速退缩变化,这将导致储存于冰冻圈中的污染物发生“二次释放”,将对生态环境和人体健康产生不利影响[4,9].据估计,格陵兰冰盖西南部融水径流中汞通量约为3~42.4 kg·a-1[20].Schaefer等[21]模拟了RCP8.5排放情景下冻土退化导致Yukon流域汞输出将增加一倍(约为11 Mg·a-1).Dastoord等[18]估算出亚北极地区(60° N以北)河流向北冰洋输出溶解汞和颗粒汞总量约为(41±4) Mg·a-1,主要来源于季节性积雪和多年冻土区. ...
Mercury in active-layer tundra soils of Alaska: concentrations, pools, origins, and spatial distribution
2
2018
... 多年冻土蕴藏着大量的汞存储量,有研究估算了北极苔原土壤储存汞约184 Gg;青藏高原活动层储存汞约16.6 Gg[16-17].冻土甲基汞的分布和转化等过程更为复杂,受各种物理、化学和生物因素控制,包括温度、溶解性有机质(DOC)、碱度、pH、总汞浓度、微生物以及其他金属元素或离子(如铁、锰)等[22,46,99-101].MacMillan等[22]发现冻土环境中甲基汞与溶解有机碳等营养物质,以及甲基汞与微生物之间有很强的相关性.当温度较高(8 ℃)时,在富含有机物和矿物质的土壤层中,甲基汞净产量比温度较低(-2 ℃)时增加了10倍以上[23].可溶性土壤有机碳类型及其生物可利用性,对促进甲基汞的初始合成特别重要[99].当冻土活动层融化时,低分子量溶解有机质增加促使形成大量生物可利用的Hg-DOM复合物,从而使无机汞的可利用性增加,更有利于冻土汞甲基化[23-24,100]. ...
... Concentrations of MeHg in the cryospheric environment
Table 2| 地点 | 样品类型 | 甲基汞 | 总汞 | 文献来源 |
|---|
| Devon Island, Canada | 多年冻土 | 0.3~3.1 ng·g-1 | — | [58] |
| Barrow, Alaska | 热融湖塘 | 0.3~2.3 ng·L-1 | 25.3~141.2 ng·L-1 | [23] |
| Tibetan Plateau, China | 多年冻土 | — | 5.6~39.3 ng·g-1 | [17] |
| Fennoscandia | 多年冻土 | 0.004~28 ng·g-1 | 1.1~210 ng·g-1 | [24] |
| Tussock tundra, Alaska | 多年冻土 | — | 56~226 ng·g-1 | [16] |
| Laohugou No.12, Tibetan Plateau, China | 雪冰 | 0.005~0.5 ng·L-1 | 24.4~172.1 ng·L-1 | [56] |
| Tibetan Plateau, China | 冰尘 | 0.4~1.7 ng·g-1 | 17.9~114.5 ng·g-1 | [19] |
| French Alps | 表层雪 | 0.002~0.3 ng·L-1 | 1.8~169.5 ng·L-1 | [57] |
| Spitsbergen Island, Norway | 表层雪 | 0.004~0.5 ng·L-1 | 1~90 ng·L-1 | [123] |
| Resolute Bay, High Arctic, Canada | 表层雪 | <0.015~0.2 ng·L-1 | 2.6~150 ng·L-1 | [125] |
| Hudson Bay, Canada | 表层雪 | 0.04~0.14 ng·L-1 | 1.6~5.1 ng·L-1 | [120] |
| Alberta, Canada | 表层雪 | 0.02~0.3 ng·L-1 | 0.8~9.0 ng·L-1 | [137] |
| Antarctic | 积雪 | <0.02~0.3 ng·L-1 | 1.0~30.6 ng·L-1 | [59] |
| McMurdo/Ross Island, Antarctic | 积雪 | — | 40~430 ng·L-1 | [114] |
| Casey Station, Antarctic | 海冰 | <0.009~0.09 ng·L-1 | 0.2~5.7 ng·L-1 | [129] |
| Antarctic | 海冰 | <0.02~0.2 ng·L-1 | 1.4~179 ng·L-1 | [59] |
| Beaufort Sea and McClure Strait, Arctic | 海冰 | <0.02~0.5 ng·L-1 | 0.1~12.2 ng·L-1 | [28] |
| Southern Ocean | 海水 | <0.02~0.1 ng·L-1 | 0. 2~23 ng·L-1 | [59] |
| Arctic Ocean | 海水 | 0.006~0.07 ng·L-1 | 0.09~1.4 ng·L-1 | [135] |
| Resolute Bay, High Arctic, Canada | 海水 | 0.06~0.1 ng·L-1 | 0.1~0.2 ng·L-1 | [125] |
注:表格中海水均为海冰覆盖水域. ...
Permafrost degradation enhances the risk of mercury release on Qinghai-Tibetan Plateau
6
2020
... 冰冻圈在地球上覆盖范围超过9.6×107 km2,包括冰盖、山地冰川、多年冻土、积雪、海冰、冰架等,它们主要分布在南北极和青藏高原(“第三极”)[9-10].在人类活动深刻影响自然环境的背景下,远离人类活动区的冰冻圈亦受到可全球传输的污染物影响.人类活动释放的汞经长距离传输进入偏远地区,封存于冰冻圈,使冰川、冻土等冰冻圈要素成为汞的临时储库[11-17].Schuster等[15]估算了整个北半球冻土区的汞储量约为(1 656±962) Gg,Mu等[17]估算出青藏高原活动层储存汞约16.6 Gg.沉积于北极冰川中的汞总量约为(2 415±22) Gg,Huang等[18-19]估算出中国西部冰川区每年冰尘中汞总量为34.3 kg.然而,随着全球气候持续变暖,冰冻圈要素快速退缩变化,这将导致储存于冰冻圈中的污染物发生“二次释放”,将对生态环境和人体健康产生不利影响[4,9].据估计,格陵兰冰盖西南部融水径流中汞通量约为3~42.4 kg·a-1[20].Schaefer等[21]模拟了RCP8.5排放情景下冻土退化导致Yukon流域汞输出将增加一倍(约为11 Mg·a-1).Dastoord等[18]估算出亚北极地区(60° N以北)河流向北冰洋输出溶解汞和颗粒汞总量约为(41±4) Mg·a-1,主要来源于季节性积雪和多年冻土区. ...
... [17]估算出青藏高原活动层储存汞约16.6 Gg.沉积于北极冰川中的汞总量约为(2 415±22) Gg,Huang等[18-19]估算出中国西部冰川区每年冰尘中汞总量为34.3 kg.然而,随着全球气候持续变暖,冰冻圈要素快速退缩变化,这将导致储存于冰冻圈中的污染物发生“二次释放”,将对生态环境和人体健康产生不利影响[4,9].据估计,格陵兰冰盖西南部融水径流中汞通量约为3~42.4 kg·a-1[20].Schaefer等[21]模拟了RCP8.5排放情景下冻土退化导致Yukon流域汞输出将增加一倍(约为11 Mg·a-1).Dastoord等[18]估算出亚北极地区(60° N以北)河流向北冰洋输出溶解汞和颗粒汞总量约为(41±4) Mg·a-1,主要来源于季节性积雪和多年冻土区. ...
... 冰冻圈中甲基汞含量极低(pg级),因此要特别注意在采样和分析过程中潜在的汞污染问题,需严格遵循“clean hands-dirty hands”的操作规范[50].甲基汞浓度测定通常采用EPA推荐的方法(Method 1631)进行测试,即“蒸馏-乙基化-吹洗和捕获-冷原子荧光光谱法(CVAFS)”相结合的方法[51].样品在进行预处理后,还需要对汞形态进行分离,最常用的汞形态分离技术为气相色谱(GC)或高效液相色谱技术(HPLC).国内对甲基汞检测方法的研究起步较晚,早期一直采用气相色谱法测定甲基汞,但方法灵敏度较低[52].近些年来,气相色谱与冷原子荧光光谱法联用技术被广泛应用于各种环境样品甲基汞的测定[17,19].之后,电感耦合等离子体质谱法(ICP-MS)、同位素稀释-电感耦合等离子体质谱法(ID-CV-ICP-MS)以及同位素稀释固相微萃取-气相色谱-电感耦合等离子体质谱法(SPME-GC-ICP-MS)等手段也陆续被应用到甲基汞的检测分析[53-55].这些方法灵敏度高,检出限低,能很好地满足冰冻圈样品中低水平甲基汞的测定,目前常用的甲基汞测试方法列于表1. ...
... Methods for the determination of MeHg in various environmental samples
Table 1| 样品类型 | 测定方法 | 检出限 | 文献来源 |
|---|
| 雪冰 | GC-CVAFS | 0.02 ng·L-1 | [56] |
| 雪冰 | GC-AFS | 0.002 ng·L-1 | [57] |
| 冻土 | HPIC-CVAFS | 0.018 ng·g-1 | [58] |
| 冻土 | CVAFS | 0.006 ng·g-1 | [17] |
| 海冰 | GC-ICP-MS | 0.02 ng·L-1 | [59] |
| 冰尘 | GC-CVAFS | 0.02 ng·g-1 | [19] |
| 融水 | GC-AFS | 0.02 ng·L-1 | [60] |
| 降水 | GC-CVAFS | 0.002 ng·L-1 | [61] |
| 鱼类 | GC-CVAFS | 0.12 ng·g-1 | [62] |
| 水稻 | GC-CVAFS | 0.003 ng·g-1 | [63] |
2.2 汞甲基化微生物早期对汞甲基化微生物的鉴定依赖于实验室培养,测试从厌氧沉积物中培养的分离物在添加无机汞后产生甲基汞的能力[64-65].然而,不同汞甲基化细菌的甲基化能力差异悬殊,如果忽视它是一个物种或菌种的特异性功能,使用这种方法预估环境中汞的甲基化潜力是欠准确的[43].目前主要通过16S rRNA基因的分析,评估微生物群落结构与环境中甲基汞和生物地球化学特性的关系[45,66-67].高通量测序作为一种新兴的免培养分子生物学技术,检测快速、准确、信息全面丰富,随着高通量测序技术的不断升级换代,测序通量、读长和准确度也有了很大提升,为认识各种环境中汞甲基化微生物多样性、群落结构组成及其生态功能提供了有利手段[68].Illumina公司的Solexa和Hiseq是目前全球使用量最大的第二代测序机器.现代分子生物学技术将从环境样品中提取的核酸(DNA或RNA)为分析对象,以进行目标片段的聚合酶链式反应(PCR)扩增,得到的PCR产物通过测序技术来鉴定微生物组成及其多样性.Mpbio公司的FastDNA Spin Kit for Soil试剂盒目前已广泛应用于DNA的提取,它是获得雪冰微生物高分子量、高纯度的基因组DNA非常重要的前提. ...
... 多年冻土蕴藏着大量的汞存储量,有研究估算了北极苔原土壤储存汞约184 Gg;青藏高原活动层储存汞约16.6 Gg[16-17].冻土甲基汞的分布和转化等过程更为复杂,受各种物理、化学和生物因素控制,包括温度、溶解性有机质(DOC)、碱度、pH、总汞浓度、微生物以及其他金属元素或离子(如铁、锰)等[22,46,99-101].MacMillan等[22]发现冻土环境中甲基汞与溶解有机碳等营养物质,以及甲基汞与微生物之间有很强的相关性.当温度较高(8 ℃)时,在富含有机物和矿物质的土壤层中,甲基汞净产量比温度较低(-2 ℃)时增加了10倍以上[23].可溶性土壤有机碳类型及其生物可利用性,对促进甲基汞的初始合成特别重要[99].当冻土活动层融化时,低分子量溶解有机质增加促使形成大量生物可利用的Hg-DOM复合物,从而使无机汞的可利用性增加,更有利于冻土汞甲基化[23-24,100]. ...
... Concentrations of MeHg in the cryospheric environment
Table 2| 地点 | 样品类型 | 甲基汞 | 总汞 | 文献来源 |
|---|
| Devon Island, Canada | 多年冻土 | 0.3~3.1 ng·g-1 | — | [58] |
| Barrow, Alaska | 热融湖塘 | 0.3~2.3 ng·L-1 | 25.3~141.2 ng·L-1 | [23] |
| Tibetan Plateau, China | 多年冻土 | — | 5.6~39.3 ng·g-1 | [17] |
| Fennoscandia | 多年冻土 | 0.004~28 ng·g-1 | 1.1~210 ng·g-1 | [24] |
| Tussock tundra, Alaska | 多年冻土 | — | 56~226 ng·g-1 | [16] |
| Laohugou No.12, Tibetan Plateau, China | 雪冰 | 0.005~0.5 ng·L-1 | 24.4~172.1 ng·L-1 | [56] |
| Tibetan Plateau, China | 冰尘 | 0.4~1.7 ng·g-1 | 17.9~114.5 ng·g-1 | [19] |
| French Alps | 表层雪 | 0.002~0.3 ng·L-1 | 1.8~169.5 ng·L-1 | [57] |
| Spitsbergen Island, Norway | 表层雪 | 0.004~0.5 ng·L-1 | 1~90 ng·L-1 | [123] |
| Resolute Bay, High Arctic, Canada | 表层雪 | <0.015~0.2 ng·L-1 | 2.6~150 ng·L-1 | [125] |
| Hudson Bay, Canada | 表层雪 | 0.04~0.14 ng·L-1 | 1.6~5.1 ng·L-1 | [120] |
| Alberta, Canada | 表层雪 | 0.02~0.3 ng·L-1 | 0.8~9.0 ng·L-1 | [137] |
| Antarctic | 积雪 | <0.02~0.3 ng·L-1 | 1.0~30.6 ng·L-1 | [59] |
| McMurdo/Ross Island, Antarctic | 积雪 | — | 40~430 ng·L-1 | [114] |
| Casey Station, Antarctic | 海冰 | <0.009~0.09 ng·L-1 | 0.2~5.7 ng·L-1 | [129] |
| Antarctic | 海冰 | <0.02~0.2 ng·L-1 | 1.4~179 ng·L-1 | [59] |
| Beaufort Sea and McClure Strait, Arctic | 海冰 | <0.02~0.5 ng·L-1 | 0.1~12.2 ng·L-1 | [28] |
| Southern Ocean | 海水 | <0.02~0.1 ng·L-1 | 0. 2~23 ng·L-1 | [59] |
| Arctic Ocean | 海水 | 0.006~0.07 ng·L-1 | 0.09~1.4 ng·L-1 | [135] |
| Resolute Bay, High Arctic, Canada | 海水 | 0.06~0.1 ng·L-1 | 0.1~0.2 ng·L-1 | [125] |
注:表格中海水均为海冰覆盖水域. ...
Arctic mercury cycling
3
2022
... 冰冻圈在地球上覆盖范围超过9.6×107 km2,包括冰盖、山地冰川、多年冻土、积雪、海冰、冰架等,它们主要分布在南北极和青藏高原(“第三极”)[9-10].在人类活动深刻影响自然环境的背景下,远离人类活动区的冰冻圈亦受到可全球传输的污染物影响.人类活动释放的汞经长距离传输进入偏远地区,封存于冰冻圈,使冰川、冻土等冰冻圈要素成为汞的临时储库[11-17].Schuster等[15]估算了整个北半球冻土区的汞储量约为(1 656±962) Gg,Mu等[17]估算出青藏高原活动层储存汞约16.6 Gg.沉积于北极冰川中的汞总量约为(2 415±22) Gg,Huang等[18-19]估算出中国西部冰川区每年冰尘中汞总量为34.3 kg.然而,随着全球气候持续变暖,冰冻圈要素快速退缩变化,这将导致储存于冰冻圈中的污染物发生“二次释放”,将对生态环境和人体健康产生不利影响[4,9].据估计,格陵兰冰盖西南部融水径流中汞通量约为3~42.4 kg·a-1[20].Schaefer等[21]模拟了RCP8.5排放情景下冻土退化导致Yukon流域汞输出将增加一倍(约为11 Mg·a-1).Dastoord等[18]估算出亚北极地区(60° N以北)河流向北冰洋输出溶解汞和颗粒汞总量约为(41±4) Mg·a-1,主要来源于季节性积雪和多年冻土区. ...
... [18]估算出亚北极地区(60° N以北)河流向北冰洋输出溶解汞和颗粒汞总量约为(41±4) Mg·a-1,主要来源于季节性积雪和多年冻土区. ...
... 冰川对气候变化响应非常敏感,由于气候变暖,冰川正在以前所未有的速度退缩[9].冰川消融不仅给下游生态系统带来水资源,而且还可能释放出在冰川中存储了几个世纪的汞[20].在消融初期,雪冰中的汞以脉冲式释放到冰川径流,近90%的汞会被释放进入融水[76].Sun等[77-78]估算出喜马拉雅地区融水径流汞输出通量为337 kg·a-1,青藏高原冬克玛底流域中汞输出总量约为747.4 g·a-1,亚北极Yukon流域汞输出总量约为4 400 kg·a-1[79-80].Zolkos等[81]对六条北极主要河流进行全年采样调查,估算出总汞输出通量为20 Mg·a-1,Dastoor等[18]估算了北极冰川补给流域总汞年产率为(0.03~3.9 g·km2·a-1).尽管青藏高原河流输出总量较低,但山地冰川(如扎当冰川等)的汞输出通量要高于大部分河流,表明青藏高原山地冰川补给流域具有更高的汞释放效率[30-31,78].在未来全球升温1.5 ℃情景下,到21世纪末青藏高原及周边地区冰川储量将减少2 169~3 217 km3,预估计在未来约11~16吨汞污染物将从冰川释放到下游地区(冰川中汞平均浓度为5.6 ng·L-1)[82-84].富含湿地的冰川流域通常有着更高的总汞和甲基汞含量,生物群落中的汞含量亦较高[85].阿拉斯加东南部冰河湾下游集水区,存在湿地的溪流溶解汞的比例大得多(0.1~3.4 ng·L-1),而裸露的冰川径流溶解汞含量仅为0.1~0.9 ng·L-1[85].湿地可以提供丰富的溶解性有机物,同时这种厌氧环境还有利于使汞甲基化的硫酸盐还原菌生存,可以增强微生物的活性和汞甲基化[85-87].汞污染物进入湿地之后被汞甲基化微生物转化为毒性更强的甲基汞再次进入径流,对下游地区人类健康和生态系统存在潜在威胁[88-91].因此,冰川流域生物群落中的甲基汞浓度与流域湿地覆盖率之间存在密切关系,冰川流域湿地可能成为冰川径流中甲基汞净来源,并将其输送到下游生态系统[85,88,92-95]. ...
Large subglacial source of mercury from the southwestern margin of the Greenland Ice Sheet
6
2021
... 冰冻圈在地球上覆盖范围超过9.6×107 km2,包括冰盖、山地冰川、多年冻土、积雪、海冰、冰架等,它们主要分布在南北极和青藏高原(“第三极”)[9-10].在人类活动深刻影响自然环境的背景下,远离人类活动区的冰冻圈亦受到可全球传输的污染物影响.人类活动释放的汞经长距离传输进入偏远地区,封存于冰冻圈,使冰川、冻土等冰冻圈要素成为汞的临时储库[11-17].Schuster等[15]估算了整个北半球冻土区的汞储量约为(1 656±962) Gg,Mu等[17]估算出青藏高原活动层储存汞约16.6 Gg.沉积于北极冰川中的汞总量约为(2 415±22) Gg,Huang等[18-19]估算出中国西部冰川区每年冰尘中汞总量为34.3 kg.然而,随着全球气候持续变暖,冰冻圈要素快速退缩变化,这将导致储存于冰冻圈中的污染物发生“二次释放”,将对生态环境和人体健康产生不利影响[4,9].据估计,格陵兰冰盖西南部融水径流中汞通量约为3~42.4 kg·a-1[20].Schaefer等[21]模拟了RCP8.5排放情景下冻土退化导致Yukon流域汞输出将增加一倍(约为11 Mg·a-1).Dastoord等[18]估算出亚北极地区(60° N以北)河流向北冰洋输出溶解汞和颗粒汞总量约为(41±4) Mg·a-1,主要来源于季节性积雪和多年冻土区. ...
... 冰冻圈中甲基汞含量极低(pg级),因此要特别注意在采样和分析过程中潜在的汞污染问题,需严格遵循“clean hands-dirty hands”的操作规范[50].甲基汞浓度测定通常采用EPA推荐的方法(Method 1631)进行测试,即“蒸馏-乙基化-吹洗和捕获-冷原子荧光光谱法(CVAFS)”相结合的方法[51].样品在进行预处理后,还需要对汞形态进行分离,最常用的汞形态分离技术为气相色谱(GC)或高效液相色谱技术(HPLC).国内对甲基汞检测方法的研究起步较晚,早期一直采用气相色谱法测定甲基汞,但方法灵敏度较低[52].近些年来,气相色谱与冷原子荧光光谱法联用技术被广泛应用于各种环境样品甲基汞的测定[17,19].之后,电感耦合等离子体质谱法(ICP-MS)、同位素稀释-电感耦合等离子体质谱法(ID-CV-ICP-MS)以及同位素稀释固相微萃取-气相色谱-电感耦合等离子体质谱法(SPME-GC-ICP-MS)等手段也陆续被应用到甲基汞的检测分析[53-55].这些方法灵敏度高,检出限低,能很好地满足冰冻圈样品中低水平甲基汞的测定,目前常用的甲基汞测试方法列于表1. ...
... Methods for the determination of MeHg in various environmental samples
Table 1| 样品类型 | 测定方法 | 检出限 | 文献来源 |
|---|
| 雪冰 | GC-CVAFS | 0.02 ng·L-1 | [56] |
| 雪冰 | GC-AFS | 0.002 ng·L-1 | [57] |
| 冻土 | HPIC-CVAFS | 0.018 ng·g-1 | [58] |
| 冻土 | CVAFS | 0.006 ng·g-1 | [17] |
| 海冰 | GC-ICP-MS | 0.02 ng·L-1 | [59] |
| 冰尘 | GC-CVAFS | 0.02 ng·g-1 | [19] |
| 融水 | GC-AFS | 0.02 ng·L-1 | [60] |
| 降水 | GC-CVAFS | 0.002 ng·L-1 | [61] |
| 鱼类 | GC-CVAFS | 0.12 ng·g-1 | [62] |
| 水稻 | GC-CVAFS | 0.003 ng·g-1 | [63] |
2.2 汞甲基化微生物早期对汞甲基化微生物的鉴定依赖于实验室培养,测试从厌氧沉积物中培养的分离物在添加无机汞后产生甲基汞的能力[64-65].然而,不同汞甲基化细菌的甲基化能力差异悬殊,如果忽视它是一个物种或菌种的特异性功能,使用这种方法预估环境中汞的甲基化潜力是欠准确的[43].目前主要通过16S rRNA基因的分析,评估微生物群落结构与环境中甲基汞和生物地球化学特性的关系[45,66-67].高通量测序作为一种新兴的免培养分子生物学技术,检测快速、准确、信息全面丰富,随着高通量测序技术的不断升级换代,测序通量、读长和准确度也有了很大提升,为认识各种环境中汞甲基化微生物多样性、群落结构组成及其生态功能提供了有利手段[68].Illumina公司的Solexa和Hiseq是目前全球使用量最大的第二代测序机器.现代分子生物学技术将从环境样品中提取的核酸(DNA或RNA)为分析对象,以进行目标片段的聚合酶链式反应(PCR)扩增,得到的PCR产物通过测序技术来鉴定微生物组成及其多样性.Mpbio公司的FastDNA Spin Kit for Soil试剂盒目前已广泛应用于DNA的提取,它是获得雪冰微生物高分子量、高纯度的基因组DNA非常重要的前提. ...
... 冰川作为冰冻圈重要组成因素,全球山地冰川(不包含格陵兰冰盖和南极冰盖)覆盖面积约为17×104 km2,共约11×104条冰川,主要分布在亚北极和青藏高原等地[10].早期Jitaru等[70]报道了欧洲阿尔卑斯山雪冰中溶解性甲基汞的含量(0.3~4.0 ng·L-1).近年来科研人员对我国青藏高原冰川也开展了研究工作,Huang等[19]对中国西部冰川上冰尘甲基汞含量进行测试分析,测得甲基汞平均含量为(1.0±0.4) ng·g-1,并初步估算出每年冰尘中的甲基汞总量约为(0.7±0.3) kg.微生物群落在汞的生物地球化学转化过程中非常活跃,冰川微生物活动可能发生汞微生物甲基化[71-74].大气汞沉降到青藏高原冰川之后在冰尘中积累,由于冰尘洞会发生季节性融化,这里便成为各种微生物群落的栖息地[75].前人研究指出我国西部冰川冰尘甲基汞与总汞的比值较高,说明冰尘表面可能是冰川汞甲基化的潜在位点[19].Zhang等[56]对青藏高原冰川开展了汞微生物甲基化初步研究,结合老虎沟12号冰川末端6个不同类型样品的汞形态和宏基因组学分析,发现在含大量碎屑和粉尘的尘埃层中汞浓度和汞甲基化基因(hgcA)丰度最高.不过该研究中汞甲基化细菌的未分类序列占比很高,微生物群落组成仍不明晰,尚未能全面地阐明汞甲基化的微生物学机制,需要进一步深入研究. ...
... [19].Zhang等[56]对青藏高原冰川开展了汞微生物甲基化初步研究,结合老虎沟12号冰川末端6个不同类型样品的汞形态和宏基因组学分析,发现在含大量碎屑和粉尘的尘埃层中汞浓度和汞甲基化基因(hgcA)丰度最高.不过该研究中汞甲基化细菌的未分类序列占比很高,微生物群落组成仍不明晰,尚未能全面地阐明汞甲基化的微生物学机制,需要进一步深入研究. ...
... Concentrations of MeHg in the cryospheric environment
Table 2| 地点 | 样品类型 | 甲基汞 | 总汞 | 文献来源 |
|---|
| Devon Island, Canada | 多年冻土 | 0.3~3.1 ng·g-1 | — | [58] |
| Barrow, Alaska | 热融湖塘 | 0.3~2.3 ng·L-1 | 25.3~141.2 ng·L-1 | [23] |
| Tibetan Plateau, China | 多年冻土 | — | 5.6~39.3 ng·g-1 | [17] |
| Fennoscandia | 多年冻土 | 0.004~28 ng·g-1 | 1.1~210 ng·g-1 | [24] |
| Tussock tundra, Alaska | 多年冻土 | — | 56~226 ng·g-1 | [16] |
| Laohugou No.12, Tibetan Plateau, China | 雪冰 | 0.005~0.5 ng·L-1 | 24.4~172.1 ng·L-1 | [56] |
| Tibetan Plateau, China | 冰尘 | 0.4~1.7 ng·g-1 | 17.9~114.5 ng·g-1 | [19] |
| French Alps | 表层雪 | 0.002~0.3 ng·L-1 | 1.8~169.5 ng·L-1 | [57] |
| Spitsbergen Island, Norway | 表层雪 | 0.004~0.5 ng·L-1 | 1~90 ng·L-1 | [123] |
| Resolute Bay, High Arctic, Canada | 表层雪 | <0.015~0.2 ng·L-1 | 2.6~150 ng·L-1 | [125] |
| Hudson Bay, Canada | 表层雪 | 0.04~0.14 ng·L-1 | 1.6~5.1 ng·L-1 | [120] |
| Alberta, Canada | 表层雪 | 0.02~0.3 ng·L-1 | 0.8~9.0 ng·L-1 | [137] |
| Antarctic | 积雪 | <0.02~0.3 ng·L-1 | 1.0~30.6 ng·L-1 | [59] |
| McMurdo/Ross Island, Antarctic | 积雪 | — | 40~430 ng·L-1 | [114] |
| Casey Station, Antarctic | 海冰 | <0.009~0.09 ng·L-1 | 0.2~5.7 ng·L-1 | [129] |
| Antarctic | 海冰 | <0.02~0.2 ng·L-1 | 1.4~179 ng·L-1 | [59] |
| Beaufort Sea and McClure Strait, Arctic | 海冰 | <0.02~0.5 ng·L-1 | 0.1~12.2 ng·L-1 | [28] |
| Southern Ocean | 海水 | <0.02~0.1 ng·L-1 | 0. 2~23 ng·L-1 | [59] |
| Arctic Ocean | 海水 | 0.006~0.07 ng·L-1 | 0.09~1.4 ng·L-1 | [135] |
| Resolute Bay, High Arctic, Canada | 海水 | 0.06~0.1 ng·L-1 | 0.1~0.2 ng·L-1 | [125] |
注:表格中海水均为海冰覆盖水域. ...
Accumulation of atmospheric mercury in glacier cryoconite over Western China
3
2019
... 冰冻圈在地球上覆盖范围超过9.6×107 km2,包括冰盖、山地冰川、多年冻土、积雪、海冰、冰架等,它们主要分布在南北极和青藏高原(“第三极”)[9-10].在人类活动深刻影响自然环境的背景下,远离人类活动区的冰冻圈亦受到可全球传输的污染物影响.人类活动释放的汞经长距离传输进入偏远地区,封存于冰冻圈,使冰川、冻土等冰冻圈要素成为汞的临时储库[11-17].Schuster等[15]估算了整个北半球冻土区的汞储量约为(1 656±962) Gg,Mu等[17]估算出青藏高原活动层储存汞约16.6 Gg.沉积于北极冰川中的汞总量约为(2 415±22) Gg,Huang等[18-19]估算出中国西部冰川区每年冰尘中汞总量为34.3 kg.然而,随着全球气候持续变暖,冰冻圈要素快速退缩变化,这将导致储存于冰冻圈中的污染物发生“二次释放”,将对生态环境和人体健康产生不利影响[4,9].据估计,格陵兰冰盖西南部融水径流中汞通量约为3~42.4 kg·a-1[20].Schaefer等[21]模拟了RCP8.5排放情景下冻土退化导致Yukon流域汞输出将增加一倍(约为11 Mg·a-1).Dastoord等[18]估算出亚北极地区(60° N以北)河流向北冰洋输出溶解汞和颗粒汞总量约为(41±4) Mg·a-1,主要来源于季节性积雪和多年冻土区. ...
... 冰川对气候变化响应非常敏感,由于气候变暖,冰川正在以前所未有的速度退缩[9].冰川消融不仅给下游生态系统带来水资源,而且还可能释放出在冰川中存储了几个世纪的汞[20].在消融初期,雪冰中的汞以脉冲式释放到冰川径流,近90%的汞会被释放进入融水[76].Sun等[77-78]估算出喜马拉雅地区融水径流汞输出通量为337 kg·a-1,青藏高原冬克玛底流域中汞输出总量约为747.4 g·a-1,亚北极Yukon流域汞输出总量约为4 400 kg·a-1[79-80].Zolkos等[81]对六条北极主要河流进行全年采样调查,估算出总汞输出通量为20 Mg·a-1,Dastoor等[18]估算了北极冰川补给流域总汞年产率为(0.03~3.9 g·km2·a-1).尽管青藏高原河流输出总量较低,但山地冰川(如扎当冰川等)的汞输出通量要高于大部分河流,表明青藏高原山地冰川补给流域具有更高的汞释放效率[30-31,78].在未来全球升温1.5 ℃情景下,到21世纪末青藏高原及周边地区冰川储量将减少2 169~3 217 km3,预估计在未来约11~16吨汞污染物将从冰川释放到下游地区(冰川中汞平均浓度为5.6 ng·L-1)[82-84].富含湿地的冰川流域通常有着更高的总汞和甲基汞含量,生物群落中的汞含量亦较高[85].阿拉斯加东南部冰河湾下游集水区,存在湿地的溪流溶解汞的比例大得多(0.1~3.4 ng·L-1),而裸露的冰川径流溶解汞含量仅为0.1~0.9 ng·L-1[85].湿地可以提供丰富的溶解性有机物,同时这种厌氧环境还有利于使汞甲基化的硫酸盐还原菌生存,可以增强微生物的活性和汞甲基化[85-87].汞污染物进入湿地之后被汞甲基化微生物转化为毒性更强的甲基汞再次进入径流,对下游地区人类健康和生态系统存在潜在威胁[88-91].因此,冰川流域生物群落中的甲基汞浓度与流域湿地覆盖率之间存在密切关系,冰川流域湿地可能成为冰川径流中甲基汞净来源,并将其输送到下游生态系统[85,88,92-95]. ...
... 大量研究表明偏远冰冻圈下游地区水生生态系统甲基汞的富集效应是非常显著的[85,96-97].格陵兰冰盖下游湾峡水生生态系统中,融水径流汞浓度(44~348 ng·L-1)比北极其他河流(0.3~2.8 ng·L-1)高出1~2个数量级,甲基汞浓度(1.0~3.5 ng·L-1)甚至高于大多数淡水,与湿地水平相当[20].我国学者亦研究发现青藏高原河流和湖泊中鱼类甲基汞含量(24.9~1 196 ng·g-1)为中国野生鱼类汞浓度的最高水平[96].此外,前人在冰川补给湖观测到甲基汞的峰值出现在化学跃层,甲基汞占总汞的比例高达57%,同时16S rRNA数据显示检测出硫酸盐还原菌,因此受冰川融水补给的冰川湖化学跃层存在着较强的汞甲基化潜力[97-98]. ...
Potential impacts of mercury released from thawing permafrost
1
2020
... 冰冻圈在地球上覆盖范围超过9.6×107 km2,包括冰盖、山地冰川、多年冻土、积雪、海冰、冰架等,它们主要分布在南北极和青藏高原(“第三极”)[9-10].在人类活动深刻影响自然环境的背景下,远离人类活动区的冰冻圈亦受到可全球传输的污染物影响.人类活动释放的汞经长距离传输进入偏远地区,封存于冰冻圈,使冰川、冻土等冰冻圈要素成为汞的临时储库[11-17].Schuster等[15]估算了整个北半球冻土区的汞储量约为(1 656±962) Gg,Mu等[17]估算出青藏高原活动层储存汞约16.6 Gg.沉积于北极冰川中的汞总量约为(2 415±22) Gg,Huang等[18-19]估算出中国西部冰川区每年冰尘中汞总量为34.3 kg.然而,随着全球气候持续变暖,冰冻圈要素快速退缩变化,这将导致储存于冰冻圈中的污染物发生“二次释放”,将对生态环境和人体健康产生不利影响[4,9].据估计,格陵兰冰盖西南部融水径流中汞通量约为3~42.4 kg·a-1[20].Schaefer等[21]模拟了RCP8.5排放情景下冻土退化导致Yukon流域汞输出将增加一倍(约为11 Mg·a-1).Dastoord等[18]估算出亚北极地区(60° N以北)河流向北冰洋输出溶解汞和颗粒汞总量约为(41±4) Mg·a-1,主要来源于季节性积雪和多年冻土区. ...
High methylmercury in Arctic and subarctic ponds is related to nutrient levels in the warming eastern Canadian Arctic
5
2015
... 不仅如此,气候变化会刺激冻土、融雪和湿地环境微生物汞甲基化,同时冻土融化的增强、降水的改变以及融雪导致地表水流量的增加,促使甲基汞向下游生态系统输送[22-25].冰川、冰盖和海冰的减少也会对冰冻圈汞生物地球化学循环产生重大影响[26-27].以上事实都表明,冰川、冻土、积雪和海冰等冰冻圈介质融化将成为陆地和海洋生态系统汞的一个重要来源,越来越多的证据亦指出冰冻圈消融向下游生态系统输送的汞总量巨大,这无疑会加剧甲基汞暴露风险[24-25,28-31].甲基汞可以在生物体内积累,进而对高营养级生物甚至人类产生潜在危害[1,8].已有研究报道北极生物体内有高水平的汞含量(如鸟类和鱼类),Franklin海湾冰下鳕鱼的总汞浓度为0.38 μg·g-1,其中80%为甲基汞[32].有研究报道北极地区居民每周从海洋食品摄入汞约846 μg,远超过世界卫生组织限定的阈值[33]. ...
... 多年冻土蕴藏着大量的汞存储量,有研究估算了北极苔原土壤储存汞约184 Gg;青藏高原活动层储存汞约16.6 Gg[16-17].冻土甲基汞的分布和转化等过程更为复杂,受各种物理、化学和生物因素控制,包括温度、溶解性有机质(DOC)、碱度、pH、总汞浓度、微生物以及其他金属元素或离子(如铁、锰)等[22,46,99-101].MacMillan等[22]发现冻土环境中甲基汞与溶解有机碳等营养物质,以及甲基汞与微生物之间有很强的相关性.当温度较高(8 ℃)时,在富含有机物和矿物质的土壤层中,甲基汞净产量比温度较低(-2 ℃)时增加了10倍以上[23].可溶性土壤有机碳类型及其生物可利用性,对促进甲基汞的初始合成特别重要[99].当冻土活动层融化时,低分子量溶解有机质增加促使形成大量生物可利用的Hg-DOM复合物,从而使无机汞的可利用性增加,更有利于冻土汞甲基化[23-24,100]. ...
... [22]发现冻土环境中甲基汞与溶解有机碳等营养物质,以及甲基汞与微生物之间有很强的相关性.当温度较高(8 ℃)时,在富含有机物和矿物质的土壤层中,甲基汞净产量比温度较低(-2 ℃)时增加了10倍以上[23].可溶性土壤有机碳类型及其生物可利用性,对促进甲基汞的初始合成特别重要[99].当冻土活动层融化时,低分子量溶解有机质增加促使形成大量生物可利用的Hg-DOM复合物,从而使无机汞的可利用性增加,更有利于冻土汞甲基化[23-24,100]. ...
... 由于气候变暖,多年冻土快速融化、塌陷,形成热融喀斯特地貌,其中最显著的是形成热融湖塘,此过程更有利于微生物将无机汞转化为甲基汞[22-24,102-104].Tarbier等[24]在亚北极多年冻土区测量了178个土壤样品,甲基汞的浓度范围为0.01~28 ng·g-1,与其他地区土壤甲基汞相比要高得多(例如阿拉斯加北部<0.1~3.4 ng·g-1),同时发现热融池水和热岩孔隙水中甲基汞含量亦较高(0.01~3.1 ng·L-1)[22,24,58,100].在融化阶段,塌陷的冻土沼泽地比完整冻土带活动层的净甲基化程度还要高一个数量级(甲基汞占总汞的比例高出13倍),表明当热融湖塘形成时,冻土区的汞甲基化率将会显著提高[24].这主要是由于冻融塌陷使得冻土泥炭表面被淹没在水位以下,从而形成缺氧环境,这种条件非常有利于具有汞甲基化能力的厌氧微生物生存(包括硫酸盐还原菌、铁还原菌及产甲烷菌等)[58,91,100,104-105]. ...
... [22,24,58,100].在融化阶段,塌陷的冻土沼泽地比完整冻土带活动层的净甲基化程度还要高一个数量级(甲基汞占总汞的比例高出13倍),表明当热融湖塘形成时,冻土区的汞甲基化率将会显著提高[24].这主要是由于冻融塌陷使得冻土泥炭表面被淹没在水位以下,从而形成缺氧环境,这种条件非常有利于具有汞甲基化能力的厌氧微生物生存(包括硫酸盐还原菌、铁还原菌及产甲烷菌等)[58,91,100,104-105]. ...
Warming increases methylmercury production in an Arctic soil
3
2016
... 多年冻土蕴藏着大量的汞存储量,有研究估算了北极苔原土壤储存汞约184 Gg;青藏高原活动层储存汞约16.6 Gg[16-17].冻土甲基汞的分布和转化等过程更为复杂,受各种物理、化学和生物因素控制,包括温度、溶解性有机质(DOC)、碱度、pH、总汞浓度、微生物以及其他金属元素或离子(如铁、锰)等[22,46,99-101].MacMillan等[22]发现冻土环境中甲基汞与溶解有机碳等营养物质,以及甲基汞与微生物之间有很强的相关性.当温度较高(8 ℃)时,在富含有机物和矿物质的土壤层中,甲基汞净产量比温度较低(-2 ℃)时增加了10倍以上[23].可溶性土壤有机碳类型及其生物可利用性,对促进甲基汞的初始合成特别重要[99].当冻土活动层融化时,低分子量溶解有机质增加促使形成大量生物可利用的Hg-DOM复合物,从而使无机汞的可利用性增加,更有利于冻土汞甲基化[23-24,100]. ...
... [23-24,100]. ...
... Concentrations of MeHg in the cryospheric environment
Table 2| 地点 | 样品类型 | 甲基汞 | 总汞 | 文献来源 |
|---|
| Devon Island, Canada | 多年冻土 | 0.3~3.1 ng·g-1 | — | [58] |
| Barrow, Alaska | 热融湖塘 | 0.3~2.3 ng·L-1 | 25.3~141.2 ng·L-1 | [23] |
| Tibetan Plateau, China | 多年冻土 | — | 5.6~39.3 ng·g-1 | [17] |
| Fennoscandia | 多年冻土 | 0.004~28 ng·g-1 | 1.1~210 ng·g-1 | [24] |
| Tussock tundra, Alaska | 多年冻土 | — | 56~226 ng·g-1 | [16] |
| Laohugou No.12, Tibetan Plateau, China | 雪冰 | 0.005~0.5 ng·L-1 | 24.4~172.1 ng·L-1 | [56] |
| Tibetan Plateau, China | 冰尘 | 0.4~1.7 ng·g-1 | 17.9~114.5 ng·g-1 | [19] |
| French Alps | 表层雪 | 0.002~0.3 ng·L-1 | 1.8~169.5 ng·L-1 | [57] |
| Spitsbergen Island, Norway | 表层雪 | 0.004~0.5 ng·L-1 | 1~90 ng·L-1 | [123] |
| Resolute Bay, High Arctic, Canada | 表层雪 | <0.015~0.2 ng·L-1 | 2.6~150 ng·L-1 | [125] |
| Hudson Bay, Canada | 表层雪 | 0.04~0.14 ng·L-1 | 1.6~5.1 ng·L-1 | [120] |
| Alberta, Canada | 表层雪 | 0.02~0.3 ng·L-1 | 0.8~9.0 ng·L-1 | [137] |
| Antarctic | 积雪 | <0.02~0.3 ng·L-1 | 1.0~30.6 ng·L-1 | [59] |
| McMurdo/Ross Island, Antarctic | 积雪 | — | 40~430 ng·L-1 | [114] |
| Casey Station, Antarctic | 海冰 | <0.009~0.09 ng·L-1 | 0.2~5.7 ng·L-1 | [129] |
| Antarctic | 海冰 | <0.02~0.2 ng·L-1 | 1.4~179 ng·L-1 | [59] |
| Beaufort Sea and McClure Strait, Arctic | 海冰 | <0.02~0.5 ng·L-1 | 0.1~12.2 ng·L-1 | [28] |
| Southern Ocean | 海水 | <0.02~0.1 ng·L-1 | 0. 2~23 ng·L-1 | [59] |
| Arctic Ocean | 海水 | 0.006~0.07 ng·L-1 | 0.09~1.4 ng·L-1 | [135] |
| Resolute Bay, High Arctic, Canada | 海水 | 0.06~0.1 ng·L-1 | 0.1~0.2 ng·L-1 | [125] |
注:表格中海水均为海冰覆盖水域. ...
Permafrost thaw increases methylmercury formation in subarctic Fennoscandia
7
2021
... 不仅如此,气候变化会刺激冻土、融雪和湿地环境微生物汞甲基化,同时冻土融化的增强、降水的改变以及融雪导致地表水流量的增加,促使甲基汞向下游生态系统输送[22-25].冰川、冰盖和海冰的减少也会对冰冻圈汞生物地球化学循环产生重大影响[26-27].以上事实都表明,冰川、冻土、积雪和海冰等冰冻圈介质融化将成为陆地和海洋生态系统汞的一个重要来源,越来越多的证据亦指出冰冻圈消融向下游生态系统输送的汞总量巨大,这无疑会加剧甲基汞暴露风险[24-25,28-31].甲基汞可以在生物体内积累,进而对高营养级生物甚至人类产生潜在危害[1,8].已有研究报道北极生物体内有高水平的汞含量(如鸟类和鱼类),Franklin海湾冰下鳕鱼的总汞浓度为0.38 μg·g-1,其中80%为甲基汞[32].有研究报道北极地区居民每周从海洋食品摄入汞约846 μg,远超过世界卫生组织限定的阈值[33]. ...
... 多年冻土蕴藏着大量的汞存储量,有研究估算了北极苔原土壤储存汞约184 Gg;青藏高原活动层储存汞约16.6 Gg[16-17].冻土甲基汞的分布和转化等过程更为复杂,受各种物理、化学和生物因素控制,包括温度、溶解性有机质(DOC)、碱度、pH、总汞浓度、微生物以及其他金属元素或离子(如铁、锰)等[22,46,99-101].MacMillan等[22]发现冻土环境中甲基汞与溶解有机碳等营养物质,以及甲基汞与微生物之间有很强的相关性.当温度较高(8 ℃)时,在富含有机物和矿物质的土壤层中,甲基汞净产量比温度较低(-2 ℃)时增加了10倍以上[23].可溶性土壤有机碳类型及其生物可利用性,对促进甲基汞的初始合成特别重要[99].当冻土活动层融化时,低分子量溶解有机质增加促使形成大量生物可利用的Hg-DOM复合物,从而使无机汞的可利用性增加,更有利于冻土汞甲基化[23-24,100]. ...
... 由于气候变暖,多年冻土快速融化、塌陷,形成热融喀斯特地貌,其中最显著的是形成热融湖塘,此过程更有利于微生物将无机汞转化为甲基汞[22-24,102-104].Tarbier等[24]在亚北极多年冻土区测量了178个土壤样品,甲基汞的浓度范围为0.01~28 ng·g-1,与其他地区土壤甲基汞相比要高得多(例如阿拉斯加北部<0.1~3.4 ng·g-1),同时发现热融池水和热岩孔隙水中甲基汞含量亦较高(0.01~3.1 ng·L-1)[22,24,58,100].在融化阶段,塌陷的冻土沼泽地比完整冻土带活动层的净甲基化程度还要高一个数量级(甲基汞占总汞的比例高出13倍),表明当热融湖塘形成时,冻土区的汞甲基化率将会显著提高[24].这主要是由于冻融塌陷使得冻土泥炭表面被淹没在水位以下,从而形成缺氧环境,这种条件非常有利于具有汞甲基化能力的厌氧微生物生存(包括硫酸盐还原菌、铁还原菌及产甲烷菌等)[58,91,100,104-105]. ...
... [24]在亚北极多年冻土区测量了178个土壤样品,甲基汞的浓度范围为0.01~28 ng·g-1,与其他地区土壤甲基汞相比要高得多(例如阿拉斯加北部<0.1~3.4 ng·g-1),同时发现热融池水和热岩孔隙水中甲基汞含量亦较高(0.01~3.1 ng·L-1)[22,24,58,100].在融化阶段,塌陷的冻土沼泽地比完整冻土带活动层的净甲基化程度还要高一个数量级(甲基汞占总汞的比例高出13倍),表明当热融湖塘形成时,冻土区的汞甲基化率将会显著提高[24].这主要是由于冻融塌陷使得冻土泥炭表面被淹没在水位以下,从而形成缺氧环境,这种条件非常有利于具有汞甲基化能力的厌氧微生物生存(包括硫酸盐还原菌、铁还原菌及产甲烷菌等)[58,91,100,104-105]. ...
... ,24,58,100].在融化阶段,塌陷的冻土沼泽地比完整冻土带活动层的净甲基化程度还要高一个数量级(甲基汞占总汞的比例高出13倍),表明当热融湖塘形成时,冻土区的汞甲基化率将会显著提高[24].这主要是由于冻融塌陷使得冻土泥炭表面被淹没在水位以下,从而形成缺氧环境,这种条件非常有利于具有汞甲基化能力的厌氧微生物生存(包括硫酸盐还原菌、铁还原菌及产甲烷菌等)[58,91,100,104-105]. ...
... [24].这主要是由于冻融塌陷使得冻土泥炭表面被淹没在水位以下,从而形成缺氧环境,这种条件非常有利于具有汞甲基化能力的厌氧微生物生存(包括硫酸盐还原菌、铁还原菌及产甲烷菌等)[58,91,100,104-105]. ...
... Concentrations of MeHg in the cryospheric environment
Table 2| 地点 | 样品类型 | 甲基汞 | 总汞 | 文献来源 |
|---|
| Devon Island, Canada | 多年冻土 | 0.3~3.1 ng·g-1 | — | [58] |
| Barrow, Alaska | 热融湖塘 | 0.3~2.3 ng·L-1 | 25.3~141.2 ng·L-1 | [23] |
| Tibetan Plateau, China | 多年冻土 | — | 5.6~39.3 ng·g-1 | [17] |
| Fennoscandia | 多年冻土 | 0.004~28 ng·g-1 | 1.1~210 ng·g-1 | [24] |
| Tussock tundra, Alaska | 多年冻土 | — | 56~226 ng·g-1 | [16] |
| Laohugou No.12, Tibetan Plateau, China | 雪冰 | 0.005~0.5 ng·L-1 | 24.4~172.1 ng·L-1 | [56] |
| Tibetan Plateau, China | 冰尘 | 0.4~1.7 ng·g-1 | 17.9~114.5 ng·g-1 | [19] |
| French Alps | 表层雪 | 0.002~0.3 ng·L-1 | 1.8~169.5 ng·L-1 | [57] |
| Spitsbergen Island, Norway | 表层雪 | 0.004~0.5 ng·L-1 | 1~90 ng·L-1 | [123] |
| Resolute Bay, High Arctic, Canada | 表层雪 | <0.015~0.2 ng·L-1 | 2.6~150 ng·L-1 | [125] |
| Hudson Bay, Canada | 表层雪 | 0.04~0.14 ng·L-1 | 1.6~5.1 ng·L-1 | [120] |
| Alberta, Canada | 表层雪 | 0.02~0.3 ng·L-1 | 0.8~9.0 ng·L-1 | [137] |
| Antarctic | 积雪 | <0.02~0.3 ng·L-1 | 1.0~30.6 ng·L-1 | [59] |
| McMurdo/Ross Island, Antarctic | 积雪 | — | 40~430 ng·L-1 | [114] |
| Casey Station, Antarctic | 海冰 | <0.009~0.09 ng·L-1 | 0.2~5.7 ng·L-1 | [129] |
| Antarctic | 海冰 | <0.02~0.2 ng·L-1 | 1.4~179 ng·L-1 | [59] |
| Beaufort Sea and McClure Strait, Arctic | 海冰 | <0.02~0.5 ng·L-1 | 0.1~12.2 ng·L-1 | [28] |
| Southern Ocean | 海水 | <0.02~0.1 ng·L-1 | 0. 2~23 ng·L-1 | [59] |
| Arctic Ocean | 海水 | 0.006~0.07 ng·L-1 | 0.09~1.4 ng·L-1 | [135] |
| Resolute Bay, High Arctic, Canada | 海水 | 0.06~0.1 ng·L-1 | 0.1~0.2 ng·L-1 | [125] |
注:表格中海水均为海冰覆盖水域. ...
Methylmercury transport and fate shows strong seasonal and spatial variability along a high arctic freshwater hydrologic continuum
3
2021
... 不仅如此,气候变化会刺激冻土、融雪和湿地环境微生物汞甲基化,同时冻土融化的增强、降水的改变以及融雪导致地表水流量的增加,促使甲基汞向下游生态系统输送[22-25].冰川、冰盖和海冰的减少也会对冰冻圈汞生物地球化学循环产生重大影响[26-27].以上事实都表明,冰川、冻土、积雪和海冰等冰冻圈介质融化将成为陆地和海洋生态系统汞的一个重要来源,越来越多的证据亦指出冰冻圈消融向下游生态系统输送的汞总量巨大,这无疑会加剧甲基汞暴露风险[24-25,28-31].甲基汞可以在生物体内积累,进而对高营养级生物甚至人类产生潜在危害[1,8].已有研究报道北极生物体内有高水平的汞含量(如鸟类和鱼类),Franklin海湾冰下鳕鱼的总汞浓度为0.38 μg·g-1,其中80%为甲基汞[32].有研究报道北极地区居民每周从海洋食品摄入汞约846 μg,远超过世界卫生组织限定的阈值[33]. ...
... -25,28-31].甲基汞可以在生物体内积累,进而对高营养级生物甚至人类产生潜在危害[1,8].已有研究报道北极生物体内有高水平的汞含量(如鸟类和鱼类),Franklin海湾冰下鳕鱼的总汞浓度为0.38 μg·g-1,其中80%为甲基汞[32].有研究报道北极地区居民每周从海洋食品摄入汞约846 μg,远超过世界卫生组织限定的阈值[33]. ...
... 积雪中甲基汞的来源在过去被归因于大气沉降,然而随着研究的不断深入,新的证据表明在低纬度苔原的积雪中可能存在着较强的汞甲基化活动[108,111,123-127].在融雪期间,雪冰融水中甲基汞占总汞的比例增大(从2.7%增加到7.6%)[126].导致这一现象的原因一方面由于甲基汞从无机/有机物中浸出,在雪冰融化时趋于集中;另一方面是由于雪冰内部可能发生甲基化作用,微生物活动和反应可能在积雪中产生了甲基汞[108,120,123-127].极地积雪的pH值通常为酸性,有机物以及微生物等存在于雪层中,因此在消融的积雪中存在微生物汞甲基化的可能[120].随着气候变暖,温度升高会促使积雪中微生物种群数量增长,从而导致微生物影响汞甲基化的强度大幅增加[25].Loseto等[60]估算出北极地区积雪中甲基汞产量约为1.5 mg·km-2·d-1,并指出北极康沃利斯岛陆地积雪融化是地表径流中最重要的甲基汞来源. ...
Mercury, food webs, and marine mammals: implications of diet and climate change for human health
3
2005
... 不仅如此,气候变化会刺激冻土、融雪和湿地环境微生物汞甲基化,同时冻土融化的增强、降水的改变以及融雪导致地表水流量的增加,促使甲基汞向下游生态系统输送[22-25].冰川、冰盖和海冰的减少也会对冰冻圈汞生物地球化学循环产生重大影响[26-27].以上事实都表明,冰川、冻土、积雪和海冰等冰冻圈介质融化将成为陆地和海洋生态系统汞的一个重要来源,越来越多的证据亦指出冰冻圈消融向下游生态系统输送的汞总量巨大,这无疑会加剧甲基汞暴露风险[24-25,28-31].甲基汞可以在生物体内积累,进而对高营养级生物甚至人类产生潜在危害[1,8].已有研究报道北极生物体内有高水平的汞含量(如鸟类和鱼类),Franklin海湾冰下鳕鱼的总汞浓度为0.38 μg·g-1,其中80%为甲基汞[32].有研究报道北极地区居民每周从海洋食品摄入汞约846 μg,远超过世界卫生组织限定的阈值[33]. ...
... 最新的研究不断揭示出潜在的新型海洋汞甲基化微生物,它们具有更强的耐氧性和更广泛的生境范围,这些都是有利于海冰汞甲基化的理想条件[59].Gionfriddo等[59]通过对南极海冰汞甲基化研究表明,南极海冰中存在甲基汞的富集现象,主要是由微嗜氧细菌-硝化刺菌属(Nitrospina)主导产生的,参与调控微生物汞甲基化的基因为hgcA-like.这与之前的认知有所不同,过去认为只有厌氧条件支持显著的汞甲基化过程,且汞甲基化微生物都属于厌氧细菌.因此,海冰汞的研究不仅揭示了更多潜在的新型海洋汞甲基化微生物,而且极大地拓宽了我们对汞甲基化微生物耐氧性的认识.气候变暖背景下,海冰变薄、季节性海冰区扩大、表层海洋分层加剧和浮游生物生态动力学的变化,这些过程都会造成海洋甲基汞产量增加[26,136].从海冰融化释放出来的大量甲基汞进入海洋生态系统之后会在食物链中富集和放大,将会对南北极水生食物链中高营养级的动物构成很大威胁[26,28,59,134].通过对北极多年海冰的融化情况的估算,每年输入北冰洋的总汞和甲基汞的通量估计分别为420 kg·a-1和42 kg·a-1[28].南极海冰融化亦会给海洋分别带来超过9 600 kg·a-1总汞和18 kg·a-1甲基汞,总汞的估算值要比北冰洋高出一个数量级[59,134].为方便后续研究,将冰冻圈总汞和甲基汞浓度水平相关信息汇总于表2. ...
... [26,28,59,134].通过对北极多年海冰的融化情况的估算,每年输入北冰洋的总汞和甲基汞的通量估计分别为420 kg·a-1和42 kg·a-1[28].南极海冰融化亦会给海洋分别带来超过9 600 kg·a-1总汞和18 kg·a-1甲基汞,总汞的估算值要比北冰洋高出一个数量级[59,134].为方便后续研究,将冰冻圈总汞和甲基汞浓度水平相关信息汇总于表2. ...
Influence of the arctic sea-ice regime shift on sea-ice methylated mercury trends
1
2020
... 不仅如此,气候变化会刺激冻土、融雪和湿地环境微生物汞甲基化,同时冻土融化的增强、降水的改变以及融雪导致地表水流量的增加,促使甲基汞向下游生态系统输送[22-25].冰川、冰盖和海冰的减少也会对冰冻圈汞生物地球化学循环产生重大影响[26-27].以上事实都表明,冰川、冻土、积雪和海冰等冰冻圈介质融化将成为陆地和海洋生态系统汞的一个重要来源,越来越多的证据亦指出冰冻圈消融向下游生态系统输送的汞总量巨大,这无疑会加剧甲基汞暴露风险[24-25,28-31].甲基汞可以在生物体内积累,进而对高营养级生物甚至人类产生潜在危害[1,8].已有研究报道北极生物体内有高水平的汞含量(如鸟类和鱼类),Franklin海湾冰下鳕鱼的总汞浓度为0.38 μg·g-1,其中80%为甲基汞[32].有研究报道北极地区居民每周从海洋食品摄入汞约846 μg,远超过世界卫生组织限定的阈值[33]. ...
Total and methylated mercury in Arctic multiyear sea ice
7
2014
... 不仅如此,气候变化会刺激冻土、融雪和湿地环境微生物汞甲基化,同时冻土融化的增强、降水的改变以及融雪导致地表水流量的增加,促使甲基汞向下游生态系统输送[22-25].冰川、冰盖和海冰的减少也会对冰冻圈汞生物地球化学循环产生重大影响[26-27].以上事实都表明,冰川、冻土、积雪和海冰等冰冻圈介质融化将成为陆地和海洋生态系统汞的一个重要来源,越来越多的证据亦指出冰冻圈消融向下游生态系统输送的汞总量巨大,这无疑会加剧甲基汞暴露风险[24-25,28-31].甲基汞可以在生物体内积累,进而对高营养级生物甚至人类产生潜在危害[1,8].已有研究报道北极生物体内有高水平的汞含量(如鸟类和鱼类),Franklin海湾冰下鳕鱼的总汞浓度为0.38 μg·g-1,其中80%为甲基汞[32].有研究报道北极地区居民每周从海洋食品摄入汞约846 μg,远超过世界卫生组织限定的阈值[33]. ...
... 海冰中的甲基汞来源主要包括大气沉降和原位汞甲基化.海洋中的浮游植物生产的二甲基汞从海面释放,在大气中降解为甲基汞,最后又沉降到海冰或海水中,这是海冰甲基汞重要来源之一[121].Baya等[128]研究指出海冰覆盖和初级生产是促进海洋浮游植物生产二甲基汞的主要驱动因素.海冰甲基汞来源除了大气和海洋之外,海冰原位产甲基汞亦非常重要.大量研究已表明海冰中下部和冰水界面是已知的可能产生甲基汞的区位[28,129].因此,海冰中汞的原位甲基化使得海冰融化极有可能成为海洋生态系统新的甲基汞来源. ...
... Cossa等[129]在南极地区开展了“积雪-海冰-卤水-海水”连续系统中的汞循环研究.结果表明,尽管海冰与海水的甲基汞浓度范围大致相同,但在整个系统中分布较为复杂,海冰底部的甲基汞浓度达到峰值.Beattie等[28]在北极海冰中亦观测到了类似的分布模式,海冰中下部甲基汞与总汞比值通常非常高.相比底层水体,海冰具有更高的微生物活性、丰度和多样性[130-133].大气汞通过干湿沉降的方式积累在海冰和海水中为微生物甲基化提供了底物[134-135],并且海冰内部存在局部厌氧条件,成为硫酸盐还原菌和铁还原菌繁衍的氧化还原区,这种现象非常有利于海冰汞甲基化的发生[131].此外海冰-海水界面通常具有丰富的藻类、叶绿素和有机质[132-133].在春季和夏季,由于冰雪融化和浮游植物生长,无机汞被浮游植物吸收利用,这些过程极有可能在海冰-海水界面发生甲基化[133-134].已有研究表明,在南极和北极海冰底部都观测到较高水平的甲基汞含量,这与原位产生甲基汞密切相关,因为海冰底部也有较高的细菌丰度和有机质含量,而且厌氧环境亦有利于汞甲基化细菌的生长繁殖[28,129]. ...
... [28,129]. ...
... 最新的研究不断揭示出潜在的新型海洋汞甲基化微生物,它们具有更强的耐氧性和更广泛的生境范围,这些都是有利于海冰汞甲基化的理想条件[59].Gionfriddo等[59]通过对南极海冰汞甲基化研究表明,南极海冰中存在甲基汞的富集现象,主要是由微嗜氧细菌-硝化刺菌属(Nitrospina)主导产生的,参与调控微生物汞甲基化的基因为hgcA-like.这与之前的认知有所不同,过去认为只有厌氧条件支持显著的汞甲基化过程,且汞甲基化微生物都属于厌氧细菌.因此,海冰汞的研究不仅揭示了更多潜在的新型海洋汞甲基化微生物,而且极大地拓宽了我们对汞甲基化微生物耐氧性的认识.气候变暖背景下,海冰变薄、季节性海冰区扩大、表层海洋分层加剧和浮游生物生态动力学的变化,这些过程都会造成海洋甲基汞产量增加[26,136].从海冰融化释放出来的大量甲基汞进入海洋生态系统之后会在食物链中富集和放大,将会对南北极水生食物链中高营养级的动物构成很大威胁[26,28,59,134].通过对北极多年海冰的融化情况的估算,每年输入北冰洋的总汞和甲基汞的通量估计分别为420 kg·a-1和42 kg·a-1[28].南极海冰融化亦会给海洋分别带来超过9 600 kg·a-1总汞和18 kg·a-1甲基汞,总汞的估算值要比北冰洋高出一个数量级[59,134].为方便后续研究,将冰冻圈总汞和甲基汞浓度水平相关信息汇总于表2. ...
... -1[28].南极海冰融化亦会给海洋分别带来超过9 600 kg·a-1总汞和18 kg·a-1甲基汞,总汞的估算值要比北冰洋高出一个数量级[59,134].为方便后续研究,将冰冻圈总汞和甲基汞浓度水平相关信息汇总于表2. ...
... Concentrations of MeHg in the cryospheric environment
Table 2| 地点 | 样品类型 | 甲基汞 | 总汞 | 文献来源 |
|---|
| Devon Island, Canada | 多年冻土 | 0.3~3.1 ng·g-1 | — | [58] |
| Barrow, Alaska | 热融湖塘 | 0.3~2.3 ng·L-1 | 25.3~141.2 ng·L-1 | [23] |
| Tibetan Plateau, China | 多年冻土 | — | 5.6~39.3 ng·g-1 | [17] |
| Fennoscandia | 多年冻土 | 0.004~28 ng·g-1 | 1.1~210 ng·g-1 | [24] |
| Tussock tundra, Alaska | 多年冻土 | — | 56~226 ng·g-1 | [16] |
| Laohugou No.12, Tibetan Plateau, China | 雪冰 | 0.005~0.5 ng·L-1 | 24.4~172.1 ng·L-1 | [56] |
| Tibetan Plateau, China | 冰尘 | 0.4~1.7 ng·g-1 | 17.9~114.5 ng·g-1 | [19] |
| French Alps | 表层雪 | 0.002~0.3 ng·L-1 | 1.8~169.5 ng·L-1 | [57] |
| Spitsbergen Island, Norway | 表层雪 | 0.004~0.5 ng·L-1 | 1~90 ng·L-1 | [123] |
| Resolute Bay, High Arctic, Canada | 表层雪 | <0.015~0.2 ng·L-1 | 2.6~150 ng·L-1 | [125] |
| Hudson Bay, Canada | 表层雪 | 0.04~0.14 ng·L-1 | 1.6~5.1 ng·L-1 | [120] |
| Alberta, Canada | 表层雪 | 0.02~0.3 ng·L-1 | 0.8~9.0 ng·L-1 | [137] |
| Antarctic | 积雪 | <0.02~0.3 ng·L-1 | 1.0~30.6 ng·L-1 | [59] |
| McMurdo/Ross Island, Antarctic | 积雪 | — | 40~430 ng·L-1 | [114] |
| Casey Station, Antarctic | 海冰 | <0.009~0.09 ng·L-1 | 0.2~5.7 ng·L-1 | [129] |
| Antarctic | 海冰 | <0.02~0.2 ng·L-1 | 1.4~179 ng·L-1 | [59] |
| Beaufort Sea and McClure Strait, Arctic | 海冰 | <0.02~0.5 ng·L-1 | 0.1~12.2 ng·L-1 | [28] |
| Southern Ocean | 海水 | <0.02~0.1 ng·L-1 | 0. 2~23 ng·L-1 | [59] |
| Arctic Ocean | 海水 | 0.006~0.07 ng·L-1 | 0.09~1.4 ng·L-1 | [135] |
| Resolute Bay, High Arctic, Canada | 海水 | 0.06~0.1 ng·L-1 | 0.1~0.2 ng·L-1 | [125] |
注:表格中海水均为海冰覆盖水域. ...
Pierre K A, Zolkos S, Shakil S,
1
2018
... Lehnherr等[91,104]在加拿大高纬度地区的湿地湖塘中观测到汞甲基化率很高,来自原位甲基化产生(1.8~40 ng·m-2·d-1)的甲基汞远高于大气沉降(0.03~0.05 ng·m-2·d-1).Fahnestock等[100]发现在亚北极多年冻土消融区汞甲基化微生物有着更高的丰度和多样性.通过对全球多种环境样品的微生物基因组序列比对发现,融化的冻土中汞甲基化基因(hgcA、hgcB)丰度较高,而且这些基因主要存在于产甲烷菌和韧皮菌中[106].北极快速变暖和降水增多正在加速多年冻土的退化,并促进了历史遗留汞释放到现代生物地球化学循环中,多年冻土大幅度退化已经被认为是北极流域汞通量快速增加的主要控制因素[29].前人已在高原观测到大量汞从融化的多年冻土活动层中释放到热融湖塘和其他水体之中[107],然而青藏高原冻土甲基汞浓度的测定以及汞甲基化过程研究目前还处于空白阶段,这些过程极有可能会对高原汞循环产生直接和重大的影响. ...
The role of melting alpine glaciers in mercury export and transport: an intensive sampling campaign in the Qugaqie Basin, inland Tibetan Plateau
1
2017
... 冰川对气候变化响应非常敏感,由于气候变暖,冰川正在以前所未有的速度退缩[9].冰川消融不仅给下游生态系统带来水资源,而且还可能释放出在冰川中存储了几个世纪的汞[20].在消融初期,雪冰中的汞以脉冲式释放到冰川径流,近90%的汞会被释放进入融水[76].Sun等[77-78]估算出喜马拉雅地区融水径流汞输出通量为337 kg·a-1,青藏高原冬克玛底流域中汞输出总量约为747.4 g·a-1,亚北极Yukon流域汞输出总量约为4 400 kg·a-1[79-80].Zolkos等[81]对六条北极主要河流进行全年采样调查,估算出总汞输出通量为20 Mg·a-1,Dastoor等[18]估算了北极冰川补给流域总汞年产率为(0.03~3.9 g·km2·a-1).尽管青藏高原河流输出总量较低,但山地冰川(如扎当冰川等)的汞输出通量要高于大部分河流,表明青藏高原山地冰川补给流域具有更高的汞释放效率[30-31,78].在未来全球升温1.5 ℃情景下,到21世纪末青藏高原及周边地区冰川储量将减少2 169~3 217 km3,预估计在未来约11~16吨汞污染物将从冰川释放到下游地区(冰川中汞平均浓度为5.6 ng·L-1)[82-84].富含湿地的冰川流域通常有着更高的总汞和甲基汞含量,生物群落中的汞含量亦较高[85].阿拉斯加东南部冰河湾下游集水区,存在湿地的溪流溶解汞的比例大得多(0.1~3.4 ng·L-1),而裸露的冰川径流溶解汞含量仅为0.1~0.9 ng·L-1[85].湿地可以提供丰富的溶解性有机物,同时这种厌氧环境还有利于使汞甲基化的硫酸盐还原菌生存,可以增强微生物的活性和汞甲基化[85-87].汞污染物进入湿地之后被汞甲基化微生物转化为毒性更强的甲基汞再次进入径流,对下游地区人类健康和生态系统存在潜在威胁[88-91].因此,冰川流域生物群落中的甲基汞浓度与流域湿地覆盖率之间存在密切关系,冰川流域湿地可能成为冰川径流中甲基汞净来源,并将其输送到下游生态系统[85,88,92-95]. ...
Understanding mercury cycling in Tibetan glacierized mountain environment: recent progress and remaining gaps
2
2019
... 不仅如此,气候变化会刺激冻土、融雪和湿地环境微生物汞甲基化,同时冻土融化的增强、降水的改变以及融雪导致地表水流量的增加,促使甲基汞向下游生态系统输送[22-25].冰川、冰盖和海冰的减少也会对冰冻圈汞生物地球化学循环产生重大影响[26-27].以上事实都表明,冰川、冻土、积雪和海冰等冰冻圈介质融化将成为陆地和海洋生态系统汞的一个重要来源,越来越多的证据亦指出冰冻圈消融向下游生态系统输送的汞总量巨大,这无疑会加剧甲基汞暴露风险[24-25,28-31].甲基汞可以在生物体内积累,进而对高营养级生物甚至人类产生潜在危害[1,8].已有研究报道北极生物体内有高水平的汞含量(如鸟类和鱼类),Franklin海湾冰下鳕鱼的总汞浓度为0.38 μg·g-1,其中80%为甲基汞[32].有研究报道北极地区居民每周从海洋食品摄入汞约846 μg,远超过世界卫生组织限定的阈值[33]. ...
... 冰川对气候变化响应非常敏感,由于气候变暖,冰川正在以前所未有的速度退缩[9].冰川消融不仅给下游生态系统带来水资源,而且还可能释放出在冰川中存储了几个世纪的汞[20].在消融初期,雪冰中的汞以脉冲式释放到冰川径流,近90%的汞会被释放进入融水[76].Sun等[77-78]估算出喜马拉雅地区融水径流汞输出通量为337 kg·a-1,青藏高原冬克玛底流域中汞输出总量约为747.4 g·a-1,亚北极Yukon流域汞输出总量约为4 400 kg·a-1[79-80].Zolkos等[81]对六条北极主要河流进行全年采样调查,估算出总汞输出通量为20 Mg·a-1,Dastoor等[18]估算了北极冰川补给流域总汞年产率为(0.03~3.9 g·km2·a-1).尽管青藏高原河流输出总量较低,但山地冰川(如扎当冰川等)的汞输出通量要高于大部分河流,表明青藏高原山地冰川补给流域具有更高的汞释放效率[30-31,78].在未来全球升温1.5 ℃情景下,到21世纪末青藏高原及周边地区冰川储量将减少2 169~3 217 km3,预估计在未来约11~16吨汞污染物将从冰川释放到下游地区(冰川中汞平均浓度为5.6 ng·L-1)[82-84].富含湿地的冰川流域通常有着更高的总汞和甲基汞含量,生物群落中的汞含量亦较高[85].阿拉斯加东南部冰河湾下游集水区,存在湿地的溪流溶解汞的比例大得多(0.1~3.4 ng·L-1),而裸露的冰川径流溶解汞含量仅为0.1~0.9 ng·L-1[85].湿地可以提供丰富的溶解性有机物,同时这种厌氧环境还有利于使汞甲基化的硫酸盐还原菌生存,可以增强微生物的活性和汞甲基化[85-87].汞污染物进入湿地之后被汞甲基化微生物转化为毒性更强的甲基汞再次进入径流,对下游地区人类健康和生态系统存在潜在威胁[88-91].因此,冰川流域生物群落中的甲基汞浓度与流域湿地覆盖率之间存在密切关系,冰川流域湿地可能成为冰川径流中甲基汞净来源,并将其输送到下游生态系统[85,88,92-95]. ...
Linking mercury exposure to habitat and feeding behaviour in Beaufort Sea beluga whales
1
2008
... 不仅如此,气候变化会刺激冻土、融雪和湿地环境微生物汞甲基化,同时冻土融化的增强、降水的改变以及融雪导致地表水流量的增加,促使甲基汞向下游生态系统输送[22-25].冰川、冰盖和海冰的减少也会对冰冻圈汞生物地球化学循环产生重大影响[26-27].以上事实都表明,冰川、冻土、积雪和海冰等冰冻圈介质融化将成为陆地和海洋生态系统汞的一个重要来源,越来越多的证据亦指出冰冻圈消融向下游生态系统输送的汞总量巨大,这无疑会加剧甲基汞暴露风险[24-25,28-31].甲基汞可以在生物体内积累,进而对高营养级生物甚至人类产生潜在危害[1,8].已有研究报道北极生物体内有高水平的汞含量(如鸟类和鱼类),Franklin海湾冰下鳕鱼的总汞浓度为0.38 μg·g-1,其中80%为甲基汞[32].有研究报道北极地区居民每周从海洋食品摄入汞约846 μg,远超过世界卫生组织限定的阈值[33]. ...
Environmental health criteria 101: methylmercury
3
1990
... 不仅如此,气候变化会刺激冻土、融雪和湿地环境微生物汞甲基化,同时冻土融化的增强、降水的改变以及融雪导致地表水流量的增加,促使甲基汞向下游生态系统输送[22-25].冰川、冰盖和海冰的减少也会对冰冻圈汞生物地球化学循环产生重大影响[26-27].以上事实都表明,冰川、冻土、积雪和海冰等冰冻圈介质融化将成为陆地和海洋生态系统汞的一个重要来源,越来越多的证据亦指出冰冻圈消融向下游生态系统输送的汞总量巨大,这无疑会加剧甲基汞暴露风险[24-25,28-31].甲基汞可以在生物体内积累,进而对高营养级生物甚至人类产生潜在危害[1,8].已有研究报道北极生物体内有高水平的汞含量(如鸟类和鱼类),Franklin海湾冰下鳕鱼的总汞浓度为0.38 μg·g-1,其中80%为甲基汞[32].有研究报道北极地区居民每周从海洋食品摄入汞约846 μg,远超过世界卫生组织限定的阈值[33]. ...
... 冰冻圈甲基汞富集作用同样对生活在该区域的人类健康构成重大威胁.北极地区居住着土著居民(如因纽特人),他们依赖于捕获鱼类和海洋哺乳动物为食,通过饮食摄入是当地居民暴露甲基汞的主要途径[3,155-157].北极因纽特人原住民由于常年捕食海洋生物,人体内的甲基汞含量都远超过了毒理学阈值[104,158].由于甲基汞对成人和儿童的毒性效应有所不同,处于发育中的儿童中枢神经系统比成年人对甲基汞更为敏感,而孕妇长期处于低剂量甲基汞暴露将会对婴儿的智力发育产生严重影响[33].研究表明,北极一些地区50%以上的母亲和育龄妇女的血液中的汞含量已超过美国环境保护署建议的标准浓度(5.8 μg·L-1),且在格陵兰岛部分地区近乎90%的育龄妇女血液中的汞含量都超标,这些都表明我们迫切需要重视冰冻圈甲基汞暴露风险[33,157-158]. ...
... [33,157-158]. ...
Microbial mercury methylation in the cryosphere: progress and prospects
3
2019
... 冰冻圈具有地球上独特的极端环境特征,甲基汞在冰冻圈的积累、迁移和转化等行为是全球汞生物地球化学循环过程中非常重要的环节(图1)[34-36].然而控制冰冻圈甲基汞浓度的因素复杂多变,以及缺乏足够灵敏的超痕量形态分析方法使得甲基汞浓度难以量化,迄今全球对冰冻圈甲基汞的认识仍处于初级阶段,存在很大的不确定性.冰冻圈汞甲基化在生物和非生物方面都具有其独特性,目前对冰冻圈各要素中甲基汞浓度的测定和微生物多样性的认识仍匮乏,这对于研究探讨寒冷环境条件下微生物与汞甲基化的关系成为挑战.总而言之,我们对冰冻圈甲基汞的来源、产生/降解机制以及对控制甲基汞迁移和转化的生物地球化学过程的因素仍然知之甚少. ...
... 对hgcAB基因簇、mer基因簇等分子层面的研究极大地拓宽了我们对汞甲基化机制的认识[41,48].Gionfriddo等[69]开发出了一套全新的引物,并将汞甲基化细菌基因序列参考库扩展到了283个,从而可以更准确地获取汞甲基化微生物多样性和环境中甲基汞产生的潜力.当今采用高通量测序获取基因和微生物多样性的生物学信息,开辟了冰冻圈汞甲基化研究的新领域.冰冻圈汞甲基化微生物分析方法概括归纳如图2所示.分析冰冻圈中hgcAB基因的方法一种是基于聚合酶链式反应技术,利用hgcAB特异引物获得样品中的hgcAB基因序列和丰度;另一种是通过BLAST方法从样本宏基因组数据中筛选出hgcAB同源序列[34]. ...
... 自工业革命以来,人类活动已深刻改变了全球汞循环过程,造成大量人为汞污染物进入冰冻圈.近期研究发现,嗜冷菌微生物已经进化出了特殊机制来克服低温环境,并积极参与到冰冻圈汞甲基化过程,将冰冻圈自然环境中无机汞转化为甲基汞[34,138-139].由于生物富集和累积作用,海豹、鲸鱼、海鸟及北极熊体内的汞含量非常高,这些处于食物链高层的捕食者体内的汞几乎完全是甲基汞形式[140-152].在北极生物群落中海鸟作为顶级捕食者,寿命长、分布广泛而且对汞吸收很敏感,因此海鸟经常被用作海洋生态系统中汞富集研究的生物指示剂[145].研究发现海鸟羽毛中的汞浓度已超过5 μg·g-1(dw)的毒性阈值,而这种高水平浓度通常被认为有害健康和影响生殖[144-145,153-154].南极磷虾是污染物向上层营养级生物转移的重要环节,磷虾体内总汞浓度范围为4.04~19.4 ng·g-1,甲基汞浓度范围为0.26~2.94 ng·g-1,依赖磷虾的消费者(如幼鱼)通过食物链大量富集和积累甲基汞[146-148].已有研究报道南极海洋食物链顶端的物种(如企鹅和哺乳动物)体内甲基汞富集系数高达多个数量级,在北极地区捕食性鱼类中甲基汞含量甚至比海水高出约一百万倍[149-150]. ...
Environmental biogeochemistry of mercury in Antarctic ecosystems
0
2007
Should we not further study the impact of microbial activity on snow and polar atmospheric chemistry?
1
2019
... 冰冻圈具有地球上独特的极端环境特征,甲基汞在冰冻圈的积累、迁移和转化等行为是全球汞生物地球化学循环过程中非常重要的环节(图1)[34-36].然而控制冰冻圈甲基汞浓度的因素复杂多变,以及缺乏足够灵敏的超痕量形态分析方法使得甲基汞浓度难以量化,迄今全球对冰冻圈甲基汞的认识仍处于初级阶段,存在很大的不确定性.冰冻圈汞甲基化在生物和非生物方面都具有其独特性,目前对冰冻圈各要素中甲基汞浓度的测定和微生物多样性的认识仍匮乏,这对于研究探讨寒冷环境条件下微生物与汞甲基化的关系成为挑战.总而言之,我们对冰冻圈甲基汞的来源、产生/降解机制以及对控制甲基汞迁移和转化的生物地球化学过程的因素仍然知之甚少. ...
Review of possible paths for abiotic methylation of mercury (II) in the aquatic environment
1
1993
... 汞的甲基化包括生物和非生物两种途径[5,37-40].在自然条件下汞的非生物甲基化效率非常低,通常认为环境中的甲基汞是由微生物主导产生[38-40].Jensen等[40]首次通过实验验证了沉积物中的甲基汞主要是由微生物甲基化产生的,主要的汞甲基化微生物包括硫酸盐还原菌(Sulphate reduction bacteria, SRB)、铁还原菌(Iron-reducing bacteria, FeRB)和产甲烷菌(Methanogenus)等.在过去很长一段时间内,对汞甲基化的遗传机制并不清楚.直到2013年美国橡树岭国家实验室研究人员报道了hgcA和hgcB两个基因簇,可以直接调控微生物汞甲基化,从而揭示了汞的微生物甲基化遗传机制[41].其中hgcA编码类咕啉蛋白,hgcB编码铁还原蛋白,这两种蛋白分别在甲基运输以及电子提供中起着重要的作用[41-42].研究表明,在所有已被证实的汞甲基化细菌中都存在这两个基因簇,hgcAB基因的发现成为可靠的甲基化微生物的分子标记物,可以预测汞甲基化过程的存在[41-43].目前使用这一分子探针已识别到30个门类的汞甲基化细菌,Gilmour等[43-44]还在许多新环境鉴定出了汞甲基化细菌以及除硫酸盐还原菌等常见菌种之外的许多细菌.随着对微生物甲基化分子机制理解的深入,可以根据环境中hgcAB基因的含量进行甲基汞风险预测,已有证据表明土壤中hgcA丰度与甲基汞浓度呈显著正相关[45]. ...
Mercury methylation bacteria and methylmercury producing: a review
1
2013
... 汞的甲基化包括生物和非生物两种途径[5,37-40].在自然条件下汞的非生物甲基化效率非常低,通常认为环境中的甲基汞是由微生物主导产生[38-40].Jensen等[40]首次通过实验验证了沉积物中的甲基汞主要是由微生物甲基化产生的,主要的汞甲基化微生物包括硫酸盐还原菌(Sulphate reduction bacteria, SRB)、铁还原菌(Iron-reducing bacteria, FeRB)和产甲烷菌(Methanogenus)等.在过去很长一段时间内,对汞甲基化的遗传机制并不清楚.直到2013年美国橡树岭国家实验室研究人员报道了hgcA和hgcB两个基因簇,可以直接调控微生物汞甲基化,从而揭示了汞的微生物甲基化遗传机制[41].其中hgcA编码类咕啉蛋白,hgcB编码铁还原蛋白,这两种蛋白分别在甲基运输以及电子提供中起着重要的作用[41-42].研究表明,在所有已被证实的汞甲基化细菌中都存在这两个基因簇,hgcAB基因的发现成为可靠的甲基化微生物的分子标记物,可以预测汞甲基化过程的存在[41-43].目前使用这一分子探针已识别到30个门类的汞甲基化细菌,Gilmour等[43-44]还在许多新环境鉴定出了汞甲基化细菌以及除硫酸盐还原菌等常见菌种之外的许多细菌.随着对微生物甲基化分子机制理解的深入,可以根据环境中hgcAB基因的含量进行甲基汞风险预测,已有证据表明土壤中hgcA丰度与甲基汞浓度呈显著正相关[45]. ...
汞甲基化细菌研究进展
1
2013
... 汞的甲基化包括生物和非生物两种途径[5,37-40].在自然条件下汞的非生物甲基化效率非常低,通常认为环境中的甲基汞是由微生物主导产生[38-40].Jensen等[40]首次通过实验验证了沉积物中的甲基汞主要是由微生物甲基化产生的,主要的汞甲基化微生物包括硫酸盐还原菌(Sulphate reduction bacteria, SRB)、铁还原菌(Iron-reducing bacteria, FeRB)和产甲烷菌(Methanogenus)等.在过去很长一段时间内,对汞甲基化的遗传机制并不清楚.直到2013年美国橡树岭国家实验室研究人员报道了hgcA和hgcB两个基因簇,可以直接调控微生物汞甲基化,从而揭示了汞的微生物甲基化遗传机制[41].其中hgcA编码类咕啉蛋白,hgcB编码铁还原蛋白,这两种蛋白分别在甲基运输以及电子提供中起着重要的作用[41-42].研究表明,在所有已被证实的汞甲基化细菌中都存在这两个基因簇,hgcAB基因的发现成为可靠的甲基化微生物的分子标记物,可以预测汞甲基化过程的存在[41-43].目前使用这一分子探针已识别到30个门类的汞甲基化细菌,Gilmour等[43-44]还在许多新环境鉴定出了汞甲基化细菌以及除硫酸盐还原菌等常见菌种之外的许多细菌.随着对微生物甲基化分子机制理解的深入,可以根据环境中hgcAB基因的含量进行甲基汞风险预测,已有证据表明土壤中hgcA丰度与甲基汞浓度呈显著正相关[45]. ...
Microbial transformations of mercury: potentials, challenges, and achievements in controlling mercury toxicity in the environment
0
2005
Biological methylation of mercury in aquatic organisms
3
1969
... 汞的甲基化包括生物和非生物两种途径[5,37-40].在自然条件下汞的非生物甲基化效率非常低,通常认为环境中的甲基汞是由微生物主导产生[38-40].Jensen等[40]首次通过实验验证了沉积物中的甲基汞主要是由微生物甲基化产生的,主要的汞甲基化微生物包括硫酸盐还原菌(Sulphate reduction bacteria, SRB)、铁还原菌(Iron-reducing bacteria, FeRB)和产甲烷菌(Methanogenus)等.在过去很长一段时间内,对汞甲基化的遗传机制并不清楚.直到2013年美国橡树岭国家实验室研究人员报道了hgcA和hgcB两个基因簇,可以直接调控微生物汞甲基化,从而揭示了汞的微生物甲基化遗传机制[41].其中hgcA编码类咕啉蛋白,hgcB编码铁还原蛋白,这两种蛋白分别在甲基运输以及电子提供中起着重要的作用[41-42].研究表明,在所有已被证实的汞甲基化细菌中都存在这两个基因簇,hgcAB基因的发现成为可靠的甲基化微生物的分子标记物,可以预测汞甲基化过程的存在[41-43].目前使用这一分子探针已识别到30个门类的汞甲基化细菌,Gilmour等[43-44]还在许多新环境鉴定出了汞甲基化细菌以及除硫酸盐还原菌等常见菌种之外的许多细菌.随着对微生物甲基化分子机制理解的深入,可以根据环境中hgcAB基因的含量进行甲基汞风险预测,已有证据表明土壤中hgcA丰度与甲基汞浓度呈显著正相关[45]. ...
... -40].Jensen等[40]首次通过实验验证了沉积物中的甲基汞主要是由微生物甲基化产生的,主要的汞甲基化微生物包括硫酸盐还原菌(Sulphate reduction bacteria, SRB)、铁还原菌(Iron-reducing bacteria, FeRB)和产甲烷菌(Methanogenus)等.在过去很长一段时间内,对汞甲基化的遗传机制并不清楚.直到2013年美国橡树岭国家实验室研究人员报道了hgcA和hgcB两个基因簇,可以直接调控微生物汞甲基化,从而揭示了汞的微生物甲基化遗传机制[41].其中hgcA编码类咕啉蛋白,hgcB编码铁还原蛋白,这两种蛋白分别在甲基运输以及电子提供中起着重要的作用[41-42].研究表明,在所有已被证实的汞甲基化细菌中都存在这两个基因簇,hgcAB基因的发现成为可靠的甲基化微生物的分子标记物,可以预测汞甲基化过程的存在[41-43].目前使用这一分子探针已识别到30个门类的汞甲基化细菌,Gilmour等[43-44]还在许多新环境鉴定出了汞甲基化细菌以及除硫酸盐还原菌等常见菌种之外的许多细菌.随着对微生物甲基化分子机制理解的深入,可以根据环境中hgcAB基因的含量进行甲基汞风险预测,已有证据表明土壤中hgcA丰度与甲基汞浓度呈显著正相关[45]. ...
... [40]首次通过实验验证了沉积物中的甲基汞主要是由微生物甲基化产生的,主要的汞甲基化微生物包括硫酸盐还原菌(Sulphate reduction bacteria, SRB)、铁还原菌(Iron-reducing bacteria, FeRB)和产甲烷菌(Methanogenus)等.在过去很长一段时间内,对汞甲基化的遗传机制并不清楚.直到2013年美国橡树岭国家实验室研究人员报道了hgcA和hgcB两个基因簇,可以直接调控微生物汞甲基化,从而揭示了汞的微生物甲基化遗传机制[41].其中hgcA编码类咕啉蛋白,hgcB编码铁还原蛋白,这两种蛋白分别在甲基运输以及电子提供中起着重要的作用[41-42].研究表明,在所有已被证实的汞甲基化细菌中都存在这两个基因簇,hgcAB基因的发现成为可靠的甲基化微生物的分子标记物,可以预测汞甲基化过程的存在[41-43].目前使用这一分子探针已识别到30个门类的汞甲基化细菌,Gilmour等[43-44]还在许多新环境鉴定出了汞甲基化细菌以及除硫酸盐还原菌等常见菌种之外的许多细菌.随着对微生物甲基化分子机制理解的深入,可以根据环境中hgcAB基因的含量进行甲基汞风险预测,已有证据表明土壤中hgcA丰度与甲基汞浓度呈显著正相关[45]. ...
The genetic basis for bacterial mercury methylation
4
2013
... 汞的甲基化包括生物和非生物两种途径[5,37-40].在自然条件下汞的非生物甲基化效率非常低,通常认为环境中的甲基汞是由微生物主导产生[38-40].Jensen等[40]首次通过实验验证了沉积物中的甲基汞主要是由微生物甲基化产生的,主要的汞甲基化微生物包括硫酸盐还原菌(Sulphate reduction bacteria, SRB)、铁还原菌(Iron-reducing bacteria, FeRB)和产甲烷菌(Methanogenus)等.在过去很长一段时间内,对汞甲基化的遗传机制并不清楚.直到2013年美国橡树岭国家实验室研究人员报道了hgcA和hgcB两个基因簇,可以直接调控微生物汞甲基化,从而揭示了汞的微生物甲基化遗传机制[41].其中hgcA编码类咕啉蛋白,hgcB编码铁还原蛋白,这两种蛋白分别在甲基运输以及电子提供中起着重要的作用[41-42].研究表明,在所有已被证实的汞甲基化细菌中都存在这两个基因簇,hgcAB基因的发现成为可靠的甲基化微生物的分子标记物,可以预测汞甲基化过程的存在[41-43].目前使用这一分子探针已识别到30个门类的汞甲基化细菌,Gilmour等[43-44]还在许多新环境鉴定出了汞甲基化细菌以及除硫酸盐还原菌等常见菌种之外的许多细菌.随着对微生物甲基化分子机制理解的深入,可以根据环境中hgcAB基因的含量进行甲基汞风险预测,已有证据表明土壤中hgcA丰度与甲基汞浓度呈显著正相关[45]. ...
... [41-42].研究表明,在所有已被证实的汞甲基化细菌中都存在这两个基因簇,hgcAB基因的发现成为可靠的甲基化微生物的分子标记物,可以预测汞甲基化过程的存在[41-43].目前使用这一分子探针已识别到30个门类的汞甲基化细菌,Gilmour等[43-44]还在许多新环境鉴定出了汞甲基化细菌以及除硫酸盐还原菌等常见菌种之外的许多细菌.随着对微生物甲基化分子机制理解的深入,可以根据环境中hgcAB基因的含量进行甲基汞风险预测,已有证据表明土壤中hgcA丰度与甲基汞浓度呈显著正相关[45]. ...
... [41-43].目前使用这一分子探针已识别到30个门类的汞甲基化细菌,Gilmour等[43-44]还在许多新环境鉴定出了汞甲基化细菌以及除硫酸盐还原菌等常见菌种之外的许多细菌.随着对微生物甲基化分子机制理解的深入,可以根据环境中hgcAB基因的含量进行甲基汞风险预测,已有证据表明土壤中hgcA丰度与甲基汞浓度呈显著正相关[45]. ...
... 对hgcAB基因簇、mer基因簇等分子层面的研究极大地拓宽了我们对汞甲基化机制的认识[41,48].Gionfriddo等[69]开发出了一套全新的引物,并将汞甲基化细菌基因序列参考库扩展到了283个,从而可以更准确地获取汞甲基化微生物多样性和环境中甲基汞产生的潜力.当今采用高通量测序获取基因和微生物多样性的生物学信息,开辟了冰冻圈汞甲基化研究的新领域.冰冻圈汞甲基化微生物分析方法概括归纳如图2所示.分析冰冻圈中hgcAB基因的方法一种是基于聚合酶链式反应技术,利用hgcAB特异引物获得样品中的hgcAB基因序列和丰度;另一种是通过BLAST方法从样本宏基因组数据中筛选出hgcAB同源序列[34]. ...
Cracking the mercury methylation code
1
2013
... 汞的甲基化包括生物和非生物两种途径[5,37-40].在自然条件下汞的非生物甲基化效率非常低,通常认为环境中的甲基汞是由微生物主导产生[38-40].Jensen等[40]首次通过实验验证了沉积物中的甲基汞主要是由微生物甲基化产生的,主要的汞甲基化微生物包括硫酸盐还原菌(Sulphate reduction bacteria, SRB)、铁还原菌(Iron-reducing bacteria, FeRB)和产甲烷菌(Methanogenus)等.在过去很长一段时间内,对汞甲基化的遗传机制并不清楚.直到2013年美国橡树岭国家实验室研究人员报道了hgcA和hgcB两个基因簇,可以直接调控微生物汞甲基化,从而揭示了汞的微生物甲基化遗传机制[41].其中hgcA编码类咕啉蛋白,hgcB编码铁还原蛋白,这两种蛋白分别在甲基运输以及电子提供中起着重要的作用[41-42].研究表明,在所有已被证实的汞甲基化细菌中都存在这两个基因簇,hgcAB基因的发现成为可靠的甲基化微生物的分子标记物,可以预测汞甲基化过程的存在[41-43].目前使用这一分子探针已识别到30个门类的汞甲基化细菌,Gilmour等[43-44]还在许多新环境鉴定出了汞甲基化细菌以及除硫酸盐还原菌等常见菌种之外的许多细菌.随着对微生物甲基化分子机制理解的深入,可以根据环境中hgcAB基因的含量进行甲基汞风险预测,已有证据表明土壤中hgcA丰度与甲基汞浓度呈显著正相关[45]. ...
Mercury methylation by novel microorganisms from new environments
3
2013
... 汞的甲基化包括生物和非生物两种途径[5,37-40].在自然条件下汞的非生物甲基化效率非常低,通常认为环境中的甲基汞是由微生物主导产生[38-40].Jensen等[40]首次通过实验验证了沉积物中的甲基汞主要是由微生物甲基化产生的,主要的汞甲基化微生物包括硫酸盐还原菌(Sulphate reduction bacteria, SRB)、铁还原菌(Iron-reducing bacteria, FeRB)和产甲烷菌(Methanogenus)等.在过去很长一段时间内,对汞甲基化的遗传机制并不清楚.直到2013年美国橡树岭国家实验室研究人员报道了hgcA和hgcB两个基因簇,可以直接调控微生物汞甲基化,从而揭示了汞的微生物甲基化遗传机制[41].其中hgcA编码类咕啉蛋白,hgcB编码铁还原蛋白,这两种蛋白分别在甲基运输以及电子提供中起着重要的作用[41-42].研究表明,在所有已被证实的汞甲基化细菌中都存在这两个基因簇,hgcAB基因的发现成为可靠的甲基化微生物的分子标记物,可以预测汞甲基化过程的存在[41-43].目前使用这一分子探针已识别到30个门类的汞甲基化细菌,Gilmour等[43-44]还在许多新环境鉴定出了汞甲基化细菌以及除硫酸盐还原菌等常见菌种之外的许多细菌.随着对微生物甲基化分子机制理解的深入,可以根据环境中hgcAB基因的含量进行甲基汞风险预测,已有证据表明土壤中hgcA丰度与甲基汞浓度呈显著正相关[45]. ...
... [43-44]还在许多新环境鉴定出了汞甲基化细菌以及除硫酸盐还原菌等常见菌种之外的许多细菌.随着对微生物甲基化分子机制理解的深入,可以根据环境中hgcAB基因的含量进行甲基汞风险预测,已有证据表明土壤中hgcA丰度与甲基汞浓度呈显著正相关[45]. ...
... 早期对汞甲基化微生物的鉴定依赖于实验室培养,测试从厌氧沉积物中培养的分离物在添加无机汞后产生甲基汞的能力[64-65].然而,不同汞甲基化细菌的甲基化能力差异悬殊,如果忽视它是一个物种或菌种的特异性功能,使用这种方法预估环境中汞的甲基化潜力是欠准确的[43].目前主要通过16S rRNA基因的分析,评估微生物群落结构与环境中甲基汞和生物地球化学特性的关系[45,66-67].高通量测序作为一种新兴的免培养分子生物学技术,检测快速、准确、信息全面丰富,随着高通量测序技术的不断升级换代,测序通量、读长和准确度也有了很大提升,为认识各种环境中汞甲基化微生物多样性、群落结构组成及其生态功能提供了有利手段[68].Illumina公司的Solexa和Hiseq是目前全球使用量最大的第二代测序机器.现代分子生物学技术将从环境样品中提取的核酸(DNA或RNA)为分析对象,以进行目标片段的聚合酶链式反应(PCR)扩增,得到的PCR产物通过测序技术来鉴定微生物组成及其多样性.Mpbio公司的FastDNA Spin Kit for Soil试剂盒目前已广泛应用于DNA的提取,它是获得雪冰微生物高分子量、高纯度的基因组DNA非常重要的前提. ...
Expanded phylogenetic diversity and metabolic flexibility of mercury-methylating microorganisms
1
2020
... 汞的甲基化包括生物和非生物两种途径[5,37-40].在自然条件下汞的非生物甲基化效率非常低,通常认为环境中的甲基汞是由微生物主导产生[38-40].Jensen等[40]首次通过实验验证了沉积物中的甲基汞主要是由微生物甲基化产生的,主要的汞甲基化微生物包括硫酸盐还原菌(Sulphate reduction bacteria, SRB)、铁还原菌(Iron-reducing bacteria, FeRB)和产甲烷菌(Methanogenus)等.在过去很长一段时间内,对汞甲基化的遗传机制并不清楚.直到2013年美国橡树岭国家实验室研究人员报道了hgcA和hgcB两个基因簇,可以直接调控微生物汞甲基化,从而揭示了汞的微生物甲基化遗传机制[41].其中hgcA编码类咕啉蛋白,hgcB编码铁还原蛋白,这两种蛋白分别在甲基运输以及电子提供中起着重要的作用[41-42].研究表明,在所有已被证实的汞甲基化细菌中都存在这两个基因簇,hgcAB基因的发现成为可靠的甲基化微生物的分子标记物,可以预测汞甲基化过程的存在[41-43].目前使用这一分子探针已识别到30个门类的汞甲基化细菌,Gilmour等[43-44]还在许多新环境鉴定出了汞甲基化细菌以及除硫酸盐还原菌等常见菌种之外的许多细菌.随着对微生物甲基化分子机制理解的深入,可以根据环境中hgcAB基因的含量进行甲基汞风险预测,已有证据表明土壤中hgcA丰度与甲基汞浓度呈显著正相关[45]. ...
Analysis of the microbial community structure by monitoring an Hg methylation gene (hgcA) in paddy soils along an Hg gradient
2
2014
... 汞的甲基化包括生物和非生物两种途径[5,37-40].在自然条件下汞的非生物甲基化效率非常低,通常认为环境中的甲基汞是由微生物主导产生[38-40].Jensen等[40]首次通过实验验证了沉积物中的甲基汞主要是由微生物甲基化产生的,主要的汞甲基化微生物包括硫酸盐还原菌(Sulphate reduction bacteria, SRB)、铁还原菌(Iron-reducing bacteria, FeRB)和产甲烷菌(Methanogenus)等.在过去很长一段时间内,对汞甲基化的遗传机制并不清楚.直到2013年美国橡树岭国家实验室研究人员报道了hgcA和hgcB两个基因簇,可以直接调控微生物汞甲基化,从而揭示了汞的微生物甲基化遗传机制[41].其中hgcA编码类咕啉蛋白,hgcB编码铁还原蛋白,这两种蛋白分别在甲基运输以及电子提供中起着重要的作用[41-42].研究表明,在所有已被证实的汞甲基化细菌中都存在这两个基因簇,hgcAB基因的发现成为可靠的甲基化微生物的分子标记物,可以预测汞甲基化过程的存在[41-43].目前使用这一分子探针已识别到30个门类的汞甲基化细菌,Gilmour等[43-44]还在许多新环境鉴定出了汞甲基化细菌以及除硫酸盐还原菌等常见菌种之外的许多细菌.随着对微生物甲基化分子机制理解的深入,可以根据环境中hgcAB基因的含量进行甲基汞风险预测,已有证据表明土壤中hgcA丰度与甲基汞浓度呈显著正相关[45]. ...
... 早期对汞甲基化微生物的鉴定依赖于实验室培养,测试从厌氧沉积物中培养的分离物在添加无机汞后产生甲基汞的能力[64-65].然而,不同汞甲基化细菌的甲基化能力差异悬殊,如果忽视它是一个物种或菌种的特异性功能,使用这种方法预估环境中汞的甲基化潜力是欠准确的[43].目前主要通过16S rRNA基因的分析,评估微生物群落结构与环境中甲基汞和生物地球化学特性的关系[45,66-67].高通量测序作为一种新兴的免培养分子生物学技术,检测快速、准确、信息全面丰富,随着高通量测序技术的不断升级换代,测序通量、读长和准确度也有了很大提升,为认识各种环境中汞甲基化微生物多样性、群落结构组成及其生态功能提供了有利手段[68].Illumina公司的Solexa和Hiseq是目前全球使用量最大的第二代测序机器.现代分子生物学技术将从环境样品中提取的核酸(DNA或RNA)为分析对象,以进行目标片段的聚合酶链式反应(PCR)扩增,得到的PCR产物通过测序技术来鉴定微生物组成及其多样性.Mpbio公司的FastDNA Spin Kit for Soil试剂盒目前已广泛应用于DNA的提取,它是获得雪冰微生物高分子量、高纯度的基因组DNA非常重要的前提. ...
Mercury in the aquatic environment: a review of factors affecting methylation
2
2001
... 汞的生物甲基化是甲基汞的重要来源,它受到多种环境因素的控制,例如刺激无机汞的可用性、微生物群落以及基因调控的变化等都可以影响到汞生物甲基化[46-47].除了直接控制汞甲基化的基因hgcA和hgcB,还有一些通过控制环境中汞化合物形态和迁移转化进而间接影响甲基汞分布的基因[48].merA基因编码汞还原酶,介导的细菌能够将氧化态汞(Hg2+)还原为气态汞(Hg0),由于Hg2+是甲基化反应的底物,抗汞细菌亦可能在降低甲基化过程的底物供应方面发挥作用[48-49].merB基因编码烷基汞裂解酶,控制着去甲基化过程的发生,从而降低环境中甲基汞的总水平[48].鉴于影响甲基汞的环境因素众多且转化机理繁杂,本文仅重点关注冰冻圈汞的微生物甲基化,而对去甲基化等过程不予开展过多讨论. ...
... 多年冻土蕴藏着大量的汞存储量,有研究估算了北极苔原土壤储存汞约184 Gg;青藏高原活动层储存汞约16.6 Gg[16-17].冻土甲基汞的分布和转化等过程更为复杂,受各种物理、化学和生物因素控制,包括温度、溶解性有机质(DOC)、碱度、pH、总汞浓度、微生物以及其他金属元素或离子(如铁、锰)等[22,46,99-101].MacMillan等[22]发现冻土环境中甲基汞与溶解有机碳等营养物质,以及甲基汞与微生物之间有很强的相关性.当温度较高(8 ℃)时,在富含有机物和矿物质的土壤层中,甲基汞净产量比温度较低(-2 ℃)时增加了10倍以上[23].可溶性土壤有机碳类型及其生物可利用性,对促进甲基汞的初始合成特别重要[99].当冻土活动层融化时,低分子量溶解有机质增加促使形成大量生物可利用的Hg-DOM复合物,从而使无机汞的可利用性增加,更有利于冻土汞甲基化[23-24,100]. ...
Mercury methylation by anaerobic microorganisms: a review
1
2019
... 汞的生物甲基化是甲基汞的重要来源,它受到多种环境因素的控制,例如刺激无机汞的可用性、微生物群落以及基因调控的变化等都可以影响到汞生物甲基化[46-47].除了直接控制汞甲基化的基因hgcA和hgcB,还有一些通过控制环境中汞化合物形态和迁移转化进而间接影响甲基汞分布的基因[48].merA基因编码汞还原酶,介导的细菌能够将氧化态汞(Hg2+)还原为气态汞(Hg0),由于Hg2+是甲基化反应的底物,抗汞细菌亦可能在降低甲基化过程的底物供应方面发挥作用[48-49].merB基因编码烷基汞裂解酶,控制着去甲基化过程的发生,从而降低环境中甲基汞的总水平[48].鉴于影响甲基汞的环境因素众多且转化机理繁杂,本文仅重点关注冰冻圈汞的微生物甲基化,而对去甲基化等过程不予开展过多讨论. ...
Bacterial mercury resistance from atoms to ecosystems
4
2003
... 汞的生物甲基化是甲基汞的重要来源,它受到多种环境因素的控制,例如刺激无机汞的可用性、微生物群落以及基因调控的变化等都可以影响到汞生物甲基化[46-47].除了直接控制汞甲基化的基因hgcA和hgcB,还有一些通过控制环境中汞化合物形态和迁移转化进而间接影响甲基汞分布的基因[48].merA基因编码汞还原酶,介导的细菌能够将氧化态汞(Hg2+)还原为气态汞(Hg0),由于Hg2+是甲基化反应的底物,抗汞细菌亦可能在降低甲基化过程的底物供应方面发挥作用[48-49].merB基因编码烷基汞裂解酶,控制着去甲基化过程的发生,从而降低环境中甲基汞的总水平[48].鉴于影响甲基汞的环境因素众多且转化机理繁杂,本文仅重点关注冰冻圈汞的微生物甲基化,而对去甲基化等过程不予开展过多讨论. ...
... [48-49].merB基因编码烷基汞裂解酶,控制着去甲基化过程的发生,从而降低环境中甲基汞的总水平[48].鉴于影响甲基汞的环境因素众多且转化机理繁杂,本文仅重点关注冰冻圈汞的微生物甲基化,而对去甲基化等过程不予开展过多讨论. ...
... [48].鉴于影响甲基汞的环境因素众多且转化机理繁杂,本文仅重点关注冰冻圈汞的微生物甲基化,而对去甲基化等过程不予开展过多讨论. ...
... 对hgcAB基因簇、mer基因簇等分子层面的研究极大地拓宽了我们对汞甲基化机制的认识[41,48].Gionfriddo等[69]开发出了一套全新的引物,并将汞甲基化细菌基因序列参考库扩展到了283个,从而可以更准确地获取汞甲基化微生物多样性和环境中甲基汞产生的潜力.当今采用高通量测序获取基因和微生物多样性的生物学信息,开辟了冰冻圈汞甲基化研究的新领域.冰冻圈汞甲基化微生物分析方法概括归纳如图2所示.分析冰冻圈中hgcAB基因的方法一种是基于聚合酶链式反应技术,利用hgcAB特异引物获得样品中的hgcAB基因序列和丰度;另一种是通过BLAST方法从样本宏基因组数据中筛选出hgcAB同源序列[34]. ...
Diversity and characterization of mercury-resistant bacteria in snow, freshwater and sea-ice brine from the High Arctic
1
2011
... 汞的生物甲基化是甲基汞的重要来源,它受到多种环境因素的控制,例如刺激无机汞的可用性、微生物群落以及基因调控的变化等都可以影响到汞生物甲基化[46-47].除了直接控制汞甲基化的基因hgcA和hgcB,还有一些通过控制环境中汞化合物形态和迁移转化进而间接影响甲基汞分布的基因[48].merA基因编码汞还原酶,介导的细菌能够将氧化态汞(Hg2+)还原为气态汞(Hg0),由于Hg2+是甲基化反应的底物,抗汞细菌亦可能在降低甲基化过程的底物供应方面发挥作用[48-49].merB基因编码烷基汞裂解酶,控制着去甲基化过程的发生,从而降低环境中甲基汞的总水平[48].鉴于影响甲基汞的环境因素众多且转化机理繁杂,本文仅重点关注冰冻圈汞的微生物甲基化,而对去甲基化等过程不予开展过多讨论. ...
Clean hands, dirty hands: Clair Patterson and the aquatic biogeochemistry of mercury
1
1999
... 冰冻圈中甲基汞含量极低(pg级),因此要特别注意在采样和分析过程中潜在的汞污染问题,需严格遵循“clean hands-dirty hands”的操作规范[50].甲基汞浓度测定通常采用EPA推荐的方法(Method 1631)进行测试,即“蒸馏-乙基化-吹洗和捕获-冷原子荧光光谱法(CVAFS)”相结合的方法[51].样品在进行预处理后,还需要对汞形态进行分离,最常用的汞形态分离技术为气相色谱(GC)或高效液相色谱技术(HPLC).国内对甲基汞检测方法的研究起步较晚,早期一直采用气相色谱法测定甲基汞,但方法灵敏度较低[52].近些年来,气相色谱与冷原子荧光光谱法联用技术被广泛应用于各种环境样品甲基汞的测定[17,19].之后,电感耦合等离子体质谱法(ICP-MS)、同位素稀释-电感耦合等离子体质谱法(ID-CV-ICP-MS)以及同位素稀释固相微萃取-气相色谱-电感耦合等离子体质谱法(SPME-GC-ICP-MS)等手段也陆续被应用到甲基汞的检测分析[53-55].这些方法灵敏度高,检出限低,能很好地满足冰冻圈样品中低水平甲基汞的测定,目前常用的甲基汞测试方法列于表1. ...
1
2003
... 冰冻圈中甲基汞含量极低(pg级),因此要特别注意在采样和分析过程中潜在的汞污染问题,需严格遵循“clean hands-dirty hands”的操作规范[50].甲基汞浓度测定通常采用EPA推荐的方法(Method 1631)进行测试,即“蒸馏-乙基化-吹洗和捕获-冷原子荧光光谱法(CVAFS)”相结合的方法[51].样品在进行预处理后,还需要对汞形态进行分离,最常用的汞形态分离技术为气相色谱(GC)或高效液相色谱技术(HPLC).国内对甲基汞检测方法的研究起步较晚,早期一直采用气相色谱法测定甲基汞,但方法灵敏度较低[52].近些年来,气相色谱与冷原子荧光光谱法联用技术被广泛应用于各种环境样品甲基汞的测定[17,19].之后,电感耦合等离子体质谱法(ICP-MS)、同位素稀释-电感耦合等离子体质谱法(ID-CV-ICP-MS)以及同位素稀释固相微萃取-气相色谱-电感耦合等离子体质谱法(SPME-GC-ICP-MS)等手段也陆续被应用到甲基汞的检测分析[53-55].这些方法灵敏度高,检出限低,能很好地满足冰冻圈样品中低水平甲基汞的测定,目前常用的甲基汞测试方法列于表1. ...
Determination of methylmercury in sediment by Gas Chromatography with concentration of rotational sulfhydrylcotton yarn
1
1985
... 冰冻圈中甲基汞含量极低(pg级),因此要特别注意在采样和分析过程中潜在的汞污染问题,需严格遵循“clean hands-dirty hands”的操作规范[50].甲基汞浓度测定通常采用EPA推荐的方法(Method 1631)进行测试,即“蒸馏-乙基化-吹洗和捕获-冷原子荧光光谱法(CVAFS)”相结合的方法[51].样品在进行预处理后,还需要对汞形态进行分离,最常用的汞形态分离技术为气相色谱(GC)或高效液相色谱技术(HPLC).国内对甲基汞检测方法的研究起步较晚,早期一直采用气相色谱法测定甲基汞,但方法灵敏度较低[52].近些年来,气相色谱与冷原子荧光光谱法联用技术被广泛应用于各种环境样品甲基汞的测定[17,19].之后,电感耦合等离子体质谱法(ICP-MS)、同位素稀释-电感耦合等离子体质谱法(ID-CV-ICP-MS)以及同位素稀释固相微萃取-气相色谱-电感耦合等离子体质谱法(SPME-GC-ICP-MS)等手段也陆续被应用到甲基汞的检测分析[53-55].这些方法灵敏度高,检出限低,能很好地满足冰冻圈样品中低水平甲基汞的测定,目前常用的甲基汞测试方法列于表1. ...
巯基纱布旋转富集-气相色谱法测定沉积物中的甲基汞
1
1985
... 冰冻圈中甲基汞含量极低(pg级),因此要特别注意在采样和分析过程中潜在的汞污染问题,需严格遵循“clean hands-dirty hands”的操作规范[50].甲基汞浓度测定通常采用EPA推荐的方法(Method 1631)进行测试,即“蒸馏-乙基化-吹洗和捕获-冷原子荧光光谱法(CVAFS)”相结合的方法[51].样品在进行预处理后,还需要对汞形态进行分离,最常用的汞形态分离技术为气相色谱(GC)或高效液相色谱技术(HPLC).国内对甲基汞检测方法的研究起步较晚,早期一直采用气相色谱法测定甲基汞,但方法灵敏度较低[52].近些年来,气相色谱与冷原子荧光光谱法联用技术被广泛应用于各种环境样品甲基汞的测定[17,19].之后,电感耦合等离子体质谱法(ICP-MS)、同位素稀释-电感耦合等离子体质谱法(ID-CV-ICP-MS)以及同位素稀释固相微萃取-气相色谱-电感耦合等离子体质谱法(SPME-GC-ICP-MS)等手段也陆续被应用到甲基汞的检测分析[53-55].这些方法灵敏度高,检出限低,能很好地满足冰冻圈样品中低水平甲基汞的测定,目前常用的甲基汞测试方法列于表1. ...
The review of the analytical methods of methylmercury in environment samples
1
2004
... 冰冻圈中甲基汞含量极低(pg级),因此要特别注意在采样和分析过程中潜在的汞污染问题,需严格遵循“clean hands-dirty hands”的操作规范[50].甲基汞浓度测定通常采用EPA推荐的方法(Method 1631)进行测试,即“蒸馏-乙基化-吹洗和捕获-冷原子荧光光谱法(CVAFS)”相结合的方法[51].样品在进行预处理后,还需要对汞形态进行分离,最常用的汞形态分离技术为气相色谱(GC)或高效液相色谱技术(HPLC).国内对甲基汞检测方法的研究起步较晚,早期一直采用气相色谱法测定甲基汞,但方法灵敏度较低[52].近些年来,气相色谱与冷原子荧光光谱法联用技术被广泛应用于各种环境样品甲基汞的测定[17,19].之后,电感耦合等离子体质谱法(ICP-MS)、同位素稀释-电感耦合等离子体质谱法(ID-CV-ICP-MS)以及同位素稀释固相微萃取-气相色谱-电感耦合等离子体质谱法(SPME-GC-ICP-MS)等手段也陆续被应用到甲基汞的检测分析[53-55].这些方法灵敏度高,检出限低,能很好地满足冰冻圈样品中低水平甲基汞的测定,目前常用的甲基汞测试方法列于表1. ...
环境样品中甲基汞的分析方法综述
1
2004
... 冰冻圈中甲基汞含量极低(pg级),因此要特别注意在采样和分析过程中潜在的汞污染问题,需严格遵循“clean hands-dirty hands”的操作规范[50].甲基汞浓度测定通常采用EPA推荐的方法(Method 1631)进行测试,即“蒸馏-乙基化-吹洗和捕获-冷原子荧光光谱法(CVAFS)”相结合的方法[51].样品在进行预处理后,还需要对汞形态进行分离,最常用的汞形态分离技术为气相色谱(GC)或高效液相色谱技术(HPLC).国内对甲基汞检测方法的研究起步较晚,早期一直采用气相色谱法测定甲基汞,但方法灵敏度较低[52].近些年来,气相色谱与冷原子荧光光谱法联用技术被广泛应用于各种环境样品甲基汞的测定[17,19].之后,电感耦合等离子体质谱法(ICP-MS)、同位素稀释-电感耦合等离子体质谱法(ID-CV-ICP-MS)以及同位素稀释固相微萃取-气相色谱-电感耦合等离子体质谱法(SPME-GC-ICP-MS)等手段也陆续被应用到甲基汞的检测分析[53-55].这些方法灵敏度高,检出限低,能很好地满足冰冻圈样品中低水平甲基汞的测定,目前常用的甲基汞测试方法列于表1. ...
Determination of methylmercury in fish tissues by isotope dilution SPME-GC-ICP-MS
0
2003
The determination of methylmercury in biological samples by HPLC coupled to ICP‐MS detection
1
2007
... 冰冻圈中甲基汞含量极低(pg级),因此要特别注意在采样和分析过程中潜在的汞污染问题,需严格遵循“clean hands-dirty hands”的操作规范[50].甲基汞浓度测定通常采用EPA推荐的方法(Method 1631)进行测试,即“蒸馏-乙基化-吹洗和捕获-冷原子荧光光谱法(CVAFS)”相结合的方法[51].样品在进行预处理后,还需要对汞形态进行分离,最常用的汞形态分离技术为气相色谱(GC)或高效液相色谱技术(HPLC).国内对甲基汞检测方法的研究起步较晚,早期一直采用气相色谱法测定甲基汞,但方法灵敏度较低[52].近些年来,气相色谱与冷原子荧光光谱法联用技术被广泛应用于各种环境样品甲基汞的测定[17,19].之后,电感耦合等离子体质谱法(ICP-MS)、同位素稀释-电感耦合等离子体质谱法(ID-CV-ICP-MS)以及同位素稀释固相微萃取-气相色谱-电感耦合等离子体质谱法(SPME-GC-ICP-MS)等手段也陆续被应用到甲基汞的检测分析[53-55].这些方法灵敏度高,检出限低,能很好地满足冰冻圈样品中低水平甲基汞的测定,目前常用的甲基汞测试方法列于表1. ...
Microbial mercury methylation profile in terminus of a high-elevation glacier on the northern boundary of the Tibetan Plateau
3
2020
... Methods for the determination of MeHg in various environmental samples
Table 1| 样品类型 | 测定方法 | 检出限 | 文献来源 |
|---|
| 雪冰 | GC-CVAFS | 0.02 ng·L-1 | [56] |
| 雪冰 | GC-AFS | 0.002 ng·L-1 | [57] |
| 冻土 | HPIC-CVAFS | 0.018 ng·g-1 | [58] |
| 冻土 | CVAFS | 0.006 ng·g-1 | [17] |
| 海冰 | GC-ICP-MS | 0.02 ng·L-1 | [59] |
| 冰尘 | GC-CVAFS | 0.02 ng·g-1 | [19] |
| 融水 | GC-AFS | 0.02 ng·L-1 | [60] |
| 降水 | GC-CVAFS | 0.002 ng·L-1 | [61] |
| 鱼类 | GC-CVAFS | 0.12 ng·g-1 | [62] |
| 水稻 | GC-CVAFS | 0.003 ng·g-1 | [63] |
2.2 汞甲基化微生物早期对汞甲基化微生物的鉴定依赖于实验室培养,测试从厌氧沉积物中培养的分离物在添加无机汞后产生甲基汞的能力[64-65].然而,不同汞甲基化细菌的甲基化能力差异悬殊,如果忽视它是一个物种或菌种的特异性功能,使用这种方法预估环境中汞的甲基化潜力是欠准确的[43].目前主要通过16S rRNA基因的分析,评估微生物群落结构与环境中甲基汞和生物地球化学特性的关系[45,66-67].高通量测序作为一种新兴的免培养分子生物学技术,检测快速、准确、信息全面丰富,随着高通量测序技术的不断升级换代,测序通量、读长和准确度也有了很大提升,为认识各种环境中汞甲基化微生物多样性、群落结构组成及其生态功能提供了有利手段[68].Illumina公司的Solexa和Hiseq是目前全球使用量最大的第二代测序机器.现代分子生物学技术将从环境样品中提取的核酸(DNA或RNA)为分析对象,以进行目标片段的聚合酶链式反应(PCR)扩增,得到的PCR产物通过测序技术来鉴定微生物组成及其多样性.Mpbio公司的FastDNA Spin Kit for Soil试剂盒目前已广泛应用于DNA的提取,它是获得雪冰微生物高分子量、高纯度的基因组DNA非常重要的前提. ...
... 冰川作为冰冻圈重要组成因素,全球山地冰川(不包含格陵兰冰盖和南极冰盖)覆盖面积约为17×104 km2,共约11×104条冰川,主要分布在亚北极和青藏高原等地[10].早期Jitaru等[70]报道了欧洲阿尔卑斯山雪冰中溶解性甲基汞的含量(0.3~4.0 ng·L-1).近年来科研人员对我国青藏高原冰川也开展了研究工作,Huang等[19]对中国西部冰川上冰尘甲基汞含量进行测试分析,测得甲基汞平均含量为(1.0±0.4) ng·g-1,并初步估算出每年冰尘中的甲基汞总量约为(0.7±0.3) kg.微生物群落在汞的生物地球化学转化过程中非常活跃,冰川微生物活动可能发生汞微生物甲基化[71-74].大气汞沉降到青藏高原冰川之后在冰尘中积累,由于冰尘洞会发生季节性融化,这里便成为各种微生物群落的栖息地[75].前人研究指出我国西部冰川冰尘甲基汞与总汞的比值较高,说明冰尘表面可能是冰川汞甲基化的潜在位点[19].Zhang等[56]对青藏高原冰川开展了汞微生物甲基化初步研究,结合老虎沟12号冰川末端6个不同类型样品的汞形态和宏基因组学分析,发现在含大量碎屑和粉尘的尘埃层中汞浓度和汞甲基化基因(hgcA)丰度最高.不过该研究中汞甲基化细菌的未分类序列占比很高,微生物群落组成仍不明晰,尚未能全面地阐明汞甲基化的微生物学机制,需要进一步深入研究. ...
... Concentrations of MeHg in the cryospheric environment
Table 2| 地点 | 样品类型 | 甲基汞 | 总汞 | 文献来源 |
|---|
| Devon Island, Canada | 多年冻土 | 0.3~3.1 ng·g-1 | — | [58] |
| Barrow, Alaska | 热融湖塘 | 0.3~2.3 ng·L-1 | 25.3~141.2 ng·L-1 | [23] |
| Tibetan Plateau, China | 多年冻土 | — | 5.6~39.3 ng·g-1 | [17] |
| Fennoscandia | 多年冻土 | 0.004~28 ng·g-1 | 1.1~210 ng·g-1 | [24] |
| Tussock tundra, Alaska | 多年冻土 | — | 56~226 ng·g-1 | [16] |
| Laohugou No.12, Tibetan Plateau, China | 雪冰 | 0.005~0.5 ng·L-1 | 24.4~172.1 ng·L-1 | [56] |
| Tibetan Plateau, China | 冰尘 | 0.4~1.7 ng·g-1 | 17.9~114.5 ng·g-1 | [19] |
| French Alps | 表层雪 | 0.002~0.3 ng·L-1 | 1.8~169.5 ng·L-1 | [57] |
| Spitsbergen Island, Norway | 表层雪 | 0.004~0.5 ng·L-1 | 1~90 ng·L-1 | [123] |
| Resolute Bay, High Arctic, Canada | 表层雪 | <0.015~0.2 ng·L-1 | 2.6~150 ng·L-1 | [125] |
| Hudson Bay, Canada | 表层雪 | 0.04~0.14 ng·L-1 | 1.6~5.1 ng·L-1 | [120] |
| Alberta, Canada | 表层雪 | 0.02~0.3 ng·L-1 | 0.8~9.0 ng·L-1 | [137] |
| Antarctic | 积雪 | <0.02~0.3 ng·L-1 | 1.0~30.6 ng·L-1 | [59] |
| McMurdo/Ross Island, Antarctic | 积雪 | — | 40~430 ng·L-1 | [114] |
| Casey Station, Antarctic | 海冰 | <0.009~0.09 ng·L-1 | 0.2~5.7 ng·L-1 | [129] |
| Antarctic | 海冰 | <0.02~0.2 ng·L-1 | 1.4~179 ng·L-1 | [59] |
| Beaufort Sea and McClure Strait, Arctic | 海冰 | <0.02~0.5 ng·L-1 | 0.1~12.2 ng·L-1 | [28] |
| Southern Ocean | 海水 | <0.02~0.1 ng·L-1 | 0. 2~23 ng·L-1 | [59] |
| Arctic Ocean | 海水 | 0.006~0.07 ng·L-1 | 0.09~1.4 ng·L-1 | [135] |
| Resolute Bay, High Arctic, Canada | 海水 | 0.06~0.1 ng·L-1 | 0.1~0.2 ng·L-1 | [125] |
注:表格中海水均为海冰覆盖水域. ...
Total mercury and methylmercury in high altitude surface snow from the French Alps
2
2011
... Methods for the determination of MeHg in various environmental samples
Table 1| 样品类型 | 测定方法 | 检出限 | 文献来源 |
|---|
| 雪冰 | GC-CVAFS | 0.02 ng·L-1 | [56] |
| 雪冰 | GC-AFS | 0.002 ng·L-1 | [57] |
| 冻土 | HPIC-CVAFS | 0.018 ng·g-1 | [58] |
| 冻土 | CVAFS | 0.006 ng·g-1 | [17] |
| 海冰 | GC-ICP-MS | 0.02 ng·L-1 | [59] |
| 冰尘 | GC-CVAFS | 0.02 ng·g-1 | [19] |
| 融水 | GC-AFS | 0.02 ng·L-1 | [60] |
| 降水 | GC-CVAFS | 0.002 ng·L-1 | [61] |
| 鱼类 | GC-CVAFS | 0.12 ng·g-1 | [62] |
| 水稻 | GC-CVAFS | 0.003 ng·g-1 | [63] |
2.2 汞甲基化微生物早期对汞甲基化微生物的鉴定依赖于实验室培养,测试从厌氧沉积物中培养的分离物在添加无机汞后产生甲基汞的能力[64-65].然而,不同汞甲基化细菌的甲基化能力差异悬殊,如果忽视它是一个物种或菌种的特异性功能,使用这种方法预估环境中汞的甲基化潜力是欠准确的[43].目前主要通过16S rRNA基因的分析,评估微生物群落结构与环境中甲基汞和生物地球化学特性的关系[45,66-67].高通量测序作为一种新兴的免培养分子生物学技术,检测快速、准确、信息全面丰富,随着高通量测序技术的不断升级换代,测序通量、读长和准确度也有了很大提升,为认识各种环境中汞甲基化微生物多样性、群落结构组成及其生态功能提供了有利手段[68].Illumina公司的Solexa和Hiseq是目前全球使用量最大的第二代测序机器.现代分子生物学技术将从环境样品中提取的核酸(DNA或RNA)为分析对象,以进行目标片段的聚合酶链式反应(PCR)扩增,得到的PCR产物通过测序技术来鉴定微生物组成及其多样性.Mpbio公司的FastDNA Spin Kit for Soil试剂盒目前已广泛应用于DNA的提取,它是获得雪冰微生物高分子量、高纯度的基因组DNA非常重要的前提. ...
... Concentrations of MeHg in the cryospheric environment
Table 2| 地点 | 样品类型 | 甲基汞 | 总汞 | 文献来源 |
|---|
| Devon Island, Canada | 多年冻土 | 0.3~3.1 ng·g-1 | — | [58] |
| Barrow, Alaska | 热融湖塘 | 0.3~2.3 ng·L-1 | 25.3~141.2 ng·L-1 | [23] |
| Tibetan Plateau, China | 多年冻土 | — | 5.6~39.3 ng·g-1 | [17] |
| Fennoscandia | 多年冻土 | 0.004~28 ng·g-1 | 1.1~210 ng·g-1 | [24] |
| Tussock tundra, Alaska | 多年冻土 | — | 56~226 ng·g-1 | [16] |
| Laohugou No.12, Tibetan Plateau, China | 雪冰 | 0.005~0.5 ng·L-1 | 24.4~172.1 ng·L-1 | [56] |
| Tibetan Plateau, China | 冰尘 | 0.4~1.7 ng·g-1 | 17.9~114.5 ng·g-1 | [19] |
| French Alps | 表层雪 | 0.002~0.3 ng·L-1 | 1.8~169.5 ng·L-1 | [57] |
| Spitsbergen Island, Norway | 表层雪 | 0.004~0.5 ng·L-1 | 1~90 ng·L-1 | [123] |
| Resolute Bay, High Arctic, Canada | 表层雪 | <0.015~0.2 ng·L-1 | 2.6~150 ng·L-1 | [125] |
| Hudson Bay, Canada | 表层雪 | 0.04~0.14 ng·L-1 | 1.6~5.1 ng·L-1 | [120] |
| Alberta, Canada | 表层雪 | 0.02~0.3 ng·L-1 | 0.8~9.0 ng·L-1 | [137] |
| Antarctic | 积雪 | <0.02~0.3 ng·L-1 | 1.0~30.6 ng·L-1 | [59] |
| McMurdo/Ross Island, Antarctic | 积雪 | — | 40~430 ng·L-1 | [114] |
| Casey Station, Antarctic | 海冰 | <0.009~0.09 ng·L-1 | 0.2~5.7 ng·L-1 | [129] |
| Antarctic | 海冰 | <0.02~0.2 ng·L-1 | 1.4~179 ng·L-1 | [59] |
| Beaufort Sea and McClure Strait, Arctic | 海冰 | <0.02~0.5 ng·L-1 | 0.1~12.2 ng·L-1 | [28] |
| Southern Ocean | 海水 | <0.02~0.1 ng·L-1 | 0. 2~23 ng·L-1 | [59] |
| Arctic Ocean | 海水 | 0.006~0.07 ng·L-1 | 0.09~1.4 ng·L-1 | [135] |
| Resolute Bay, High Arctic, Canada | 海水 | 0.06~0.1 ng·L-1 | 0.1~0.2 ng·L-1 | [125] |
注:表格中海水均为海冰覆盖水域. ...
Methyl mercury production and loss in Arctic soil
4
2009
... Methods for the determination of MeHg in various environmental samples
Table 1| 样品类型 | 测定方法 | 检出限 | 文献来源 |
|---|
| 雪冰 | GC-CVAFS | 0.02 ng·L-1 | [56] |
| 雪冰 | GC-AFS | 0.002 ng·L-1 | [57] |
| 冻土 | HPIC-CVAFS | 0.018 ng·g-1 | [58] |
| 冻土 | CVAFS | 0.006 ng·g-1 | [17] |
| 海冰 | GC-ICP-MS | 0.02 ng·L-1 | [59] |
| 冰尘 | GC-CVAFS | 0.02 ng·g-1 | [19] |
| 融水 | GC-AFS | 0.02 ng·L-1 | [60] |
| 降水 | GC-CVAFS | 0.002 ng·L-1 | [61] |
| 鱼类 | GC-CVAFS | 0.12 ng·g-1 | [62] |
| 水稻 | GC-CVAFS | 0.003 ng·g-1 | [63] |
2.2 汞甲基化微生物早期对汞甲基化微生物的鉴定依赖于实验室培养,测试从厌氧沉积物中培养的分离物在添加无机汞后产生甲基汞的能力[64-65].然而,不同汞甲基化细菌的甲基化能力差异悬殊,如果忽视它是一个物种或菌种的特异性功能,使用这种方法预估环境中汞的甲基化潜力是欠准确的[43].目前主要通过16S rRNA基因的分析,评估微生物群落结构与环境中甲基汞和生物地球化学特性的关系[45,66-67].高通量测序作为一种新兴的免培养分子生物学技术,检测快速、准确、信息全面丰富,随着高通量测序技术的不断升级换代,测序通量、读长和准确度也有了很大提升,为认识各种环境中汞甲基化微生物多样性、群落结构组成及其生态功能提供了有利手段[68].Illumina公司的Solexa和Hiseq是目前全球使用量最大的第二代测序机器.现代分子生物学技术将从环境样品中提取的核酸(DNA或RNA)为分析对象,以进行目标片段的聚合酶链式反应(PCR)扩增,得到的PCR产物通过测序技术来鉴定微生物组成及其多样性.Mpbio公司的FastDNA Spin Kit for Soil试剂盒目前已广泛应用于DNA的提取,它是获得雪冰微生物高分子量、高纯度的基因组DNA非常重要的前提. ...
... 由于气候变暖,多年冻土快速融化、塌陷,形成热融喀斯特地貌,其中最显著的是形成热融湖塘,此过程更有利于微生物将无机汞转化为甲基汞[22-24,102-104].Tarbier等[24]在亚北极多年冻土区测量了178个土壤样品,甲基汞的浓度范围为0.01~28 ng·g-1,与其他地区土壤甲基汞相比要高得多(例如阿拉斯加北部<0.1~3.4 ng·g-1),同时发现热融池水和热岩孔隙水中甲基汞含量亦较高(0.01~3.1 ng·L-1)[22,24,58,100].在融化阶段,塌陷的冻土沼泽地比完整冻土带活动层的净甲基化程度还要高一个数量级(甲基汞占总汞的比例高出13倍),表明当热融湖塘形成时,冻土区的汞甲基化率将会显著提高[24].这主要是由于冻融塌陷使得冻土泥炭表面被淹没在水位以下,从而形成缺氧环境,这种条件非常有利于具有汞甲基化能力的厌氧微生物生存(包括硫酸盐还原菌、铁还原菌及产甲烷菌等)[58,91,100,104-105]. ...
... [58,91,100,104-105]. ...
... Concentrations of MeHg in the cryospheric environment
Table 2| 地点 | 样品类型 | 甲基汞 | 总汞 | 文献来源 |
|---|
| Devon Island, Canada | 多年冻土 | 0.3~3.1 ng·g-1 | — | [58] |
| Barrow, Alaska | 热融湖塘 | 0.3~2.3 ng·L-1 | 25.3~141.2 ng·L-1 | [23] |
| Tibetan Plateau, China | 多年冻土 | — | 5.6~39.3 ng·g-1 | [17] |
| Fennoscandia | 多年冻土 | 0.004~28 ng·g-1 | 1.1~210 ng·g-1 | [24] |
| Tussock tundra, Alaska | 多年冻土 | — | 56~226 ng·g-1 | [16] |
| Laohugou No.12, Tibetan Plateau, China | 雪冰 | 0.005~0.5 ng·L-1 | 24.4~172.1 ng·L-1 | [56] |
| Tibetan Plateau, China | 冰尘 | 0.4~1.7 ng·g-1 | 17.9~114.5 ng·g-1 | [19] |
| French Alps | 表层雪 | 0.002~0.3 ng·L-1 | 1.8~169.5 ng·L-1 | [57] |
| Spitsbergen Island, Norway | 表层雪 | 0.004~0.5 ng·L-1 | 1~90 ng·L-1 | [123] |
| Resolute Bay, High Arctic, Canada | 表层雪 | <0.015~0.2 ng·L-1 | 2.6~150 ng·L-1 | [125] |
| Hudson Bay, Canada | 表层雪 | 0.04~0.14 ng·L-1 | 1.6~5.1 ng·L-1 | [120] |
| Alberta, Canada | 表层雪 | 0.02~0.3 ng·L-1 | 0.8~9.0 ng·L-1 | [137] |
| Antarctic | 积雪 | <0.02~0.3 ng·L-1 | 1.0~30.6 ng·L-1 | [59] |
| McMurdo/Ross Island, Antarctic | 积雪 | — | 40~430 ng·L-1 | [114] |
| Casey Station, Antarctic | 海冰 | <0.009~0.09 ng·L-1 | 0.2~5.7 ng·L-1 | [129] |
| Antarctic | 海冰 | <0.02~0.2 ng·L-1 | 1.4~179 ng·L-1 | [59] |
| Beaufort Sea and McClure Strait, Arctic | 海冰 | <0.02~0.5 ng·L-1 | 0.1~12.2 ng·L-1 | [28] |
| Southern Ocean | 海水 | <0.02~0.1 ng·L-1 | 0. 2~23 ng·L-1 | [59] |
| Arctic Ocean | 海水 | 0.006~0.07 ng·L-1 | 0.09~1.4 ng·L-1 | [135] |
| Resolute Bay, High Arctic, Canada | 海水 | 0.06~0.1 ng·L-1 | 0.1~0.2 ng·L-1 | [125] |
注:表格中海水均为海冰覆盖水域. ...
Microbial mercury methylation in Antarctic sea ice
8
2016
... Methods for the determination of MeHg in various environmental samples
Table 1| 样品类型 | 测定方法 | 检出限 | 文献来源 |
|---|
| 雪冰 | GC-CVAFS | 0.02 ng·L-1 | [56] |
| 雪冰 | GC-AFS | 0.002 ng·L-1 | [57] |
| 冻土 | HPIC-CVAFS | 0.018 ng·g-1 | [58] |
| 冻土 | CVAFS | 0.006 ng·g-1 | [17] |
| 海冰 | GC-ICP-MS | 0.02 ng·L-1 | [59] |
| 冰尘 | GC-CVAFS | 0.02 ng·g-1 | [19] |
| 融水 | GC-AFS | 0.02 ng·L-1 | [60] |
| 降水 | GC-CVAFS | 0.002 ng·L-1 | [61] |
| 鱼类 | GC-CVAFS | 0.12 ng·g-1 | [62] |
| 水稻 | GC-CVAFS | 0.003 ng·g-1 | [63] |
2.2 汞甲基化微生物早期对汞甲基化微生物的鉴定依赖于实验室培养,测试从厌氧沉积物中培养的分离物在添加无机汞后产生甲基汞的能力[64-65].然而,不同汞甲基化细菌的甲基化能力差异悬殊,如果忽视它是一个物种或菌种的特异性功能,使用这种方法预估环境中汞的甲基化潜力是欠准确的[43].目前主要通过16S rRNA基因的分析,评估微生物群落结构与环境中甲基汞和生物地球化学特性的关系[45,66-67].高通量测序作为一种新兴的免培养分子生物学技术,检测快速、准确、信息全面丰富,随着高通量测序技术的不断升级换代,测序通量、读长和准确度也有了很大提升,为认识各种环境中汞甲基化微生物多样性、群落结构组成及其生态功能提供了有利手段[68].Illumina公司的Solexa和Hiseq是目前全球使用量最大的第二代测序机器.现代分子生物学技术将从环境样品中提取的核酸(DNA或RNA)为分析对象,以进行目标片段的聚合酶链式反应(PCR)扩增,得到的PCR产物通过测序技术来鉴定微生物组成及其多样性.Mpbio公司的FastDNA Spin Kit for Soil试剂盒目前已广泛应用于DNA的提取,它是获得雪冰微生物高分子量、高纯度的基因组DNA非常重要的前提. ...
... 最新的研究不断揭示出潜在的新型海洋汞甲基化微生物,它们具有更强的耐氧性和更广泛的生境范围,这些都是有利于海冰汞甲基化的理想条件[59].Gionfriddo等[59]通过对南极海冰汞甲基化研究表明,南极海冰中存在甲基汞的富集现象,主要是由微嗜氧细菌-硝化刺菌属(Nitrospina)主导产生的,参与调控微生物汞甲基化的基因为hgcA-like.这与之前的认知有所不同,过去认为只有厌氧条件支持显著的汞甲基化过程,且汞甲基化微生物都属于厌氧细菌.因此,海冰汞的研究不仅揭示了更多潜在的新型海洋汞甲基化微生物,而且极大地拓宽了我们对汞甲基化微生物耐氧性的认识.气候变暖背景下,海冰变薄、季节性海冰区扩大、表层海洋分层加剧和浮游生物生态动力学的变化,这些过程都会造成海洋甲基汞产量增加[26,136].从海冰融化释放出来的大量甲基汞进入海洋生态系统之后会在食物链中富集和放大,将会对南北极水生食物链中高营养级的动物构成很大威胁[26,28,59,134].通过对北极多年海冰的融化情况的估算,每年输入北冰洋的总汞和甲基汞的通量估计分别为420 kg·a-1和42 kg·a-1[28].南极海冰融化亦会给海洋分别带来超过9 600 kg·a-1总汞和18 kg·a-1甲基汞,总汞的估算值要比北冰洋高出一个数量级[59,134].为方便后续研究,将冰冻圈总汞和甲基汞浓度水平相关信息汇总于表2. ...
... [59]通过对南极海冰汞甲基化研究表明,南极海冰中存在甲基汞的富集现象,主要是由微嗜氧细菌-硝化刺菌属(Nitrospina)主导产生的,参与调控微生物汞甲基化的基因为hgcA-like.这与之前的认知有所不同,过去认为只有厌氧条件支持显著的汞甲基化过程,且汞甲基化微生物都属于厌氧细菌.因此,海冰汞的研究不仅揭示了更多潜在的新型海洋汞甲基化微生物,而且极大地拓宽了我们对汞甲基化微生物耐氧性的认识.气候变暖背景下,海冰变薄、季节性海冰区扩大、表层海洋分层加剧和浮游生物生态动力学的变化,这些过程都会造成海洋甲基汞产量增加[26,136].从海冰融化释放出来的大量甲基汞进入海洋生态系统之后会在食物链中富集和放大,将会对南北极水生食物链中高营养级的动物构成很大威胁[26,28,59,134].通过对北极多年海冰的融化情况的估算,每年输入北冰洋的总汞和甲基汞的通量估计分别为420 kg·a-1和42 kg·a-1[28].南极海冰融化亦会给海洋分别带来超过9 600 kg·a-1总汞和18 kg·a-1甲基汞,总汞的估算值要比北冰洋高出一个数量级[59,134].为方便后续研究,将冰冻圈总汞和甲基汞浓度水平相关信息汇总于表2. ...
... ,59,134].通过对北极多年海冰的融化情况的估算,每年输入北冰洋的总汞和甲基汞的通量估计分别为420 kg·a-1和42 kg·a-1[28].南极海冰融化亦会给海洋分别带来超过9 600 kg·a-1总汞和18 kg·a-1甲基汞,总汞的估算值要比北冰洋高出一个数量级[59,134].为方便后续研究,将冰冻圈总汞和甲基汞浓度水平相关信息汇总于表2. ...
... [59,134].为方便后续研究,将冰冻圈总汞和甲基汞浓度水平相关信息汇总于表2. ...
... Concentrations of MeHg in the cryospheric environment
Table 2| 地点 | 样品类型 | 甲基汞 | 总汞 | 文献来源 |
|---|
| Devon Island, Canada | 多年冻土 | 0.3~3.1 ng·g-1 | — | [58] |
| Barrow, Alaska | 热融湖塘 | 0.3~2.3 ng·L-1 | 25.3~141.2 ng·L-1 | [23] |
| Tibetan Plateau, China | 多年冻土 | — | 5.6~39.3 ng·g-1 | [17] |
| Fennoscandia | 多年冻土 | 0.004~28 ng·g-1 | 1.1~210 ng·g-1 | [24] |
| Tussock tundra, Alaska | 多年冻土 | — | 56~226 ng·g-1 | [16] |
| Laohugou No.12, Tibetan Plateau, China | 雪冰 | 0.005~0.5 ng·L-1 | 24.4~172.1 ng·L-1 | [56] |
| Tibetan Plateau, China | 冰尘 | 0.4~1.7 ng·g-1 | 17.9~114.5 ng·g-1 | [19] |
| French Alps | 表层雪 | 0.002~0.3 ng·L-1 | 1.8~169.5 ng·L-1 | [57] |
| Spitsbergen Island, Norway | 表层雪 | 0.004~0.5 ng·L-1 | 1~90 ng·L-1 | [123] |
| Resolute Bay, High Arctic, Canada | 表层雪 | <0.015~0.2 ng·L-1 | 2.6~150 ng·L-1 | [125] |
| Hudson Bay, Canada | 表层雪 | 0.04~0.14 ng·L-1 | 1.6~5.1 ng·L-1 | [120] |
| Alberta, Canada | 表层雪 | 0.02~0.3 ng·L-1 | 0.8~9.0 ng·L-1 | [137] |
| Antarctic | 积雪 | <0.02~0.3 ng·L-1 | 1.0~30.6 ng·L-1 | [59] |
| McMurdo/Ross Island, Antarctic | 积雪 | — | 40~430 ng·L-1 | [114] |
| Casey Station, Antarctic | 海冰 | <0.009~0.09 ng·L-1 | 0.2~5.7 ng·L-1 | [129] |
| Antarctic | 海冰 | <0.02~0.2 ng·L-1 | 1.4~179 ng·L-1 | [59] |
| Beaufort Sea and McClure Strait, Arctic | 海冰 | <0.02~0.5 ng·L-1 | 0.1~12.2 ng·L-1 | [28] |
| Southern Ocean | 海水 | <0.02~0.1 ng·L-1 | 0. 2~23 ng·L-1 | [59] |
| Arctic Ocean | 海水 | 0.006~0.07 ng·L-1 | 0.09~1.4 ng·L-1 | [135] |
| Resolute Bay, High Arctic, Canada | 海水 | 0.06~0.1 ng·L-1 | 0.1~0.2 ng·L-1 | [125] |
注:表格中海水均为海冰覆盖水域. ...
... [
59]
| Beaufort Sea and McClure Strait, Arctic | 海冰 | <0.02~0.5 ng·L-1 | 0.1~12.2 ng·L-1 | [28] |
| Southern Ocean | 海水 | <0.02~0.1 ng·L-1 | 0. 2~23 ng·L-1 | [59] |
| Arctic Ocean | 海水 | 0.006~0.07 ng·L-1 | 0.09~1.4 ng·L-1 | [135] |
| Resolute Bay, High Arctic, Canada | 海水 | 0.06~0.1 ng·L-1 | 0.1~0.2 ng·L-1 | [125] |
注:表格中海水均为海冰覆盖水域. ...
... [
59]
| Arctic Ocean | 海水 | 0.006~0.07 ng·L-1 | 0.09~1.4 ng·L-1 | [135] |
| Resolute Bay, High Arctic, Canada | 海水 | 0.06~0.1 ng·L-1 | 0.1~0.2 ng·L-1 | [125] |
注:表格中海水均为海冰覆盖水域. ...
Snowmelt sources of methylmercury to high arctic ecosystems
3
2004
... Methods for the determination of MeHg in various environmental samples
Table 1| 样品类型 | 测定方法 | 检出限 | 文献来源 |
|---|
| 雪冰 | GC-CVAFS | 0.02 ng·L-1 | [56] |
| 雪冰 | GC-AFS | 0.002 ng·L-1 | [57] |
| 冻土 | HPIC-CVAFS | 0.018 ng·g-1 | [58] |
| 冻土 | CVAFS | 0.006 ng·g-1 | [17] |
| 海冰 | GC-ICP-MS | 0.02 ng·L-1 | [59] |
| 冰尘 | GC-CVAFS | 0.02 ng·g-1 | [19] |
| 融水 | GC-AFS | 0.02 ng·L-1 | [60] |
| 降水 | GC-CVAFS | 0.002 ng·L-1 | [61] |
| 鱼类 | GC-CVAFS | 0.12 ng·g-1 | [62] |
| 水稻 | GC-CVAFS | 0.003 ng·g-1 | [63] |
2.2 汞甲基化微生物早期对汞甲基化微生物的鉴定依赖于实验室培养,测试从厌氧沉积物中培养的分离物在添加无机汞后产生甲基汞的能力[64-65].然而,不同汞甲基化细菌的甲基化能力差异悬殊,如果忽视它是一个物种或菌种的特异性功能,使用这种方法预估环境中汞的甲基化潜力是欠准确的[43].目前主要通过16S rRNA基因的分析,评估微生物群落结构与环境中甲基汞和生物地球化学特性的关系[45,66-67].高通量测序作为一种新兴的免培养分子生物学技术,检测快速、准确、信息全面丰富,随着高通量测序技术的不断升级换代,测序通量、读长和准确度也有了很大提升,为认识各种环境中汞甲基化微生物多样性、群落结构组成及其生态功能提供了有利手段[68].Illumina公司的Solexa和Hiseq是目前全球使用量最大的第二代测序机器.现代分子生物学技术将从环境样品中提取的核酸(DNA或RNA)为分析对象,以进行目标片段的聚合酶链式反应(PCR)扩增,得到的PCR产物通过测序技术来鉴定微生物组成及其多样性.Mpbio公司的FastDNA Spin Kit for Soil试剂盒目前已广泛应用于DNA的提取,它是获得雪冰微生物高分子量、高纯度的基因组DNA非常重要的前提. ...
... 降雪是冰冻圈大气降水最重要的表现形式,是大气汞进入冰冻圈的重要环节[108].积雪亦是一种活跃的媒介,消融后将成为环境汞的重要来源[60,109-110].在南北极地区,南极(0.1~1 ng·L-1)与北极(1~5 ng·L-1)表层雪冰中的总汞浓度在冬季均很低;然而在春季,受大气汞亏损事件(Atmospheric Mercury Depletion Events, AMDEs)的影响,南北极雪冰中汞的浓度均显著增加[111-119].早期研究表明积雪中甲基汞与大气沉降密切相关,但在AMDEs期间甲基汞与无机汞表现出完全不同的行为模式[108,111].Constant等[120]调查了加拿大亚北极地区AMDEs时期雪冰中的甲基汞变化规律,结果表明甲基汞的来源与AMDEs无关.雪冰甲基汞与氯化物之间存在较强的相关性,表明沿海地区积雪甲基汞可能来自海盐气溶胶[108].海洋中浮游植物可以产生二甲基汞(DMeHg),由于二甲基汞具有较高的挥发性,它可以从海洋释放到大气中,被自由基氧化成单甲基汞,然后随海盐沉降到附近的雪面之上[108,121-123].值得注意的是新沉降的甲基汞在积雪中并不稳定,15%~56%的甲基汞会在夜间通过去甲基化或以其他方式损失[124]. ...
... 积雪中甲基汞的来源在过去被归因于大气沉降,然而随着研究的不断深入,新的证据表明在低纬度苔原的积雪中可能存在着较强的汞甲基化活动[108,111,123-127].在融雪期间,雪冰融水中甲基汞占总汞的比例增大(从2.7%增加到7.6%)[126].导致这一现象的原因一方面由于甲基汞从无机/有机物中浸出,在雪冰融化时趋于集中;另一方面是由于雪冰内部可能发生甲基化作用,微生物活动和反应可能在积雪中产生了甲基汞[108,120,123-127].极地积雪的pH值通常为酸性,有机物以及微生物等存在于雪层中,因此在消融的积雪中存在微生物汞甲基化的可能[120].随着气候变暖,温度升高会促使积雪中微生物种群数量增长,从而导致微生物影响汞甲基化的强度大幅增加[25].Loseto等[60]估算出北极地区积雪中甲基汞产量约为1.5 mg·km-2·d-1,并指出北极康沃利斯岛陆地积雪融化是地表径流中最重要的甲基汞来源. ...
Characterizations of wet mercury deposition on a remote high-elevation site in the southeastern Tibetan Plateau
1
2015
... Methods for the determination of MeHg in various environmental samples
Table 1| 样品类型 | 测定方法 | 检出限 | 文献来源 |
|---|
| 雪冰 | GC-CVAFS | 0.02 ng·L-1 | [56] |
| 雪冰 | GC-AFS | 0.002 ng·L-1 | [57] |
| 冻土 | HPIC-CVAFS | 0.018 ng·g-1 | [58] |
| 冻土 | CVAFS | 0.006 ng·g-1 | [17] |
| 海冰 | GC-ICP-MS | 0.02 ng·L-1 | [59] |
| 冰尘 | GC-CVAFS | 0.02 ng·g-1 | [19] |
| 融水 | GC-AFS | 0.02 ng·L-1 | [60] |
| 降水 | GC-CVAFS | 0.002 ng·L-1 | [61] |
| 鱼类 | GC-CVAFS | 0.12 ng·g-1 | [62] |
| 水稻 | GC-CVAFS | 0.003 ng·g-1 | [63] |
2.2 汞甲基化微生物早期对汞甲基化微生物的鉴定依赖于实验室培养,测试从厌氧沉积物中培养的分离物在添加无机汞后产生甲基汞的能力[64-65].然而,不同汞甲基化细菌的甲基化能力差异悬殊,如果忽视它是一个物种或菌种的特异性功能,使用这种方法预估环境中汞的甲基化潜力是欠准确的[43].目前主要通过16S rRNA基因的分析,评估微生物群落结构与环境中甲基汞和生物地球化学特性的关系[45,66-67].高通量测序作为一种新兴的免培养分子生物学技术,检测快速、准确、信息全面丰富,随着高通量测序技术的不断升级换代,测序通量、读长和准确度也有了很大提升,为认识各种环境中汞甲基化微生物多样性、群落结构组成及其生态功能提供了有利手段[68].Illumina公司的Solexa和Hiseq是目前全球使用量最大的第二代测序机器.现代分子生物学技术将从环境样品中提取的核酸(DNA或RNA)为分析对象,以进行目标片段的聚合酶链式反应(PCR)扩增,得到的PCR产物通过测序技术来鉴定微生物组成及其多样性.Mpbio公司的FastDNA Spin Kit for Soil试剂盒目前已广泛应用于DNA的提取,它是获得雪冰微生物高分子量、高纯度的基因组DNA非常重要的前提. ...
Percentage of methylmercury in the muscle tissue of freshwater fish varies with body size and age and among species
1
2018
... Methods for the determination of MeHg in various environmental samples
Table 1| 样品类型 | 测定方法 | 检出限 | 文献来源 |
|---|
| 雪冰 | GC-CVAFS | 0.02 ng·L-1 | [56] |
| 雪冰 | GC-AFS | 0.002 ng·L-1 | [57] |
| 冻土 | HPIC-CVAFS | 0.018 ng·g-1 | [58] |
| 冻土 | CVAFS | 0.006 ng·g-1 | [17] |
| 海冰 | GC-ICP-MS | 0.02 ng·L-1 | [59] |
| 冰尘 | GC-CVAFS | 0.02 ng·g-1 | [19] |
| 融水 | GC-AFS | 0.02 ng·L-1 | [60] |
| 降水 | GC-CVAFS | 0.002 ng·L-1 | [61] |
| 鱼类 | GC-CVAFS | 0.12 ng·g-1 | [62] |
| 水稻 | GC-CVAFS | 0.003 ng·g-1 | [63] |
2.2 汞甲基化微生物早期对汞甲基化微生物的鉴定依赖于实验室培养,测试从厌氧沉积物中培养的分离物在添加无机汞后产生甲基汞的能力[64-65].然而,不同汞甲基化细菌的甲基化能力差异悬殊,如果忽视它是一个物种或菌种的特异性功能,使用这种方法预估环境中汞的甲基化潜力是欠准确的[43].目前主要通过16S rRNA基因的分析,评估微生物群落结构与环境中甲基汞和生物地球化学特性的关系[45,66-67].高通量测序作为一种新兴的免培养分子生物学技术,检测快速、准确、信息全面丰富,随着高通量测序技术的不断升级换代,测序通量、读长和准确度也有了很大提升,为认识各种环境中汞甲基化微生物多样性、群落结构组成及其生态功能提供了有利手段[68].Illumina公司的Solexa和Hiseq是目前全球使用量最大的第二代测序机器.现代分子生物学技术将从环境样品中提取的核酸(DNA或RNA)为分析对象,以进行目标片段的聚合酶链式反应(PCR)扩增,得到的PCR产物通过测序技术来鉴定微生物组成及其多样性.Mpbio公司的FastDNA Spin Kit for Soil试剂盒目前已广泛应用于DNA的提取,它是获得雪冰微生物高分子量、高纯度的基因组DNA非常重要的前提. ...
Methylmercury in rice (Oryza sativa L.) grown from the Xunyang Hg Mining area, Shaanxi Province, northwestern China
1
2011
... Methods for the determination of MeHg in various environmental samples
Table 1| 样品类型 | 测定方法 | 检出限 | 文献来源 |
|---|
| 雪冰 | GC-CVAFS | 0.02 ng·L-1 | [56] |
| 雪冰 | GC-AFS | 0.002 ng·L-1 | [57] |
| 冻土 | HPIC-CVAFS | 0.018 ng·g-1 | [58] |
| 冻土 | CVAFS | 0.006 ng·g-1 | [17] |
| 海冰 | GC-ICP-MS | 0.02 ng·L-1 | [59] |
| 冰尘 | GC-CVAFS | 0.02 ng·g-1 | [19] |
| 融水 | GC-AFS | 0.02 ng·L-1 | [60] |
| 降水 | GC-CVAFS | 0.002 ng·L-1 | [61] |
| 鱼类 | GC-CVAFS | 0.12 ng·g-1 | [62] |
| 水稻 | GC-CVAFS | 0.003 ng·g-1 | [63] |
2.2 汞甲基化微生物早期对汞甲基化微生物的鉴定依赖于实验室培养,测试从厌氧沉积物中培养的分离物在添加无机汞后产生甲基汞的能力[64-65].然而,不同汞甲基化细菌的甲基化能力差异悬殊,如果忽视它是一个物种或菌种的特异性功能,使用这种方法预估环境中汞的甲基化潜力是欠准确的[43].目前主要通过16S rRNA基因的分析,评估微生物群落结构与环境中甲基汞和生物地球化学特性的关系[45,66-67].高通量测序作为一种新兴的免培养分子生物学技术,检测快速、准确、信息全面丰富,随着高通量测序技术的不断升级换代,测序通量、读长和准确度也有了很大提升,为认识各种环境中汞甲基化微生物多样性、群落结构组成及其生态功能提供了有利手段[68].Illumina公司的Solexa和Hiseq是目前全球使用量最大的第二代测序机器.现代分子生物学技术将从环境样品中提取的核酸(DNA或RNA)为分析对象,以进行目标片段的聚合酶链式反应(PCR)扩增,得到的PCR产物通过测序技术来鉴定微生物组成及其多样性.Mpbio公司的FastDNA Spin Kit for Soil试剂盒目前已广泛应用于DNA的提取,它是获得雪冰微生物高分子量、高纯度的基因组DNA非常重要的前提. ...
Sulfate-reducing bacteria: principal methylators of mercury in anoxic estuarine sediment
1
1985
... 早期对汞甲基化微生物的鉴定依赖于实验室培养,测试从厌氧沉积物中培养的分离物在添加无机汞后产生甲基汞的能力[64-65].然而,不同汞甲基化细菌的甲基化能力差异悬殊,如果忽视它是一个物种或菌种的特异性功能,使用这种方法预估环境中汞的甲基化潜力是欠准确的[43].目前主要通过16S rRNA基因的分析,评估微生物群落结构与环境中甲基汞和生物地球化学特性的关系[45,66-67].高通量测序作为一种新兴的免培养分子生物学技术,检测快速、准确、信息全面丰富,随着高通量测序技术的不断升级换代,测序通量、读长和准确度也有了很大提升,为认识各种环境中汞甲基化微生物多样性、群落结构组成及其生态功能提供了有利手段[68].Illumina公司的Solexa和Hiseq是目前全球使用量最大的第二代测序机器.现代分子生物学技术将从环境样品中提取的核酸(DNA或RNA)为分析对象,以进行目标片段的聚合酶链式反应(PCR)扩增,得到的PCR产物通过测序技术来鉴定微生物组成及其多样性.Mpbio公司的FastDNA Spin Kit for Soil试剂盒目前已广泛应用于DNA的提取,它是获得雪冰微生物高分子量、高纯度的基因组DNA非常重要的前提. ...
Mercury methylation from unexpected sources: molybdate-inhibited freshwater sediments and an iron-reducing bacterium
1
2006
... 早期对汞甲基化微生物的鉴定依赖于实验室培养,测试从厌氧沉积物中培养的分离物在添加无机汞后产生甲基汞的能力[64-65].然而,不同汞甲基化细菌的甲基化能力差异悬殊,如果忽视它是一个物种或菌种的特异性功能,使用这种方法预估环境中汞的甲基化潜力是欠准确的[43].目前主要通过16S rRNA基因的分析,评估微生物群落结构与环境中甲基汞和生物地球化学特性的关系[45,66-67].高通量测序作为一种新兴的免培养分子生物学技术,检测快速、准确、信息全面丰富,随着高通量测序技术的不断升级换代,测序通量、读长和准确度也有了很大提升,为认识各种环境中汞甲基化微生物多样性、群落结构组成及其生态功能提供了有利手段[68].Illumina公司的Solexa和Hiseq是目前全球使用量最大的第二代测序机器.现代分子生物学技术将从环境样品中提取的核酸(DNA或RNA)为分析对象,以进行目标片段的聚合酶链式反应(PCR)扩增,得到的PCR产物通过测序技术来鉴定微生物组成及其多样性.Mpbio公司的FastDNA Spin Kit for Soil试剂盒目前已广泛应用于DNA的提取,它是获得雪冰微生物高分子量、高纯度的基因组DNA非常重要的前提. ...
Depth profile of sulfate-reducing bacterial ribosomal RNA and mercury methylation in an estuarine sediment
1
1996
... 早期对汞甲基化微生物的鉴定依赖于实验室培养,测试从厌氧沉积物中培养的分离物在添加无机汞后产生甲基汞的能力[64-65].然而,不同汞甲基化细菌的甲基化能力差异悬殊,如果忽视它是一个物种或菌种的特异性功能,使用这种方法预估环境中汞的甲基化潜力是欠准确的[43].目前主要通过16S rRNA基因的分析,评估微生物群落结构与环境中甲基汞和生物地球化学特性的关系[45,66-67].高通量测序作为一种新兴的免培养分子生物学技术,检测快速、准确、信息全面丰富,随着高通量测序技术的不断升级换代,测序通量、读长和准确度也有了很大提升,为认识各种环境中汞甲基化微生物多样性、群落结构组成及其生态功能提供了有利手段[68].Illumina公司的Solexa和Hiseq是目前全球使用量最大的第二代测序机器.现代分子生物学技术将从环境样品中提取的核酸(DNA或RNA)为分析对象,以进行目标片段的聚合酶链式反应(PCR)扩增,得到的PCR产物通过测序技术来鉴定微生物组成及其多样性.Mpbio公司的FastDNA Spin Kit for Soil试剂盒目前已广泛应用于DNA的提取,它是获得雪冰微生物高分子量、高纯度的基因组DNA非常重要的前提. ...
Sediment microbial community structure and mercury methylation in mercury-polluted Clear Lake, California
1
2000
... 早期对汞甲基化微生物的鉴定依赖于实验室培养,测试从厌氧沉积物中培养的分离物在添加无机汞后产生甲基汞的能力[64-65].然而,不同汞甲基化细菌的甲基化能力差异悬殊,如果忽视它是一个物种或菌种的特异性功能,使用这种方法预估环境中汞的甲基化潜力是欠准确的[43].目前主要通过16S rRNA基因的分析,评估微生物群落结构与环境中甲基汞和生物地球化学特性的关系[45,66-67].高通量测序作为一种新兴的免培养分子生物学技术,检测快速、准确、信息全面丰富,随着高通量测序技术的不断升级换代,测序通量、读长和准确度也有了很大提升,为认识各种环境中汞甲基化微生物多样性、群落结构组成及其生态功能提供了有利手段[68].Illumina公司的Solexa和Hiseq是目前全球使用量最大的第二代测序机器.现代分子生物学技术将从环境样品中提取的核酸(DNA或RNA)为分析对象,以进行目标片段的聚合酶链式反应(PCR)扩增,得到的PCR产物通过测序技术来鉴定微生物组成及其多样性.Mpbio公司的FastDNA Spin Kit for Soil试剂盒目前已广泛应用于DNA的提取,它是获得雪冰微生物高分子量、高纯度的基因组DNA非常重要的前提. ...
Application of high-throughput DNA sequencing in microbial ecology
1
2018
... 早期对汞甲基化微生物的鉴定依赖于实验室培养,测试从厌氧沉积物中培养的分离物在添加无机汞后产生甲基汞的能力[64-65].然而,不同汞甲基化细菌的甲基化能力差异悬殊,如果忽视它是一个物种或菌种的特异性功能,使用这种方法预估环境中汞的甲基化潜力是欠准确的[43].目前主要通过16S rRNA基因的分析,评估微生物群落结构与环境中甲基汞和生物地球化学特性的关系[45,66-67].高通量测序作为一种新兴的免培养分子生物学技术,检测快速、准确、信息全面丰富,随着高通量测序技术的不断升级换代,测序通量、读长和准确度也有了很大提升,为认识各种环境中汞甲基化微生物多样性、群落结构组成及其生态功能提供了有利手段[68].Illumina公司的Solexa和Hiseq是目前全球使用量最大的第二代测序机器.现代分子生物学技术将从环境样品中提取的核酸(DNA或RNA)为分析对象,以进行目标片段的聚合酶链式反应(PCR)扩增,得到的PCR产物通过测序技术来鉴定微生物组成及其多样性.Mpbio公司的FastDNA Spin Kit for Soil试剂盒目前已广泛应用于DNA的提取,它是获得雪冰微生物高分子量、高纯度的基因组DNA非常重要的前提. ...
高通量测序技术在微生物分子生态学研究中的应用
1
2018
... 早期对汞甲基化微生物的鉴定依赖于实验室培养,测试从厌氧沉积物中培养的分离物在添加无机汞后产生甲基汞的能力[64-65].然而,不同汞甲基化细菌的甲基化能力差异悬殊,如果忽视它是一个物种或菌种的特异性功能,使用这种方法预估环境中汞的甲基化潜力是欠准确的[43].目前主要通过16S rRNA基因的分析,评估微生物群落结构与环境中甲基汞和生物地球化学特性的关系[45,66-67].高通量测序作为一种新兴的免培养分子生物学技术,检测快速、准确、信息全面丰富,随着高通量测序技术的不断升级换代,测序通量、读长和准确度也有了很大提升,为认识各种环境中汞甲基化微生物多样性、群落结构组成及其生态功能提供了有利手段[68].Illumina公司的Solexa和Hiseq是目前全球使用量最大的第二代测序机器.现代分子生物学技术将从环境样品中提取的核酸(DNA或RNA)为分析对象,以进行目标片段的聚合酶链式反应(PCR)扩增,得到的PCR产物通过测序技术来鉴定微生物组成及其多样性.Mpbio公司的FastDNA Spin Kit for Soil试剂盒目前已广泛应用于DNA的提取,它是获得雪冰微生物高分子量、高纯度的基因组DNA非常重要的前提. ...
An improved hgcAB primer set and direct high-throughput sequencing expand Hg-methylator diversity in nature
1
2020
... 对hgcAB基因簇、mer基因簇等分子层面的研究极大地拓宽了我们对汞甲基化机制的认识[41,48].Gionfriddo等[69]开发出了一套全新的引物,并将汞甲基化细菌基因序列参考库扩展到了283个,从而可以更准确地获取汞甲基化微生物多样性和环境中甲基汞产生的潜力.当今采用高通量测序获取基因和微生物多样性的生物学信息,开辟了冰冻圈汞甲基化研究的新领域.冰冻圈汞甲基化微生物分析方法概括归纳如图2所示.分析冰冻圈中hgcAB基因的方法一种是基于聚合酶链式反应技术,利用hgcAB特异引物获得样品中的hgcAB基因序列和丰度;另一种是通过BLAST方法从样本宏基因组数据中筛选出hgcAB同源序列[34]. ...
Present century record of mercury species pollution in high altitude alpine snow and ice
1
2003
... 冰川作为冰冻圈重要组成因素,全球山地冰川(不包含格陵兰冰盖和南极冰盖)覆盖面积约为17×104 km2,共约11×104条冰川,主要分布在亚北极和青藏高原等地[10].早期Jitaru等[70]报道了欧洲阿尔卑斯山雪冰中溶解性甲基汞的含量(0.3~4.0 ng·L-1).近年来科研人员对我国青藏高原冰川也开展了研究工作,Huang等[19]对中国西部冰川上冰尘甲基汞含量进行测试分析,测得甲基汞平均含量为(1.0±0.4) ng·g-1,并初步估算出每年冰尘中的甲基汞总量约为(0.7±0.3) kg.微生物群落在汞的生物地球化学转化过程中非常活跃,冰川微生物活动可能发生汞微生物甲基化[71-74].大气汞沉降到青藏高原冰川之后在冰尘中积累,由于冰尘洞会发生季节性融化,这里便成为各种微生物群落的栖息地[75].前人研究指出我国西部冰川冰尘甲基汞与总汞的比值较高,说明冰尘表面可能是冰川汞甲基化的潜在位点[19].Zhang等[56]对青藏高原冰川开展了汞微生物甲基化初步研究,结合老虎沟12号冰川末端6个不同类型样品的汞形态和宏基因组学分析,发现在含大量碎屑和粉尘的尘埃层中汞浓度和汞甲基化基因(hgcA)丰度最高.不过该研究中汞甲基化细菌的未分类序列占比很高,微生物群落组成仍不明晰,尚未能全面地阐明汞甲基化的微生物学机制,需要进一步深入研究. ...
High microbial activity on glaciers: importance to the global carbon cycle
1
2009
... 冰川作为冰冻圈重要组成因素,全球山地冰川(不包含格陵兰冰盖和南极冰盖)覆盖面积约为17×104 km2,共约11×104条冰川,主要分布在亚北极和青藏高原等地[10].早期Jitaru等[70]报道了欧洲阿尔卑斯山雪冰中溶解性甲基汞的含量(0.3~4.0 ng·L-1).近年来科研人员对我国青藏高原冰川也开展了研究工作,Huang等[19]对中国西部冰川上冰尘甲基汞含量进行测试分析,测得甲基汞平均含量为(1.0±0.4) ng·g-1,并初步估算出每年冰尘中的甲基汞总量约为(0.7±0.3) kg.微生物群落在汞的生物地球化学转化过程中非常活跃,冰川微生物活动可能发生汞微生物甲基化[71-74].大气汞沉降到青藏高原冰川之后在冰尘中积累,由于冰尘洞会发生季节性融化,这里便成为各种微生物群落的栖息地[75].前人研究指出我国西部冰川冰尘甲基汞与总汞的比值较高,说明冰尘表面可能是冰川汞甲基化的潜在位点[19].Zhang等[56]对青藏高原冰川开展了汞微生物甲基化初步研究,结合老虎沟12号冰川末端6个不同类型样品的汞形态和宏基因组学分析,发现在含大量碎屑和粉尘的尘埃层中汞浓度和汞甲基化基因(hgcA)丰度最高.不过该研究中汞甲基化细菌的未分类序列占比很高,微生物群落组成仍不明晰,尚未能全面地阐明汞甲基化的微生物学机制,需要进一步深入研究. ...
The microbiome of glaciers and ice sheets
0
2017
Microbial ecology of the cryosphere: sea ice and glacial habitats
0
2015
Glacial ecosystems
1
2008
... 冰川作为冰冻圈重要组成因素,全球山地冰川(不包含格陵兰冰盖和南极冰盖)覆盖面积约为17×104 km2,共约11×104条冰川,主要分布在亚北极和青藏高原等地[10].早期Jitaru等[70]报道了欧洲阿尔卑斯山雪冰中溶解性甲基汞的含量(0.3~4.0 ng·L-1).近年来科研人员对我国青藏高原冰川也开展了研究工作,Huang等[19]对中国西部冰川上冰尘甲基汞含量进行测试分析,测得甲基汞平均含量为(1.0±0.4) ng·g-1,并初步估算出每年冰尘中的甲基汞总量约为(0.7±0.3) kg.微生物群落在汞的生物地球化学转化过程中非常活跃,冰川微生物活动可能发生汞微生物甲基化[71-74].大气汞沉降到青藏高原冰川之后在冰尘中积累,由于冰尘洞会发生季节性融化,这里便成为各种微生物群落的栖息地[75].前人研究指出我国西部冰川冰尘甲基汞与总汞的比值较高,说明冰尘表面可能是冰川汞甲基化的潜在位点[19].Zhang等[56]对青藏高原冰川开展了汞微生物甲基化初步研究,结合老虎沟12号冰川末端6个不同类型样品的汞形态和宏基因组学分析,发现在含大量碎屑和粉尘的尘埃层中汞浓度和汞甲基化基因(hgcA)丰度最高.不过该研究中汞甲基化细菌的未分类序列占比很高,微生物群落组成仍不明晰,尚未能全面地阐明汞甲基化的微生物学机制,需要进一步深入研究. ...
Cryoconite: the dark biological secret of the cryosphere
1
2016
... 冰川作为冰冻圈重要组成因素,全球山地冰川(不包含格陵兰冰盖和南极冰盖)覆盖面积约为17×104 km2,共约11×104条冰川,主要分布在亚北极和青藏高原等地[10].早期Jitaru等[70]报道了欧洲阿尔卑斯山雪冰中溶解性甲基汞的含量(0.3~4.0 ng·L-1).近年来科研人员对我国青藏高原冰川也开展了研究工作,Huang等[19]对中国西部冰川上冰尘甲基汞含量进行测试分析,测得甲基汞平均含量为(1.0±0.4) ng·g-1,并初步估算出每年冰尘中的甲基汞总量约为(0.7±0.3) kg.微生物群落在汞的生物地球化学转化过程中非常活跃,冰川微生物活动可能发生汞微生物甲基化[71-74].大气汞沉降到青藏高原冰川之后在冰尘中积累,由于冰尘洞会发生季节性融化,这里便成为各种微生物群落的栖息地[75].前人研究指出我国西部冰川冰尘甲基汞与总汞的比值较高,说明冰尘表面可能是冰川汞甲基化的潜在位点[19].Zhang等[56]对青藏高原冰川开展了汞微生物甲基化初步研究,结合老虎沟12号冰川末端6个不同类型样品的汞形态和宏基因组学分析,发现在含大量碎屑和粉尘的尘埃层中汞浓度和汞甲基化基因(hgcA)丰度最高.不过该研究中汞甲基化细菌的未分类序列占比很高,微生物群落组成仍不明晰,尚未能全面地阐明汞甲基化的微生物学机制,需要进一步深入研究. ...
The fate of mercury species in a sub-arctic snowpack during snowmelt
1
2003
... 冰川对气候变化响应非常敏感,由于气候变暖,冰川正在以前所未有的速度退缩[9].冰川消融不仅给下游生态系统带来水资源,而且还可能释放出在冰川中存储了几个世纪的汞[20].在消融初期,雪冰中的汞以脉冲式释放到冰川径流,近90%的汞会被释放进入融水[76].Sun等[77-78]估算出喜马拉雅地区融水径流汞输出通量为337 kg·a-1,青藏高原冬克玛底流域中汞输出总量约为747.4 g·a-1,亚北极Yukon流域汞输出总量约为4 400 kg·a-1[79-80].Zolkos等[81]对六条北极主要河流进行全年采样调查,估算出总汞输出通量为20 Mg·a-1,Dastoor等[18]估算了北极冰川补给流域总汞年产率为(0.03~3.9 g·km2·a-1).尽管青藏高原河流输出总量较低,但山地冰川(如扎当冰川等)的汞输出通量要高于大部分河流,表明青藏高原山地冰川补给流域具有更高的汞释放效率[30-31,78].在未来全球升温1.5 ℃情景下,到21世纪末青藏高原及周边地区冰川储量将减少2 169~3 217 km3,预估计在未来约11~16吨汞污染物将从冰川释放到下游地区(冰川中汞平均浓度为5.6 ng·L-1)[82-84].富含湿地的冰川流域通常有着更高的总汞和甲基汞含量,生物群落中的汞含量亦较高[85].阿拉斯加东南部冰河湾下游集水区,存在湿地的溪流溶解汞的比例大得多(0.1~3.4 ng·L-1),而裸露的冰川径流溶解汞含量仅为0.1~0.9 ng·L-1[85].湿地可以提供丰富的溶解性有机物,同时这种厌氧环境还有利于使汞甲基化的硫酸盐还原菌生存,可以增强微生物的活性和汞甲基化[85-87].汞污染物进入湿地之后被汞甲基化微生物转化为毒性更强的甲基汞再次进入径流,对下游地区人类健康和生态系统存在潜在威胁[88-91].因此,冰川流域生物群落中的甲基汞浓度与流域湿地覆盖率之间存在密切关系,冰川流域湿地可能成为冰川径流中甲基汞净来源,并将其输送到下游生态系统[85,88,92-95]. ...
Melting Himalayas and mercury export: results of continuous observations from the Rongbuk Glacier on Mt. Everest and future insights
1
2022
... 冰川对气候变化响应非常敏感,由于气候变暖,冰川正在以前所未有的速度退缩[9].冰川消融不仅给下游生态系统带来水资源,而且还可能释放出在冰川中存储了几个世纪的汞[20].在消融初期,雪冰中的汞以脉冲式释放到冰川径流,近90%的汞会被释放进入融水[76].Sun等[77-78]估算出喜马拉雅地区融水径流汞输出通量为337 kg·a-1,青藏高原冬克玛底流域中汞输出总量约为747.4 g·a-1,亚北极Yukon流域汞输出总量约为4 400 kg·a-1[79-80].Zolkos等[81]对六条北极主要河流进行全年采样调查,估算出总汞输出通量为20 Mg·a-1,Dastoor等[18]估算了北极冰川补给流域总汞年产率为(0.03~3.9 g·km2·a-1).尽管青藏高原河流输出总量较低,但山地冰川(如扎当冰川等)的汞输出通量要高于大部分河流,表明青藏高原山地冰川补给流域具有更高的汞释放效率[30-31,78].在未来全球升温1.5 ℃情景下,到21世纪末青藏高原及周边地区冰川储量将减少2 169~3 217 km3,预估计在未来约11~16吨汞污染物将从冰川释放到下游地区(冰川中汞平均浓度为5.6 ng·L-1)[82-84].富含湿地的冰川流域通常有着更高的总汞和甲基汞含量,生物群落中的汞含量亦较高[85].阿拉斯加东南部冰河湾下游集水区,存在湿地的溪流溶解汞的比例大得多(0.1~3.4 ng·L-1),而裸露的冰川径流溶解汞含量仅为0.1~0.9 ng·L-1[85].湿地可以提供丰富的溶解性有机物,同时这种厌氧环境还有利于使汞甲基化的硫酸盐还原菌生存,可以增强微生物的活性和汞甲基化[85-87].汞污染物进入湿地之后被汞甲基化微生物转化为毒性更强的甲基汞再次进入径流,对下游地区人类健康和生态系统存在潜在威胁[88-91].因此,冰川流域生物群落中的甲基汞浓度与流域湿地覆盖率之间存在密切关系,冰川流域湿地可能成为冰川径流中甲基汞净来源,并将其输送到下游生态系统[85,88,92-95]. ...
Behavior and environmental effects of mercury relevant to the melt of alpine glacier: a review
2
2017
... 冰川对气候变化响应非常敏感,由于气候变暖,冰川正在以前所未有的速度退缩[9].冰川消融不仅给下游生态系统带来水资源,而且还可能释放出在冰川中存储了几个世纪的汞[20].在消融初期,雪冰中的汞以脉冲式释放到冰川径流,近90%的汞会被释放进入融水[76].Sun等[77-78]估算出喜马拉雅地区融水径流汞输出通量为337 kg·a-1,青藏高原冬克玛底流域中汞输出总量约为747.4 g·a-1,亚北极Yukon流域汞输出总量约为4 400 kg·a-1[79-80].Zolkos等[81]对六条北极主要河流进行全年采样调查,估算出总汞输出通量为20 Mg·a-1,Dastoor等[18]估算了北极冰川补给流域总汞年产率为(0.03~3.9 g·km2·a-1).尽管青藏高原河流输出总量较低,但山地冰川(如扎当冰川等)的汞输出通量要高于大部分河流,表明青藏高原山地冰川补给流域具有更高的汞释放效率[30-31,78].在未来全球升温1.5 ℃情景下,到21世纪末青藏高原及周边地区冰川储量将减少2 169~3 217 km3,预估计在未来约11~16吨汞污染物将从冰川释放到下游地区(冰川中汞平均浓度为5.6 ng·L-1)[82-84].富含湿地的冰川流域通常有着更高的总汞和甲基汞含量,生物群落中的汞含量亦较高[85].阿拉斯加东南部冰河湾下游集水区,存在湿地的溪流溶解汞的比例大得多(0.1~3.4 ng·L-1),而裸露的冰川径流溶解汞含量仅为0.1~0.9 ng·L-1[85].湿地可以提供丰富的溶解性有机物,同时这种厌氧环境还有利于使汞甲基化的硫酸盐还原菌生存,可以增强微生物的活性和汞甲基化[85-87].汞污染物进入湿地之后被汞甲基化微生物转化为毒性更强的甲基汞再次进入径流,对下游地区人类健康和生态系统存在潜在威胁[88-91].因此,冰川流域生物群落中的甲基汞浓度与流域湿地覆盖率之间存在密切关系,冰川流域湿地可能成为冰川径流中甲基汞净来源,并将其输送到下游生态系统[85,88,92-95]. ...
... ,78].在未来全球升温1.5 ℃情景下,到21世纪末青藏高原及周边地区冰川储量将减少2 169~3 217 km3,预估计在未来约11~16吨汞污染物将从冰川释放到下游地区(冰川中汞平均浓度为5.6 ng·L-1)[82-84].富含湿地的冰川流域通常有着更高的总汞和甲基汞含量,生物群落中的汞含量亦较高[85].阿拉斯加东南部冰河湾下游集水区,存在湿地的溪流溶解汞的比例大得多(0.1~3.4 ng·L-1),而裸露的冰川径流溶解汞含量仅为0.1~0.9 ng·L-1[85].湿地可以提供丰富的溶解性有机物,同时这种厌氧环境还有利于使汞甲基化的硫酸盐还原菌生存,可以增强微生物的活性和汞甲基化[85-87].汞污染物进入湿地之后被汞甲基化微生物转化为毒性更强的甲基汞再次进入径流,对下游地区人类健康和生态系统存在潜在威胁[88-91].因此,冰川流域生物群落中的甲基汞浓度与流域湿地覆盖率之间存在密切关系,冰川流域湿地可能成为冰川径流中甲基汞净来源,并将其输送到下游生态系统[85,88,92-95]. ...
山地冰川消融过程中汞的行为及环境效应综述
2
2017
... 冰川对气候变化响应非常敏感,由于气候变暖,冰川正在以前所未有的速度退缩[9].冰川消融不仅给下游生态系统带来水资源,而且还可能释放出在冰川中存储了几个世纪的汞[20].在消融初期,雪冰中的汞以脉冲式释放到冰川径流,近90%的汞会被释放进入融水[76].Sun等[77-78]估算出喜马拉雅地区融水径流汞输出通量为337 kg·a-1,青藏高原冬克玛底流域中汞输出总量约为747.4 g·a-1,亚北极Yukon流域汞输出总量约为4 400 kg·a-1[79-80].Zolkos等[81]对六条北极主要河流进行全年采样调查,估算出总汞输出通量为20 Mg·a-1,Dastoor等[18]估算了北极冰川补给流域总汞年产率为(0.03~3.9 g·km2·a-1).尽管青藏高原河流输出总量较低,但山地冰川(如扎当冰川等)的汞输出通量要高于大部分河流,表明青藏高原山地冰川补给流域具有更高的汞释放效率[30-31,78].在未来全球升温1.5 ℃情景下,到21世纪末青藏高原及周边地区冰川储量将减少2 169~3 217 km3,预估计在未来约11~16吨汞污染物将从冰川释放到下游地区(冰川中汞平均浓度为5.6 ng·L-1)[82-84].富含湿地的冰川流域通常有着更高的总汞和甲基汞含量,生物群落中的汞含量亦较高[85].阿拉斯加东南部冰河湾下游集水区,存在湿地的溪流溶解汞的比例大得多(0.1~3.4 ng·L-1),而裸露的冰川径流溶解汞含量仅为0.1~0.9 ng·L-1[85].湿地可以提供丰富的溶解性有机物,同时这种厌氧环境还有利于使汞甲基化的硫酸盐还原菌生存,可以增强微生物的活性和汞甲基化[85-87].汞污染物进入湿地之后被汞甲基化微生物转化为毒性更强的甲基汞再次进入径流,对下游地区人类健康和生态系统存在潜在威胁[88-91].因此,冰川流域生物群落中的甲基汞浓度与流域湿地覆盖率之间存在密切关系,冰川流域湿地可能成为冰川径流中甲基汞净来源,并将其输送到下游生态系统[85,88,92-95]. ...
... ,78].在未来全球升温1.5 ℃情景下,到21世纪末青藏高原及周边地区冰川储量将减少2 169~3 217 km3,预估计在未来约11~16吨汞污染物将从冰川释放到下游地区(冰川中汞平均浓度为5.6 ng·L-1)[82-84].富含湿地的冰川流域通常有着更高的总汞和甲基汞含量,生物群落中的汞含量亦较高[85].阿拉斯加东南部冰河湾下游集水区,存在湿地的溪流溶解汞的比例大得多(0.1~3.4 ng·L-1),而裸露的冰川径流溶解汞含量仅为0.1~0.9 ng·L-1[85].湿地可以提供丰富的溶解性有机物,同时这种厌氧环境还有利于使汞甲基化的硫酸盐还原菌生存,可以增强微生物的活性和汞甲基化[85-87].汞污染物进入湿地之后被汞甲基化微生物转化为毒性更强的甲基汞再次进入径流,对下游地区人类健康和生态系统存在潜在威胁[88-91].因此,冰川流域生物群落中的甲基汞浓度与流域湿地覆盖率之间存在密切关系,冰川流域湿地可能成为冰川径流中甲基汞净来源,并将其输送到下游生态系统[85,88,92-95]. ...
Insights into mercury deposition and spatiotemporal variation in the glacier and melt water from the central Tibetan Plateau
1
2017
... 冰川对气候变化响应非常敏感,由于气候变暖,冰川正在以前所未有的速度退缩[9].冰川消融不仅给下游生态系统带来水资源,而且还可能释放出在冰川中存储了几个世纪的汞[20].在消融初期,雪冰中的汞以脉冲式释放到冰川径流,近90%的汞会被释放进入融水[76].Sun等[77-78]估算出喜马拉雅地区融水径流汞输出通量为337 kg·a-1,青藏高原冬克玛底流域中汞输出总量约为747.4 g·a-1,亚北极Yukon流域汞输出总量约为4 400 kg·a-1[79-80].Zolkos等[81]对六条北极主要河流进行全年采样调查,估算出总汞输出通量为20 Mg·a-1,Dastoor等[18]估算了北极冰川补给流域总汞年产率为(0.03~3.9 g·km2·a-1).尽管青藏高原河流输出总量较低,但山地冰川(如扎当冰川等)的汞输出通量要高于大部分河流,表明青藏高原山地冰川补给流域具有更高的汞释放效率[30-31,78].在未来全球升温1.5 ℃情景下,到21世纪末青藏高原及周边地区冰川储量将减少2 169~3 217 km3,预估计在未来约11~16吨汞污染物将从冰川释放到下游地区(冰川中汞平均浓度为5.6 ng·L-1)[82-84].富含湿地的冰川流域通常有着更高的总汞和甲基汞含量,生物群落中的汞含量亦较高[85].阿拉斯加东南部冰河湾下游集水区,存在湿地的溪流溶解汞的比例大得多(0.1~3.4 ng·L-1),而裸露的冰川径流溶解汞含量仅为0.1~0.9 ng·L-1[85].湿地可以提供丰富的溶解性有机物,同时这种厌氧环境还有利于使汞甲基化的硫酸盐还原菌生存,可以增强微生物的活性和汞甲基化[85-87].汞污染物进入湿地之后被汞甲基化微生物转化为毒性更强的甲基汞再次进入径流,对下游地区人类健康和生态系统存在潜在威胁[88-91].因此,冰川流域生物群落中的甲基汞浓度与流域湿地覆盖率之间存在密切关系,冰川流域湿地可能成为冰川径流中甲基汞净来源,并将其输送到下游生态系统[85,88,92-95]. ...
Mercury export from the Yukon River Basin and potential response to a changing climate
1
2011
... 冰川对气候变化响应非常敏感,由于气候变暖,冰川正在以前所未有的速度退缩[9].冰川消融不仅给下游生态系统带来水资源,而且还可能释放出在冰川中存储了几个世纪的汞[20].在消融初期,雪冰中的汞以脉冲式释放到冰川径流,近90%的汞会被释放进入融水[76].Sun等[77-78]估算出喜马拉雅地区融水径流汞输出通量为337 kg·a-1,青藏高原冬克玛底流域中汞输出总量约为747.4 g·a-1,亚北极Yukon流域汞输出总量约为4 400 kg·a-1[79-80].Zolkos等[81]对六条北极主要河流进行全年采样调查,估算出总汞输出通量为20 Mg·a-1,Dastoor等[18]估算了北极冰川补给流域总汞年产率为(0.03~3.9 g·km2·a-1).尽管青藏高原河流输出总量较低,但山地冰川(如扎当冰川等)的汞输出通量要高于大部分河流,表明青藏高原山地冰川补给流域具有更高的汞释放效率[30-31,78].在未来全球升温1.5 ℃情景下,到21世纪末青藏高原及周边地区冰川储量将减少2 169~3 217 km3,预估计在未来约11~16吨汞污染物将从冰川释放到下游地区(冰川中汞平均浓度为5.6 ng·L-1)[82-84].富含湿地的冰川流域通常有着更高的总汞和甲基汞含量,生物群落中的汞含量亦较高[85].阿拉斯加东南部冰河湾下游集水区,存在湿地的溪流溶解汞的比例大得多(0.1~3.4 ng·L-1),而裸露的冰川径流溶解汞含量仅为0.1~0.9 ng·L-1[85].湿地可以提供丰富的溶解性有机物,同时这种厌氧环境还有利于使汞甲基化的硫酸盐还原菌生存,可以增强微生物的活性和汞甲基化[85-87].汞污染物进入湿地之后被汞甲基化微生物转化为毒性更强的甲基汞再次进入径流,对下游地区人类健康和生态系统存在潜在威胁[88-91].因此,冰川流域生物群落中的甲基汞浓度与流域湿地覆盖率之间存在密切关系,冰川流域湿地可能成为冰川径流中甲基汞净来源,并将其输送到下游生态系统[85,88,92-95]. ...
Mercury export from Arctic great rivers
1
2020
... 冰川对气候变化响应非常敏感,由于气候变暖,冰川正在以前所未有的速度退缩[9].冰川消融不仅给下游生态系统带来水资源,而且还可能释放出在冰川中存储了几个世纪的汞[20].在消融初期,雪冰中的汞以脉冲式释放到冰川径流,近90%的汞会被释放进入融水[76].Sun等[77-78]估算出喜马拉雅地区融水径流汞输出通量为337 kg·a-1,青藏高原冬克玛底流域中汞输出总量约为747.4 g·a-1,亚北极Yukon流域汞输出总量约为4 400 kg·a-1[79-80].Zolkos等[81]对六条北极主要河流进行全年采样调查,估算出总汞输出通量为20 Mg·a-1,Dastoor等[18]估算了北极冰川补给流域总汞年产率为(0.03~3.9 g·km2·a-1).尽管青藏高原河流输出总量较低,但山地冰川(如扎当冰川等)的汞输出通量要高于大部分河流,表明青藏高原山地冰川补给流域具有更高的汞释放效率[30-31,78].在未来全球升温1.5 ℃情景下,到21世纪末青藏高原及周边地区冰川储量将减少2 169~3 217 km3,预估计在未来约11~16吨汞污染物将从冰川释放到下游地区(冰川中汞平均浓度为5.6 ng·L-1)[82-84].富含湿地的冰川流域通常有着更高的总汞和甲基汞含量,生物群落中的汞含量亦较高[85].阿拉斯加东南部冰河湾下游集水区,存在湿地的溪流溶解汞的比例大得多(0.1~3.4 ng·L-1),而裸露的冰川径流溶解汞含量仅为0.1~0.9 ng·L-1[85].湿地可以提供丰富的溶解性有机物,同时这种厌氧环境还有利于使汞甲基化的硫酸盐还原菌生存,可以增强微生物的活性和汞甲基化[85-87].汞污染物进入湿地之后被汞甲基化微生物转化为毒性更强的甲基汞再次进入径流,对下游地区人类健康和生态系统存在潜在威胁[88-91].因此,冰川流域生物群落中的甲基汞浓度与流域湿地覆盖率之间存在密切关系,冰川流域湿地可能成为冰川径流中甲基汞净来源,并将其输送到下游生态系统[85,88,92-95]. ...
Spatiotemporal pattern, trend, and influence of glacier change in Tibetan Plateau and surroundings under global warming
1
2019
... 冰川对气候变化响应非常敏感,由于气候变暖,冰川正在以前所未有的速度退缩[9].冰川消融不仅给下游生态系统带来水资源,而且还可能释放出在冰川中存储了几个世纪的汞[20].在消融初期,雪冰中的汞以脉冲式释放到冰川径流,近90%的汞会被释放进入融水[76].Sun等[77-78]估算出喜马拉雅地区融水径流汞输出通量为337 kg·a-1,青藏高原冬克玛底流域中汞输出总量约为747.4 g·a-1,亚北极Yukon流域汞输出总量约为4 400 kg·a-1[79-80].Zolkos等[81]对六条北极主要河流进行全年采样调查,估算出总汞输出通量为20 Mg·a-1,Dastoor等[18]估算了北极冰川补给流域总汞年产率为(0.03~3.9 g·km2·a-1).尽管青藏高原河流输出总量较低,但山地冰川(如扎当冰川等)的汞输出通量要高于大部分河流,表明青藏高原山地冰川补给流域具有更高的汞释放效率[30-31,78].在未来全球升温1.5 ℃情景下,到21世纪末青藏高原及周边地区冰川储量将减少2 169~3 217 km3,预估计在未来约11~16吨汞污染物将从冰川释放到下游地区(冰川中汞平均浓度为5.6 ng·L-1)[82-84].富含湿地的冰川流域通常有着更高的总汞和甲基汞含量,生物群落中的汞含量亦较高[85].阿拉斯加东南部冰河湾下游集水区,存在湿地的溪流溶解汞的比例大得多(0.1~3.4 ng·L-1),而裸露的冰川径流溶解汞含量仅为0.1~0.9 ng·L-1[85].湿地可以提供丰富的溶解性有机物,同时这种厌氧环境还有利于使汞甲基化的硫酸盐还原菌生存,可以增强微生物的活性和汞甲基化[85-87].汞污染物进入湿地之后被汞甲基化微生物转化为毒性更强的甲基汞再次进入径流,对下游地区人类健康和生态系统存在潜在威胁[88-91].因此,冰川流域生物群落中的甲基汞浓度与流域湿地覆盖率之间存在密切关系,冰川流域湿地可能成为冰川径流中甲基汞净来源,并将其输送到下游生态系统[85,88,92-95]. ...
全球变暖背景下青藏高原及周边地区冰川变化的时空格局与趋势及影响
1
2019
... 冰川对气候变化响应非常敏感,由于气候变暖,冰川正在以前所未有的速度退缩[9].冰川消融不仅给下游生态系统带来水资源,而且还可能释放出在冰川中存储了几个世纪的汞[20].在消融初期,雪冰中的汞以脉冲式释放到冰川径流,近90%的汞会被释放进入融水[76].Sun等[77-78]估算出喜马拉雅地区融水径流汞输出通量为337 kg·a-1,青藏高原冬克玛底流域中汞输出总量约为747.4 g·a-1,亚北极Yukon流域汞输出总量约为4 400 kg·a-1[79-80].Zolkos等[81]对六条北极主要河流进行全年采样调查,估算出总汞输出通量为20 Mg·a-1,Dastoor等[18]估算了北极冰川补给流域总汞年产率为(0.03~3.9 g·km2·a-1).尽管青藏高原河流输出总量较低,但山地冰川(如扎当冰川等)的汞输出通量要高于大部分河流,表明青藏高原山地冰川补给流域具有更高的汞释放效率[30-31,78].在未来全球升温1.5 ℃情景下,到21世纪末青藏高原及周边地区冰川储量将减少2 169~3 217 km3,预估计在未来约11~16吨汞污染物将从冰川释放到下游地区(冰川中汞平均浓度为5.6 ng·L-1)[82-84].富含湿地的冰川流域通常有着更高的总汞和甲基汞含量,生物群落中的汞含量亦较高[85].阿拉斯加东南部冰河湾下游集水区,存在湿地的溪流溶解汞的比例大得多(0.1~3.4 ng·L-1),而裸露的冰川径流溶解汞含量仅为0.1~0.9 ng·L-1[85].湿地可以提供丰富的溶解性有机物,同时这种厌氧环境还有利于使汞甲基化的硫酸盐还原菌生存,可以增强微生物的活性和汞甲基化[85-87].汞污染物进入湿地之后被汞甲基化微生物转化为毒性更强的甲基汞再次进入径流,对下游地区人类健康和生态系统存在潜在威胁[88-91].因此,冰川流域生物群落中的甲基汞浓度与流域湿地覆盖率之间存在密切关系,冰川流域湿地可能成为冰川径流中甲基汞净来源,并将其输送到下游生态系统[85,88,92-95]. ...
RGI Consortium Randolph Glacier Inventory (v
0
6
Global glacier mass changes and their contributions to sea-level rise from 1961 to 2016
1
2019
... 冰川对气候变化响应非常敏感,由于气候变暖,冰川正在以前所未有的速度退缩[9].冰川消融不仅给下游生态系统带来水资源,而且还可能释放出在冰川中存储了几个世纪的汞[20].在消融初期,雪冰中的汞以脉冲式释放到冰川径流,近90%的汞会被释放进入融水[76].Sun等[77-78]估算出喜马拉雅地区融水径流汞输出通量为337 kg·a-1,青藏高原冬克玛底流域中汞输出总量约为747.4 g·a-1,亚北极Yukon流域汞输出总量约为4 400 kg·a-1[79-80].Zolkos等[81]对六条北极主要河流进行全年采样调查,估算出总汞输出通量为20 Mg·a-1,Dastoor等[18]估算了北极冰川补给流域总汞年产率为(0.03~3.9 g·km2·a-1).尽管青藏高原河流输出总量较低,但山地冰川(如扎当冰川等)的汞输出通量要高于大部分河流,表明青藏高原山地冰川补给流域具有更高的汞释放效率[30-31,78].在未来全球升温1.5 ℃情景下,到21世纪末青藏高原及周边地区冰川储量将减少2 169~3 217 km3,预估计在未来约11~16吨汞污染物将从冰川释放到下游地区(冰川中汞平均浓度为5.6 ng·L-1)[82-84].富含湿地的冰川流域通常有着更高的总汞和甲基汞含量,生物群落中的汞含量亦较高[85].阿拉斯加东南部冰河湾下游集水区,存在湿地的溪流溶解汞的比例大得多(0.1~3.4 ng·L-1),而裸露的冰川径流溶解汞含量仅为0.1~0.9 ng·L-1[85].湿地可以提供丰富的溶解性有机物,同时这种厌氧环境还有利于使汞甲基化的硫酸盐还原菌生存,可以增强微生物的活性和汞甲基化[85-87].汞污染物进入湿地之后被汞甲基化微生物转化为毒性更强的甲基汞再次进入径流,对下游地区人类健康和生态系统存在潜在威胁[88-91].因此,冰川流域生物群落中的甲基汞浓度与流域湿地覆盖率之间存在密切关系,冰川流域湿地可能成为冰川径流中甲基汞净来源,并将其输送到下游生态系统[85,88,92-95]. ...
Spatial distribution of mercury in southeastern Alaskan streams influenced by glaciers, wetlands, and salmon
5
2014
... 冰川对气候变化响应非常敏感,由于气候变暖,冰川正在以前所未有的速度退缩[9].冰川消融不仅给下游生态系统带来水资源,而且还可能释放出在冰川中存储了几个世纪的汞[20].在消融初期,雪冰中的汞以脉冲式释放到冰川径流,近90%的汞会被释放进入融水[76].Sun等[77-78]估算出喜马拉雅地区融水径流汞输出通量为337 kg·a-1,青藏高原冬克玛底流域中汞输出总量约为747.4 g·a-1,亚北极Yukon流域汞输出总量约为4 400 kg·a-1[79-80].Zolkos等[81]对六条北极主要河流进行全年采样调查,估算出总汞输出通量为20 Mg·a-1,Dastoor等[18]估算了北极冰川补给流域总汞年产率为(0.03~3.9 g·km2·a-1).尽管青藏高原河流输出总量较低,但山地冰川(如扎当冰川等)的汞输出通量要高于大部分河流,表明青藏高原山地冰川补给流域具有更高的汞释放效率[30-31,78].在未来全球升温1.5 ℃情景下,到21世纪末青藏高原及周边地区冰川储量将减少2 169~3 217 km3,预估计在未来约11~16吨汞污染物将从冰川释放到下游地区(冰川中汞平均浓度为5.6 ng·L-1)[82-84].富含湿地的冰川流域通常有着更高的总汞和甲基汞含量,生物群落中的汞含量亦较高[85].阿拉斯加东南部冰河湾下游集水区,存在湿地的溪流溶解汞的比例大得多(0.1~3.4 ng·L-1),而裸露的冰川径流溶解汞含量仅为0.1~0.9 ng·L-1[85].湿地可以提供丰富的溶解性有机物,同时这种厌氧环境还有利于使汞甲基化的硫酸盐还原菌生存,可以增强微生物的活性和汞甲基化[85-87].汞污染物进入湿地之后被汞甲基化微生物转化为毒性更强的甲基汞再次进入径流,对下游地区人类健康和生态系统存在潜在威胁[88-91].因此,冰川流域生物群落中的甲基汞浓度与流域湿地覆盖率之间存在密切关系,冰川流域湿地可能成为冰川径流中甲基汞净来源,并将其输送到下游生态系统[85,88,92-95]. ...
... -1[85].湿地可以提供丰富的溶解性有机物,同时这种厌氧环境还有利于使汞甲基化的硫酸盐还原菌生存,可以增强微生物的活性和汞甲基化[85-87].汞污染物进入湿地之后被汞甲基化微生物转化为毒性更强的甲基汞再次进入径流,对下游地区人类健康和生态系统存在潜在威胁[88-91].因此,冰川流域生物群落中的甲基汞浓度与流域湿地覆盖率之间存在密切关系,冰川流域湿地可能成为冰川径流中甲基汞净来源,并将其输送到下游生态系统[85,88,92-95]. ...
... [85-87].汞污染物进入湿地之后被汞甲基化微生物转化为毒性更强的甲基汞再次进入径流,对下游地区人类健康和生态系统存在潜在威胁[88-91].因此,冰川流域生物群落中的甲基汞浓度与流域湿地覆盖率之间存在密切关系,冰川流域湿地可能成为冰川径流中甲基汞净来源,并将其输送到下游生态系统[85,88,92-95]. ...
... [85,88,92-95]. ...
... 大量研究表明偏远冰冻圈下游地区水生生态系统甲基汞的富集效应是非常显著的[85,96-97].格陵兰冰盖下游湾峡水生生态系统中,融水径流汞浓度(44~348 ng·L-1)比北极其他河流(0.3~2.8 ng·L-1)高出1~2个数量级,甲基汞浓度(1.0~3.5 ng·L-1)甚至高于大多数淡水,与湿地水平相当[20].我国学者亦研究发现青藏高原河流和湖泊中鱼类甲基汞含量(24.9~1 196 ng·g-1)为中国野生鱼类汞浓度的最高水平[96].此外,前人在冰川补给湖观测到甲基汞的峰值出现在化学跃层,甲基汞占总汞的比例高达57%,同时16S rRNA数据显示检测出硫酸盐还原菌,因此受冰川融水补给的冰川湖化学跃层存在着较强的汞甲基化潜力[97-98]. ...
Redefining the role of wetlands as methyl mercury sources insight from wetlands before and after restoration (Doctoral Thesis)
0
2012
Dissolved organic matter enhances microbial mercury methylation under sulfidic conditions
1
2012
... 冰川对气候变化响应非常敏感,由于气候变暖,冰川正在以前所未有的速度退缩[9].冰川消融不仅给下游生态系统带来水资源,而且还可能释放出在冰川中存储了几个世纪的汞[20].在消融初期,雪冰中的汞以脉冲式释放到冰川径流,近90%的汞会被释放进入融水[76].Sun等[77-78]估算出喜马拉雅地区融水径流汞输出通量为337 kg·a-1,青藏高原冬克玛底流域中汞输出总量约为747.4 g·a-1,亚北极Yukon流域汞输出总量约为4 400 kg·a-1[79-80].Zolkos等[81]对六条北极主要河流进行全年采样调查,估算出总汞输出通量为20 Mg·a-1,Dastoor等[18]估算了北极冰川补给流域总汞年产率为(0.03~3.9 g·km2·a-1).尽管青藏高原河流输出总量较低,但山地冰川(如扎当冰川等)的汞输出通量要高于大部分河流,表明青藏高原山地冰川补给流域具有更高的汞释放效率[30-31,78].在未来全球升温1.5 ℃情景下,到21世纪末青藏高原及周边地区冰川储量将减少2 169~3 217 km3,预估计在未来约11~16吨汞污染物将从冰川释放到下游地区(冰川中汞平均浓度为5.6 ng·L-1)[82-84].富含湿地的冰川流域通常有着更高的总汞和甲基汞含量,生物群落中的汞含量亦较高[85].阿拉斯加东南部冰河湾下游集水区,存在湿地的溪流溶解汞的比例大得多(0.1~3.4 ng·L-1),而裸露的冰川径流溶解汞含量仅为0.1~0.9 ng·L-1[85].湿地可以提供丰富的溶解性有机物,同时这种厌氧环境还有利于使汞甲基化的硫酸盐还原菌生存,可以增强微生物的活性和汞甲基化[85-87].汞污染物进入湿地之后被汞甲基化微生物转化为毒性更强的甲基汞再次进入径流,对下游地区人类健康和生态系统存在潜在威胁[88-91].因此,冰川流域生物群落中的甲基汞浓度与流域湿地覆盖率之间存在密切关系,冰川流域湿地可能成为冰川径流中甲基汞净来源,并将其输送到下游生态系统[85,88,92-95]. ...
Mercury speciation and distribution in a glacierized mountain environment and their relevance to environmental risks in the inland Tibetan Plateau
2
2018
... 冰川对气候变化响应非常敏感,由于气候变暖,冰川正在以前所未有的速度退缩[9].冰川消融不仅给下游生态系统带来水资源,而且还可能释放出在冰川中存储了几个世纪的汞[20].在消融初期,雪冰中的汞以脉冲式释放到冰川径流,近90%的汞会被释放进入融水[76].Sun等[77-78]估算出喜马拉雅地区融水径流汞输出通量为337 kg·a-1,青藏高原冬克玛底流域中汞输出总量约为747.4 g·a-1,亚北极Yukon流域汞输出总量约为4 400 kg·a-1[79-80].Zolkos等[81]对六条北极主要河流进行全年采样调查,估算出总汞输出通量为20 Mg·a-1,Dastoor等[18]估算了北极冰川补给流域总汞年产率为(0.03~3.9 g·km2·a-1).尽管青藏高原河流输出总量较低,但山地冰川(如扎当冰川等)的汞输出通量要高于大部分河流,表明青藏高原山地冰川补给流域具有更高的汞释放效率[30-31,78].在未来全球升温1.5 ℃情景下,到21世纪末青藏高原及周边地区冰川储量将减少2 169~3 217 km3,预估计在未来约11~16吨汞污染物将从冰川释放到下游地区(冰川中汞平均浓度为5.6 ng·L-1)[82-84].富含湿地的冰川流域通常有着更高的总汞和甲基汞含量,生物群落中的汞含量亦较高[85].阿拉斯加东南部冰河湾下游集水区,存在湿地的溪流溶解汞的比例大得多(0.1~3.4 ng·L-1),而裸露的冰川径流溶解汞含量仅为0.1~0.9 ng·L-1[85].湿地可以提供丰富的溶解性有机物,同时这种厌氧环境还有利于使汞甲基化的硫酸盐还原菌生存,可以增强微生物的活性和汞甲基化[85-87].汞污染物进入湿地之后被汞甲基化微生物转化为毒性更强的甲基汞再次进入径流,对下游地区人类健康和生态系统存在潜在威胁[88-91].因此,冰川流域生物群落中的甲基汞浓度与流域湿地覆盖率之间存在密切关系,冰川流域湿地可能成为冰川径流中甲基汞净来源,并将其输送到下游生态系统[85,88,92-95]. ...
... ,88,92-95]. ...
Snowmelt, glacial and atmospheric sources of mercury to a subarctic mountain lake catchment, Yukon, Canada
0
2018
Recent glacier variations at the Aconcagua basin, central Chilean Andes
0
2008
Methylmercury cycling in High Arctic wetland ponds: sources and sinks
3
2012
... 冰川对气候变化响应非常敏感,由于气候变暖,冰川正在以前所未有的速度退缩[9].冰川消融不仅给下游生态系统带来水资源,而且还可能释放出在冰川中存储了几个世纪的汞[20].在消融初期,雪冰中的汞以脉冲式释放到冰川径流,近90%的汞会被释放进入融水[76].Sun等[77-78]估算出喜马拉雅地区融水径流汞输出通量为337 kg·a-1,青藏高原冬克玛底流域中汞输出总量约为747.4 g·a-1,亚北极Yukon流域汞输出总量约为4 400 kg·a-1[79-80].Zolkos等[81]对六条北极主要河流进行全年采样调查,估算出总汞输出通量为20 Mg·a-1,Dastoor等[18]估算了北极冰川补给流域总汞年产率为(0.03~3.9 g·km2·a-1).尽管青藏高原河流输出总量较低,但山地冰川(如扎当冰川等)的汞输出通量要高于大部分河流,表明青藏高原山地冰川补给流域具有更高的汞释放效率[30-31,78].在未来全球升温1.5 ℃情景下,到21世纪末青藏高原及周边地区冰川储量将减少2 169~3 217 km3,预估计在未来约11~16吨汞污染物将从冰川释放到下游地区(冰川中汞平均浓度为5.6 ng·L-1)[82-84].富含湿地的冰川流域通常有着更高的总汞和甲基汞含量,生物群落中的汞含量亦较高[85].阿拉斯加东南部冰河湾下游集水区,存在湿地的溪流溶解汞的比例大得多(0.1~3.4 ng·L-1),而裸露的冰川径流溶解汞含量仅为0.1~0.9 ng·L-1[85].湿地可以提供丰富的溶解性有机物,同时这种厌氧环境还有利于使汞甲基化的硫酸盐还原菌生存,可以增强微生物的活性和汞甲基化[85-87].汞污染物进入湿地之后被汞甲基化微生物转化为毒性更强的甲基汞再次进入径流,对下游地区人类健康和生态系统存在潜在威胁[88-91].因此,冰川流域生物群落中的甲基汞浓度与流域湿地覆盖率之间存在密切关系,冰川流域湿地可能成为冰川径流中甲基汞净来源,并将其输送到下游生态系统[85,88,92-95]. ...
... 由于气候变暖,多年冻土快速融化、塌陷,形成热融喀斯特地貌,其中最显著的是形成热融湖塘,此过程更有利于微生物将无机汞转化为甲基汞[22-24,102-104].Tarbier等[24]在亚北极多年冻土区测量了178个土壤样品,甲基汞的浓度范围为0.01~28 ng·g-1,与其他地区土壤甲基汞相比要高得多(例如阿拉斯加北部<0.1~3.4 ng·g-1),同时发现热融池水和热岩孔隙水中甲基汞含量亦较高(0.01~3.1 ng·L-1)[22,24,58,100].在融化阶段,塌陷的冻土沼泽地比完整冻土带活动层的净甲基化程度还要高一个数量级(甲基汞占总汞的比例高出13倍),表明当热融湖塘形成时,冻土区的汞甲基化率将会显著提高[24].这主要是由于冻融塌陷使得冻土泥炭表面被淹没在水位以下,从而形成缺氧环境,这种条件非常有利于具有汞甲基化能力的厌氧微生物生存(包括硫酸盐还原菌、铁还原菌及产甲烷菌等)[58,91,100,104-105]. ...
... Lehnherr等[91,104]在加拿大高纬度地区的湿地湖塘中观测到汞甲基化率很高,来自原位甲基化产生(1.8~40 ng·m-2·d-1)的甲基汞远高于大气沉降(0.03~0.05 ng·m-2·d-1).Fahnestock等[100]发现在亚北极多年冻土消融区汞甲基化微生物有着更高的丰度和多样性.通过对全球多种环境样品的微生物基因组序列比对发现,融化的冻土中汞甲基化基因(hgcA、hgcB)丰度较高,而且这些基因主要存在于产甲烷菌和韧皮菌中[106].北极快速变暖和降水增多正在加速多年冻土的退化,并促进了历史遗留汞释放到现代生物地球化学循环中,多年冻土大幅度退化已经被认为是北极流域汞通量快速增加的主要控制因素[29].前人已在高原观测到大量汞从融化的多年冻土活动层中释放到热融湖塘和其他水体之中[107],然而青藏高原冻土甲基汞浓度的测定以及汞甲基化过程研究目前还处于空白阶段,这些过程极有可能会对高原汞循环产生直接和重大的影响. ...
Mercury cycling in stream ecosystems. 1. Water column chemistry and transport
1
2009
... 冰川对气候变化响应非常敏感,由于气候变暖,冰川正在以前所未有的速度退缩[9].冰川消融不仅给下游生态系统带来水资源,而且还可能释放出在冰川中存储了几个世纪的汞[20].在消融初期,雪冰中的汞以脉冲式释放到冰川径流,近90%的汞会被释放进入融水[76].Sun等[77-78]估算出喜马拉雅地区融水径流汞输出通量为337 kg·a-1,青藏高原冬克玛底流域中汞输出总量约为747.4 g·a-1,亚北极Yukon流域汞输出总量约为4 400 kg·a-1[79-80].Zolkos等[81]对六条北极主要河流进行全年采样调查,估算出总汞输出通量为20 Mg·a-1,Dastoor等[18]估算了北极冰川补给流域总汞年产率为(0.03~3.9 g·km2·a-1).尽管青藏高原河流输出总量较低,但山地冰川(如扎当冰川等)的汞输出通量要高于大部分河流,表明青藏高原山地冰川补给流域具有更高的汞释放效率[30-31,78].在未来全球升温1.5 ℃情景下,到21世纪末青藏高原及周边地区冰川储量将减少2 169~3 217 km3,预估计在未来约11~16吨汞污染物将从冰川释放到下游地区(冰川中汞平均浓度为5.6 ng·L-1)[82-84].富含湿地的冰川流域通常有着更高的总汞和甲基汞含量,生物群落中的汞含量亦较高[85].阿拉斯加东南部冰河湾下游集水区,存在湿地的溪流溶解汞的比例大得多(0.1~3.4 ng·L-1),而裸露的冰川径流溶解汞含量仅为0.1~0.9 ng·L-1[85].湿地可以提供丰富的溶解性有机物,同时这种厌氧环境还有利于使汞甲基化的硫酸盐还原菌生存,可以增强微生物的活性和汞甲基化[85-87].汞污染物进入湿地之后被汞甲基化微生物转化为毒性更强的甲基汞再次进入径流,对下游地区人类健康和生态系统存在潜在威胁[88-91].因此,冰川流域生物群落中的甲基汞浓度与流域湿地覆盖率之间存在密切关系,冰川流域湿地可能成为冰川径流中甲基汞净来源,并将其输送到下游生态系统[85,88,92-95]. ...
Mercury cycling in stream ecosystems. 3. Trophic dynamics and methylmercury bioaccumulation
0
2009
Mercury cycling in terrestrial watersheds
0
2012
The chemistry and transport of mercury in a small wetland in the Adirondack region of New York, USA
1
1998
... 冰川对气候变化响应非常敏感,由于气候变暖,冰川正在以前所未有的速度退缩[9].冰川消融不仅给下游生态系统带来水资源,而且还可能释放出在冰川中存储了几个世纪的汞[20].在消融初期,雪冰中的汞以脉冲式释放到冰川径流,近90%的汞会被释放进入融水[76].Sun等[77-78]估算出喜马拉雅地区融水径流汞输出通量为337 kg·a-1,青藏高原冬克玛底流域中汞输出总量约为747.4 g·a-1,亚北极Yukon流域汞输出总量约为4 400 kg·a-1[79-80].Zolkos等[81]对六条北极主要河流进行全年采样调查,估算出总汞输出通量为20 Mg·a-1,Dastoor等[18]估算了北极冰川补给流域总汞年产率为(0.03~3.9 g·km2·a-1).尽管青藏高原河流输出总量较低,但山地冰川(如扎当冰川等)的汞输出通量要高于大部分河流,表明青藏高原山地冰川补给流域具有更高的汞释放效率[30-31,78].在未来全球升温1.5 ℃情景下,到21世纪末青藏高原及周边地区冰川储量将减少2 169~3 217 km3,预估计在未来约11~16吨汞污染物将从冰川释放到下游地区(冰川中汞平均浓度为5.6 ng·L-1)[82-84].富含湿地的冰川流域通常有着更高的总汞和甲基汞含量,生物群落中的汞含量亦较高[85].阿拉斯加东南部冰河湾下游集水区,存在湿地的溪流溶解汞的比例大得多(0.1~3.4 ng·L-1),而裸露的冰川径流溶解汞含量仅为0.1~0.9 ng·L-1[85].湿地可以提供丰富的溶解性有机物,同时这种厌氧环境还有利于使汞甲基化的硫酸盐还原菌生存,可以增强微生物的活性和汞甲基化[85-87].汞污染物进入湿地之后被汞甲基化微生物转化为毒性更强的甲基汞再次进入径流,对下游地区人类健康和生态系统存在潜在威胁[88-91].因此,冰川流域生物群落中的甲基汞浓度与流域湿地覆盖率之间存在密切关系,冰川流域湿地可能成为冰川径流中甲基汞净来源,并将其输送到下游生态系统[85,88,92-95]. ...
Mercury in wild fish from high-altitude aquatic ecosystems in the Tibetan Plateau
2
2014
... 大量研究表明偏远冰冻圈下游地区水生生态系统甲基汞的富集效应是非常显著的[85,96-97].格陵兰冰盖下游湾峡水生生态系统中,融水径流汞浓度(44~348 ng·L-1)比北极其他河流(0.3~2.8 ng·L-1)高出1~2个数量级,甲基汞浓度(1.0~3.5 ng·L-1)甚至高于大多数淡水,与湿地水平相当[20].我国学者亦研究发现青藏高原河流和湖泊中鱼类甲基汞含量(24.9~1 196 ng·g-1)为中国野生鱼类汞浓度的最高水平[96].此外,前人在冰川补给湖观测到甲基汞的峰值出现在化学跃层,甲基汞占总汞的比例高达57%,同时16S rRNA数据显示检测出硫酸盐还原菌,因此受冰川融水补给的冰川湖化学跃层存在着较强的汞甲基化潜力[97-98]. ...
... [96].此外,前人在冰川补给湖观测到甲基汞的峰值出现在化学跃层,甲基汞占总汞的比例高达57%,同时16S rRNA数据显示检测出硫酸盐还原菌,因此受冰川融水补给的冰川湖化学跃层存在着较强的汞甲基化潜力[97-98]. ...
Mercury geochemistry and microbial diversity in meromictic Glacier Lake, Jamesville, NY
2
2020
... 大量研究表明偏远冰冻圈下游地区水生生态系统甲基汞的富集效应是非常显著的[85,96-97].格陵兰冰盖下游湾峡水生生态系统中,融水径流汞浓度(44~348 ng·L-1)比北极其他河流(0.3~2.8 ng·L-1)高出1~2个数量级,甲基汞浓度(1.0~3.5 ng·L-1)甚至高于大多数淡水,与湿地水平相当[20].我国学者亦研究发现青藏高原河流和湖泊中鱼类甲基汞含量(24.9~1 196 ng·g-1)为中国野生鱼类汞浓度的最高水平[96].此外,前人在冰川补给湖观测到甲基汞的峰值出现在化学跃层,甲基汞占总汞的比例高达57%,同时16S rRNA数据显示检测出硫酸盐还原菌,因此受冰川融水补给的冰川湖化学跃层存在着较强的汞甲基化潜力[97-98]. ...
... [97-98]. ...
Total- and methyl-mercury concentrations and methylation rates across the freshwater to hypersaline continuum of the Great Salt Lake, Utah, USA
1
2015
... 大量研究表明偏远冰冻圈下游地区水生生态系统甲基汞的富集效应是非常显著的[85,96-97].格陵兰冰盖下游湾峡水生生态系统中,融水径流汞浓度(44~348 ng·L-1)比北极其他河流(0.3~2.8 ng·L-1)高出1~2个数量级,甲基汞浓度(1.0~3.5 ng·L-1)甚至高于大多数淡水,与湿地水平相当[20].我国学者亦研究发现青藏高原河流和湖泊中鱼类甲基汞含量(24.9~1 196 ng·g-1)为中国野生鱼类汞浓度的最高水平[96].此外,前人在冰川补给湖观测到甲基汞的峰值出现在化学跃层,甲基汞占总汞的比例高达57%,同时16S rRNA数据显示检测出硫酸盐还原菌,因此受冰川融水补给的冰川湖化学跃层存在着较强的汞甲基化潜力[97-98]. ...
Enhanced availability of mercury bound to dissolved organic matter for methylation in marine sediments
2
2016
... 多年冻土蕴藏着大量的汞存储量,有研究估算了北极苔原土壤储存汞约184 Gg;青藏高原活动层储存汞约16.6 Gg[16-17].冻土甲基汞的分布和转化等过程更为复杂,受各种物理、化学和生物因素控制,包括温度、溶解性有机质(DOC)、碱度、pH、总汞浓度、微生物以及其他金属元素或离子(如铁、锰)等[22,46,99-101].MacMillan等[22]发现冻土环境中甲基汞与溶解有机碳等营养物质,以及甲基汞与微生物之间有很强的相关性.当温度较高(8 ℃)时,在富含有机物和矿物质的土壤层中,甲基汞净产量比温度较低(-2 ℃)时增加了10倍以上[23].可溶性土壤有机碳类型及其生物可利用性,对促进甲基汞的初始合成特别重要[99].当冻土活动层融化时,低分子量溶解有机质增加促使形成大量生物可利用的Hg-DOM复合物,从而使无机汞的可利用性增加,更有利于冻土汞甲基化[23-24,100]. ...
... [99].当冻土活动层融化时,低分子量溶解有机质增加促使形成大量生物可利用的Hg-DOM复合物,从而使无机汞的可利用性增加,更有利于冻土汞甲基化[23-24,100]. ...
Mercury reallocation in thawing subarctic peatlands
4
2019
... 多年冻土蕴藏着大量的汞存储量,有研究估算了北极苔原土壤储存汞约184 Gg;青藏高原活动层储存汞约16.6 Gg[16-17].冻土甲基汞的分布和转化等过程更为复杂,受各种物理、化学和生物因素控制,包括温度、溶解性有机质(DOC)、碱度、pH、总汞浓度、微生物以及其他金属元素或离子(如铁、锰)等[22,46,99-101].MacMillan等[22]发现冻土环境中甲基汞与溶解有机碳等营养物质,以及甲基汞与微生物之间有很强的相关性.当温度较高(8 ℃)时,在富含有机物和矿物质的土壤层中,甲基汞净产量比温度较低(-2 ℃)时增加了10倍以上[23].可溶性土壤有机碳类型及其生物可利用性,对促进甲基汞的初始合成特别重要[99].当冻土活动层融化时,低分子量溶解有机质增加促使形成大量生物可利用的Hg-DOM复合物,从而使无机汞的可利用性增加,更有利于冻土汞甲基化[23-24,100]. ...
... 由于气候变暖,多年冻土快速融化、塌陷,形成热融喀斯特地貌,其中最显著的是形成热融湖塘,此过程更有利于微生物将无机汞转化为甲基汞[22-24,102-104].Tarbier等[24]在亚北极多年冻土区测量了178个土壤样品,甲基汞的浓度范围为0.01~28 ng·g-1,与其他地区土壤甲基汞相比要高得多(例如阿拉斯加北部<0.1~3.4 ng·g-1),同时发现热融池水和热岩孔隙水中甲基汞含量亦较高(0.01~3.1 ng·L-1)[22,24,58,100].在融化阶段,塌陷的冻土沼泽地比完整冻土带活动层的净甲基化程度还要高一个数量级(甲基汞占总汞的比例高出13倍),表明当热融湖塘形成时,冻土区的汞甲基化率将会显著提高[24].这主要是由于冻融塌陷使得冻土泥炭表面被淹没在水位以下,从而形成缺氧环境,这种条件非常有利于具有汞甲基化能力的厌氧微生物生存(包括硫酸盐还原菌、铁还原菌及产甲烷菌等)[58,91,100,104-105]. ...
... ,100,104-105]. ...
... Lehnherr等[91,104]在加拿大高纬度地区的湿地湖塘中观测到汞甲基化率很高,来自原位甲基化产生(1.8~40 ng·m-2·d-1)的甲基汞远高于大气沉降(0.03~0.05 ng·m-2·d-1).Fahnestock等[100]发现在亚北极多年冻土消融区汞甲基化微生物有着更高的丰度和多样性.通过对全球多种环境样品的微生物基因组序列比对发现,融化的冻土中汞甲基化基因(hgcA、hgcB)丰度较高,而且这些基因主要存在于产甲烷菌和韧皮菌中[106].北极快速变暖和降水增多正在加速多年冻土的退化,并促进了历史遗留汞释放到现代生物地球化学循环中,多年冻土大幅度退化已经被认为是北极流域汞通量快速增加的主要控制因素[29].前人已在高原观测到大量汞从融化的多年冻土活动层中释放到热融湖塘和其他水体之中[107],然而青藏高原冻土甲基汞浓度的测定以及汞甲基化过程研究目前还处于空白阶段,这些过程极有可能会对高原汞循环产生直接和重大的影响. ...
Significant interaction effects from sulfate deposition and climate on sulfur concentrations constitute major controls on methylmercury production in peatlands
1
2013
... 多年冻土蕴藏着大量的汞存储量,有研究估算了北极苔原土壤储存汞约184 Gg;青藏高原活动层储存汞约16.6 Gg[16-17].冻土甲基汞的分布和转化等过程更为复杂,受各种物理、化学和生物因素控制,包括温度、溶解性有机质(DOC)、碱度、pH、总汞浓度、微生物以及其他金属元素或离子(如铁、锰)等[22,46,99-101].MacMillan等[22]发现冻土环境中甲基汞与溶解有机碳等营养物质,以及甲基汞与微生物之间有很强的相关性.当温度较高(8 ℃)时,在富含有机物和矿物质的土壤层中,甲基汞净产量比温度较低(-2 ℃)时增加了10倍以上[23].可溶性土壤有机碳类型及其生物可利用性,对促进甲基汞的初始合成特别重要[99].当冻土活动层融化时,低分子量溶解有机质增加促使形成大量生物可利用的Hg-DOM复合物,从而使无机汞的可利用性增加,更有利于冻土汞甲基化[23-24,100]. ...
Permafrost is warming at a global scale
1
2019
... 由于气候变暖,多年冻土快速融化、塌陷,形成热融喀斯特地貌,其中最显著的是形成热融湖塘,此过程更有利于微生物将无机汞转化为甲基汞[22-24,102-104].Tarbier等[24]在亚北极多年冻土区测量了178个土壤样品,甲基汞的浓度范围为0.01~28 ng·g-1,与其他地区土壤甲基汞相比要高得多(例如阿拉斯加北部<0.1~3.4 ng·g-1),同时发现热融池水和热岩孔隙水中甲基汞含量亦较高(0.01~3.1 ng·L-1)[22,24,58,100].在融化阶段,塌陷的冻土沼泽地比完整冻土带活动层的净甲基化程度还要高一个数量级(甲基汞占总汞的比例高出13倍),表明当热融湖塘形成时,冻土区的汞甲基化率将会显著提高[24].这主要是由于冻融塌陷使得冻土泥炭表面被淹没在水位以下,从而形成缺氧环境,这种条件非常有利于具有汞甲基化能力的厌氧微生物生存(包括硫酸盐还原菌、铁还原菌及产甲烷菌等)[58,91,100,104-105]. ...
Strong degradation of palsas and peat plateaus in northern Norway during the last 60 years
0
2017
Methylmercury biogeochemistry: a review with special reference to Arctic aquatic ecosystems
4
2014
... 由于气候变暖,多年冻土快速融化、塌陷,形成热融喀斯特地貌,其中最显著的是形成热融湖塘,此过程更有利于微生物将无机汞转化为甲基汞[22-24,102-104].Tarbier等[24]在亚北极多年冻土区测量了178个土壤样品,甲基汞的浓度范围为0.01~28 ng·g-1,与其他地区土壤甲基汞相比要高得多(例如阿拉斯加北部<0.1~3.4 ng·g-1),同时发现热融池水和热岩孔隙水中甲基汞含量亦较高(0.01~3.1 ng·L-1)[22,24,58,100].在融化阶段,塌陷的冻土沼泽地比完整冻土带活动层的净甲基化程度还要高一个数量级(甲基汞占总汞的比例高出13倍),表明当热融湖塘形成时,冻土区的汞甲基化率将会显著提高[24].这主要是由于冻融塌陷使得冻土泥炭表面被淹没在水位以下,从而形成缺氧环境,这种条件非常有利于具有汞甲基化能力的厌氧微生物生存(包括硫酸盐还原菌、铁还原菌及产甲烷菌等)[58,91,100,104-105]. ...
... ,104-105]. ...
... Lehnherr等[91,104]在加拿大高纬度地区的湿地湖塘中观测到汞甲基化率很高,来自原位甲基化产生(1.8~40 ng·m-2·d-1)的甲基汞远高于大气沉降(0.03~0.05 ng·m-2·d-1).Fahnestock等[100]发现在亚北极多年冻土消融区汞甲基化微生物有着更高的丰度和多样性.通过对全球多种环境样品的微生物基因组序列比对发现,融化的冻土中汞甲基化基因(hgcA、hgcB)丰度较高,而且这些基因主要存在于产甲烷菌和韧皮菌中[106].北极快速变暖和降水增多正在加速多年冻土的退化,并促进了历史遗留汞释放到现代生物地球化学循环中,多年冻土大幅度退化已经被认为是北极流域汞通量快速增加的主要控制因素[29].前人已在高原观测到大量汞从融化的多年冻土活动层中释放到热融湖塘和其他水体之中[107],然而青藏高原冻土甲基汞浓度的测定以及汞甲基化过程研究目前还处于空白阶段,这些过程极有可能会对高原汞循环产生直接和重大的影响. ...
... 冰冻圈甲基汞富集作用同样对生活在该区域的人类健康构成重大威胁.北极地区居住着土著居民(如因纽特人),他们依赖于捕获鱼类和海洋哺乳动物为食,通过饮食摄入是当地居民暴露甲基汞的主要途径[3,155-157].北极因纽特人原住民由于常年捕食海洋生物,人体内的甲基汞含量都远超过了毒理学阈值[104,158].由于甲基汞对成人和儿童的毒性效应有所不同,处于发育中的儿童中枢神经系统比成年人对甲基汞更为敏感,而孕妇长期处于低剂量甲基汞暴露将会对婴儿的智力发育产生严重影响[33].研究表明,北极一些地区50%以上的母亲和育龄妇女的血液中的汞含量已超过美国环境保护署建议的标准浓度(5.8 μg·L-1),且在格陵兰岛部分地区近乎90%的育龄妇女血液中的汞含量都超标,这些都表明我们迫切需要重视冰冻圈甲基汞暴露风险[33,157-158]. ...
Biogeochemical factors affecting mercury methylation in sediments of the Venice Lagoon, Italy
1
2007
... 由于气候变暖,多年冻土快速融化、塌陷,形成热融喀斯特地貌,其中最显著的是形成热融湖塘,此过程更有利于微生物将无机汞转化为甲基汞[22-24,102-104].Tarbier等[24]在亚北极多年冻土区测量了178个土壤样品,甲基汞的浓度范围为0.01~28 ng·g-1,与其他地区土壤甲基汞相比要高得多(例如阿拉斯加北部<0.1~3.4 ng·g-1),同时发现热融池水和热岩孔隙水中甲基汞含量亦较高(0.01~3.1 ng·L-1)[22,24,58,100].在融化阶段,塌陷的冻土沼泽地比完整冻土带活动层的净甲基化程度还要高一个数量级(甲基汞占总汞的比例高出13倍),表明当热融湖塘形成时,冻土区的汞甲基化率将会显著提高[24].这主要是由于冻融塌陷使得冻土泥炭表面被淹没在水位以下,从而形成缺氧环境,这种条件非常有利于具有汞甲基化能力的厌氧微生物生存(包括硫酸盐还原菌、铁还原菌及产甲烷菌等)[58,91,100,104-105]. ...
Global prevalence and distribution of genes and microorganisms involved in mercury methylation
1
2015
... Lehnherr等[91,104]在加拿大高纬度地区的湿地湖塘中观测到汞甲基化率很高,来自原位甲基化产生(1.8~40 ng·m-2·d-1)的甲基汞远高于大气沉降(0.03~0.05 ng·m-2·d-1).Fahnestock等[100]发现在亚北极多年冻土消融区汞甲基化微生物有着更高的丰度和多样性.通过对全球多种环境样品的微生物基因组序列比对发现,融化的冻土中汞甲基化基因(hgcA、hgcB)丰度较高,而且这些基因主要存在于产甲烷菌和韧皮菌中[106].北极快速变暖和降水增多正在加速多年冻土的退化,并促进了历史遗留汞释放到现代生物地球化学循环中,多年冻土大幅度退化已经被认为是北极流域汞通量快速增加的主要控制因素[29].前人已在高原观测到大量汞从融化的多年冻土活动层中释放到热融湖塘和其他水体之中[107],然而青藏高原冻土甲基汞浓度的测定以及汞甲基化过程研究目前还处于空白阶段,这些过程极有可能会对高原汞循环产生直接和重大的影响. ...
Permafrost thaw dominates mercury emission in Tibetan thermokarst ponds
1
2020
... Lehnherr等[91,104]在加拿大高纬度地区的湿地湖塘中观测到汞甲基化率很高,来自原位甲基化产生(1.8~40 ng·m-2·d-1)的甲基汞远高于大气沉降(0.03~0.05 ng·m-2·d-1).Fahnestock等[100]发现在亚北极多年冻土消融区汞甲基化微生物有着更高的丰度和多样性.通过对全球多种环境样品的微生物基因组序列比对发现,融化的冻土中汞甲基化基因(hgcA、hgcB)丰度较高,而且这些基因主要存在于产甲烷菌和韧皮菌中[106].北极快速变暖和降水增多正在加速多年冻土的退化,并促进了历史遗留汞释放到现代生物地球化学循环中,多年冻土大幅度退化已经被认为是北极流域汞通量快速增加的主要控制因素[29].前人已在高原观测到大量汞从融化的多年冻土活动层中释放到热融湖塘和其他水体之中[107],然而青藏高原冻土甲基汞浓度的测定以及汞甲基化过程研究目前还处于空白阶段,这些过程极有可能会对高原汞循环产生直接和重大的影响. ...
Some sources and sinks of monomethyl and inorganic mercury on Ellesmere Island in the Canadian High Arctic
6
2005
... 降雪是冰冻圈大气降水最重要的表现形式,是大气汞进入冰冻圈的重要环节[108].积雪亦是一种活跃的媒介,消融后将成为环境汞的重要来源[60,109-110].在南北极地区,南极(0.1~1 ng·L-1)与北极(1~5 ng·L-1)表层雪冰中的总汞浓度在冬季均很低;然而在春季,受大气汞亏损事件(Atmospheric Mercury Depletion Events, AMDEs)的影响,南北极雪冰中汞的浓度均显著增加[111-119].早期研究表明积雪中甲基汞与大气沉降密切相关,但在AMDEs期间甲基汞与无机汞表现出完全不同的行为模式[108,111].Constant等[120]调查了加拿大亚北极地区AMDEs时期雪冰中的甲基汞变化规律,结果表明甲基汞的来源与AMDEs无关.雪冰甲基汞与氯化物之间存在较强的相关性,表明沿海地区积雪甲基汞可能来自海盐气溶胶[108].海洋中浮游植物可以产生二甲基汞(DMeHg),由于二甲基汞具有较高的挥发性,它可以从海洋释放到大气中,被自由基氧化成单甲基汞,然后随海盐沉降到附近的雪面之上[108,121-123].值得注意的是新沉降的甲基汞在积雪中并不稳定,15%~56%的甲基汞会在夜间通过去甲基化或以其他方式损失[124]. ...
... [108,111].Constant等[120]调查了加拿大亚北极地区AMDEs时期雪冰中的甲基汞变化规律,结果表明甲基汞的来源与AMDEs无关.雪冰甲基汞与氯化物之间存在较强的相关性,表明沿海地区积雪甲基汞可能来自海盐气溶胶[108].海洋中浮游植物可以产生二甲基汞(DMeHg),由于二甲基汞具有较高的挥发性,它可以从海洋释放到大气中,被自由基氧化成单甲基汞,然后随海盐沉降到附近的雪面之上[108,121-123].值得注意的是新沉降的甲基汞在积雪中并不稳定,15%~56%的甲基汞会在夜间通过去甲基化或以其他方式损失[124]. ...
... [108].海洋中浮游植物可以产生二甲基汞(DMeHg),由于二甲基汞具有较高的挥发性,它可以从海洋释放到大气中,被自由基氧化成单甲基汞,然后随海盐沉降到附近的雪面之上[108,121-123].值得注意的是新沉降的甲基汞在积雪中并不稳定,15%~56%的甲基汞会在夜间通过去甲基化或以其他方式损失[124]. ...
... [108,121-123].值得注意的是新沉降的甲基汞在积雪中并不稳定,15%~56%的甲基汞会在夜间通过去甲基化或以其他方式损失[124]. ...
... 积雪中甲基汞的来源在过去被归因于大气沉降,然而随着研究的不断深入,新的证据表明在低纬度苔原的积雪中可能存在着较强的汞甲基化活动[108,111,123-127].在融雪期间,雪冰融水中甲基汞占总汞的比例增大(从2.7%增加到7.6%)[126].导致这一现象的原因一方面由于甲基汞从无机/有机物中浸出,在雪冰融化时趋于集中;另一方面是由于雪冰内部可能发生甲基化作用,微生物活动和反应可能在积雪中产生了甲基汞[108,120,123-127].极地积雪的pH值通常为酸性,有机物以及微生物等存在于雪层中,因此在消融的积雪中存在微生物汞甲基化的可能[120].随着气候变暖,温度升高会促使积雪中微生物种群数量增长,从而导致微生物影响汞甲基化的强度大幅增加[25].Loseto等[60]估算出北极地区积雪中甲基汞产量约为1.5 mg·km-2·d-1,并指出北极康沃利斯岛陆地积雪融化是地表径流中最重要的甲基汞来源. ...
... [108,120,123-127].极地积雪的pH值通常为酸性,有机物以及微生物等存在于雪层中,因此在消融的积雪中存在微生物汞甲基化的可能[120].随着气候变暖,温度升高会促使积雪中微生物种群数量增长,从而导致微生物影响汞甲基化的强度大幅增加[25].Loseto等[60]估算出北极地区积雪中甲基汞产量约为1.5 mg·km-2·d-1,并指出北极康沃利斯岛陆地积雪融化是地表径流中最重要的甲基汞来源. ...
An overview of snow photochemistry: evidence, mechanisms and impacts
1
2007
... 降雪是冰冻圈大气降水最重要的表现形式,是大气汞进入冰冻圈的重要环节[108].积雪亦是一种活跃的媒介,消融后将成为环境汞的重要来源[60,109-110].在南北极地区,南极(0.1~1 ng·L-1)与北极(1~5 ng·L-1)表层雪冰中的总汞浓度在冬季均很低;然而在春季,受大气汞亏损事件(Atmospheric Mercury Depletion Events, AMDEs)的影响,南北极雪冰中汞的浓度均显著增加[111-119].早期研究表明积雪中甲基汞与大气沉降密切相关,但在AMDEs期间甲基汞与无机汞表现出完全不同的行为模式[108,111].Constant等[120]调查了加拿大亚北极地区AMDEs时期雪冰中的甲基汞变化规律,结果表明甲基汞的来源与AMDEs无关.雪冰甲基汞与氯化物之间存在较强的相关性,表明沿海地区积雪甲基汞可能来自海盐气溶胶[108].海洋中浮游植物可以产生二甲基汞(DMeHg),由于二甲基汞具有较高的挥发性,它可以从海洋释放到大气中,被自由基氧化成单甲基汞,然后随海盐沉降到附近的雪面之上[108,121-123].值得注意的是新沉降的甲基汞在积雪中并不稳定,15%~56%的甲基汞会在夜间通过去甲基化或以其他方式损失[124]. ...
A simulation of the enrichment of atmospheric pollutants in snow cover runoff
1
1981
... 降雪是冰冻圈大气降水最重要的表现形式,是大气汞进入冰冻圈的重要环节[108].积雪亦是一种活跃的媒介,消融后将成为环境汞的重要来源[60,109-110].在南北极地区,南极(0.1~1 ng·L-1)与北极(1~5 ng·L-1)表层雪冰中的总汞浓度在冬季均很低;然而在春季,受大气汞亏损事件(Atmospheric Mercury Depletion Events, AMDEs)的影响,南北极雪冰中汞的浓度均显著增加[111-119].早期研究表明积雪中甲基汞与大气沉降密切相关,但在AMDEs期间甲基汞与无机汞表现出完全不同的行为模式[108,111].Constant等[120]调查了加拿大亚北极地区AMDEs时期雪冰中的甲基汞变化规律,结果表明甲基汞的来源与AMDEs无关.雪冰甲基汞与氯化物之间存在较强的相关性,表明沿海地区积雪甲基汞可能来自海盐气溶胶[108].海洋中浮游植物可以产生二甲基汞(DMeHg),由于二甲基汞具有较高的挥发性,它可以从海洋释放到大气中,被自由基氧化成单甲基汞,然后随海盐沉降到附近的雪面之上[108,121-123].值得注意的是新沉降的甲基汞在积雪中并不稳定,15%~56%的甲基汞会在夜间通过去甲基化或以其他方式损失[124]. ...
Total and methyl mercury patterns in Arctic snow during springtime at Resolute, Nunavut, Canada
3
2005
... 降雪是冰冻圈大气降水最重要的表现形式,是大气汞进入冰冻圈的重要环节[108].积雪亦是一种活跃的媒介,消融后将成为环境汞的重要来源[60,109-110].在南北极地区,南极(0.1~1 ng·L-1)与北极(1~5 ng·L-1)表层雪冰中的总汞浓度在冬季均很低;然而在春季,受大气汞亏损事件(Atmospheric Mercury Depletion Events, AMDEs)的影响,南北极雪冰中汞的浓度均显著增加[111-119].早期研究表明积雪中甲基汞与大气沉降密切相关,但在AMDEs期间甲基汞与无机汞表现出完全不同的行为模式[108,111].Constant等[120]调查了加拿大亚北极地区AMDEs时期雪冰中的甲基汞变化规律,结果表明甲基汞的来源与AMDEs无关.雪冰甲基汞与氯化物之间存在较强的相关性,表明沿海地区积雪甲基汞可能来自海盐气溶胶[108].海洋中浮游植物可以产生二甲基汞(DMeHg),由于二甲基汞具有较高的挥发性,它可以从海洋释放到大气中,被自由基氧化成单甲基汞,然后随海盐沉降到附近的雪面之上[108,121-123].值得注意的是新沉降的甲基汞在积雪中并不稳定,15%~56%的甲基汞会在夜间通过去甲基化或以其他方式损失[124]. ...
... ,111].Constant等[120]调查了加拿大亚北极地区AMDEs时期雪冰中的甲基汞变化规律,结果表明甲基汞的来源与AMDEs无关.雪冰甲基汞与氯化物之间存在较强的相关性,表明沿海地区积雪甲基汞可能来自海盐气溶胶[108].海洋中浮游植物可以产生二甲基汞(DMeHg),由于二甲基汞具有较高的挥发性,它可以从海洋释放到大气中,被自由基氧化成单甲基汞,然后随海盐沉降到附近的雪面之上[108,121-123].值得注意的是新沉降的甲基汞在积雪中并不稳定,15%~56%的甲基汞会在夜间通过去甲基化或以其他方式损失[124]. ...
... 积雪中甲基汞的来源在过去被归因于大气沉降,然而随着研究的不断深入,新的证据表明在低纬度苔原的积雪中可能存在着较强的汞甲基化活动[108,111,123-127].在融雪期间,雪冰融水中甲基汞占总汞的比例增大(从2.7%增加到7.6%)[126].导致这一现象的原因一方面由于甲基汞从无机/有机物中浸出,在雪冰融化时趋于集中;另一方面是由于雪冰内部可能发生甲基化作用,微生物活动和反应可能在积雪中产生了甲基汞[108,120,123-127].极地积雪的pH值通常为酸性,有机物以及微生物等存在于雪层中,因此在消融的积雪中存在微生物汞甲基化的可能[120].随着气候变暖,温度升高会促使积雪中微生物种群数量增长,从而导致微生物影响汞甲基化的强度大幅增加[25].Loseto等[60]估算出北极地区积雪中甲基汞产量约为1.5 mg·km-2·d-1,并指出北极康沃利斯岛陆地积雪融化是地表径流中最重要的甲基汞来源. ...
Arctic springtime depletion of mercury
0
1998
Mercury content of Antarctic surface snow: initial results
0
1990
Mercury in snow layers from the Antarctica
1
1998
... Concentrations of MeHg in the cryospheric environment
Table 2| 地点 | 样品类型 | 甲基汞 | 总汞 | 文献来源 |
|---|
| Devon Island, Canada | 多年冻土 | 0.3~3.1 ng·g-1 | — | [58] |
| Barrow, Alaska | 热融湖塘 | 0.3~2.3 ng·L-1 | 25.3~141.2 ng·L-1 | [23] |
| Tibetan Plateau, China | 多年冻土 | — | 5.6~39.3 ng·g-1 | [17] |
| Fennoscandia | 多年冻土 | 0.004~28 ng·g-1 | 1.1~210 ng·g-1 | [24] |
| Tussock tundra, Alaska | 多年冻土 | — | 56~226 ng·g-1 | [16] |
| Laohugou No.12, Tibetan Plateau, China | 雪冰 | 0.005~0.5 ng·L-1 | 24.4~172.1 ng·L-1 | [56] |
| Tibetan Plateau, China | 冰尘 | 0.4~1.7 ng·g-1 | 17.9~114.5 ng·g-1 | [19] |
| French Alps | 表层雪 | 0.002~0.3 ng·L-1 | 1.8~169.5 ng·L-1 | [57] |
| Spitsbergen Island, Norway | 表层雪 | 0.004~0.5 ng·L-1 | 1~90 ng·L-1 | [123] |
| Resolute Bay, High Arctic, Canada | 表层雪 | <0.015~0.2 ng·L-1 | 2.6~150 ng·L-1 | [125] |
| Hudson Bay, Canada | 表层雪 | 0.04~0.14 ng·L-1 | 1.6~5.1 ng·L-1 | [120] |
| Alberta, Canada | 表层雪 | 0.02~0.3 ng·L-1 | 0.8~9.0 ng·L-1 | [137] |
| Antarctic | 积雪 | <0.02~0.3 ng·L-1 | 1.0~30.6 ng·L-1 | [59] |
| McMurdo/Ross Island, Antarctic | 积雪 | — | 40~430 ng·L-1 | [114] |
| Casey Station, Antarctic | 海冰 | <0.009~0.09 ng·L-1 | 0.2~5.7 ng·L-1 | [129] |
| Antarctic | 海冰 | <0.02~0.2 ng·L-1 | 1.4~179 ng·L-1 | [59] |
| Beaufort Sea and McClure Strait, Arctic | 海冰 | <0.02~0.5 ng·L-1 | 0.1~12.2 ng·L-1 | [28] |
| Southern Ocean | 海水 | <0.02~0.1 ng·L-1 | 0. 2~23 ng·L-1 | [59] |
| Arctic Ocean | 海水 | 0.006~0.07 ng·L-1 | 0.09~1.4 ng·L-1 | [135] |
| Resolute Bay, High Arctic, Canada | 海水 | 0.06~0.1 ng·L-1 | 0.1~0.2 ng·L-1 | [125] |
注:表格中海水均为海冰覆盖水域. ...
Mercury content of Antarctic ice and snow: further results
0
1991
A synthesis of atmospheric mercury depletion event chemistry linking atmosphere, snow and water
0
2007
Measurements of atmospheric mercury species during an international study of mercury depletion events at Ny-?lesund, Svalbard, spring 2003. How reproducible are our present methods?
0
2005
Snow-to-air exchanges of mercury in an Arctic seasonal snow pack in Ny-?lesund, Svalbard
0
2005
Atmospheric mercury concentrations: measurements and profiles near snow and ice surfaces in the Canadian Arctic during Alert 2000
1
2002
... 降雪是冰冻圈大气降水最重要的表现形式,是大气汞进入冰冻圈的重要环节[108].积雪亦是一种活跃的媒介,消融后将成为环境汞的重要来源[60,109-110].在南北极地区,南极(0.1~1 ng·L-1)与北极(1~5 ng·L-1)表层雪冰中的总汞浓度在冬季均很低;然而在春季,受大气汞亏损事件(Atmospheric Mercury Depletion Events, AMDEs)的影响,南北极雪冰中汞的浓度均显著增加[111-119].早期研究表明积雪中甲基汞与大气沉降密切相关,但在AMDEs期间甲基汞与无机汞表现出完全不同的行为模式[108,111].Constant等[120]调查了加拿大亚北极地区AMDEs时期雪冰中的甲基汞变化规律,结果表明甲基汞的来源与AMDEs无关.雪冰甲基汞与氯化物之间存在较强的相关性,表明沿海地区积雪甲基汞可能来自海盐气溶胶[108].海洋中浮游植物可以产生二甲基汞(DMeHg),由于二甲基汞具有较高的挥发性,它可以从海洋释放到大气中,被自由基氧化成单甲基汞,然后随海盐沉降到附近的雪面之上[108,121-123].值得注意的是新沉降的甲基汞在积雪中并不稳定,15%~56%的甲基汞会在夜间通过去甲基化或以其他方式损失[124]. ...
Fate of inorganic mercury and methyl mercury within the snow cover in the low arctic tundra on the shore of Hudson Bay (Québec, Canada)
4
2007
... 降雪是冰冻圈大气降水最重要的表现形式,是大气汞进入冰冻圈的重要环节[108].积雪亦是一种活跃的媒介,消融后将成为环境汞的重要来源[60,109-110].在南北极地区,南极(0.1~1 ng·L-1)与北极(1~5 ng·L-1)表层雪冰中的总汞浓度在冬季均很低;然而在春季,受大气汞亏损事件(Atmospheric Mercury Depletion Events, AMDEs)的影响,南北极雪冰中汞的浓度均显著增加[111-119].早期研究表明积雪中甲基汞与大气沉降密切相关,但在AMDEs期间甲基汞与无机汞表现出完全不同的行为模式[108,111].Constant等[120]调查了加拿大亚北极地区AMDEs时期雪冰中的甲基汞变化规律,结果表明甲基汞的来源与AMDEs无关.雪冰甲基汞与氯化物之间存在较强的相关性,表明沿海地区积雪甲基汞可能来自海盐气溶胶[108].海洋中浮游植物可以产生二甲基汞(DMeHg),由于二甲基汞具有较高的挥发性,它可以从海洋释放到大气中,被自由基氧化成单甲基汞,然后随海盐沉降到附近的雪面之上[108,121-123].值得注意的是新沉降的甲基汞在积雪中并不稳定,15%~56%的甲基汞会在夜间通过去甲基化或以其他方式损失[124]. ...
... 积雪中甲基汞的来源在过去被归因于大气沉降,然而随着研究的不断深入,新的证据表明在低纬度苔原的积雪中可能存在着较强的汞甲基化活动[108,111,123-127].在融雪期间,雪冰融水中甲基汞占总汞的比例增大(从2.7%增加到7.6%)[126].导致这一现象的原因一方面由于甲基汞从无机/有机物中浸出,在雪冰融化时趋于集中;另一方面是由于雪冰内部可能发生甲基化作用,微生物活动和反应可能在积雪中产生了甲基汞[108,120,123-127].极地积雪的pH值通常为酸性,有机物以及微生物等存在于雪层中,因此在消融的积雪中存在微生物汞甲基化的可能[120].随着气候变暖,温度升高会促使积雪中微生物种群数量增长,从而导致微生物影响汞甲基化的强度大幅增加[25].Loseto等[60]估算出北极地区积雪中甲基汞产量约为1.5 mg·km-2·d-1,并指出北极康沃利斯岛陆地积雪融化是地表径流中最重要的甲基汞来源. ...
... [120].随着气候变暖,温度升高会促使积雪中微生物种群数量增长,从而导致微生物影响汞甲基化的强度大幅增加[25].Loseto等[60]估算出北极地区积雪中甲基汞产量约为1.5 mg·km-2·d-1,并指出北极康沃利斯岛陆地积雪融化是地表径流中最重要的甲基汞来源. ...
... Concentrations of MeHg in the cryospheric environment
Table 2| 地点 | 样品类型 | 甲基汞 | 总汞 | 文献来源 |
|---|
| Devon Island, Canada | 多年冻土 | 0.3~3.1 ng·g-1 | — | [58] |
| Barrow, Alaska | 热融湖塘 | 0.3~2.3 ng·L-1 | 25.3~141.2 ng·L-1 | [23] |
| Tibetan Plateau, China | 多年冻土 | — | 5.6~39.3 ng·g-1 | [17] |
| Fennoscandia | 多年冻土 | 0.004~28 ng·g-1 | 1.1~210 ng·g-1 | [24] |
| Tussock tundra, Alaska | 多年冻土 | — | 56~226 ng·g-1 | [16] |
| Laohugou No.12, Tibetan Plateau, China | 雪冰 | 0.005~0.5 ng·L-1 | 24.4~172.1 ng·L-1 | [56] |
| Tibetan Plateau, China | 冰尘 | 0.4~1.7 ng·g-1 | 17.9~114.5 ng·g-1 | [19] |
| French Alps | 表层雪 | 0.002~0.3 ng·L-1 | 1.8~169.5 ng·L-1 | [57] |
| Spitsbergen Island, Norway | 表层雪 | 0.004~0.5 ng·L-1 | 1~90 ng·L-1 | [123] |
| Resolute Bay, High Arctic, Canada | 表层雪 | <0.015~0.2 ng·L-1 | 2.6~150 ng·L-1 | [125] |
| Hudson Bay, Canada | 表层雪 | 0.04~0.14 ng·L-1 | 1.6~5.1 ng·L-1 | [120] |
| Alberta, Canada | 表层雪 | 0.02~0.3 ng·L-1 | 0.8~9.0 ng·L-1 | [137] |
| Antarctic | 积雪 | <0.02~0.3 ng·L-1 | 1.0~30.6 ng·L-1 | [59] |
| McMurdo/Ross Island, Antarctic | 积雪 | — | 40~430 ng·L-1 | [114] |
| Casey Station, Antarctic | 海冰 | <0.009~0.09 ng·L-1 | 0.2~5.7 ng·L-1 | [129] |
| Antarctic | 海冰 | <0.02~0.2 ng·L-1 | 1.4~179 ng·L-1 | [59] |
| Beaufort Sea and McClure Strait, Arctic | 海冰 | <0.02~0.5 ng·L-1 | 0.1~12.2 ng·L-1 | [28] |
| Southern Ocean | 海水 | <0.02~0.1 ng·L-1 | 0. 2~23 ng·L-1 | [59] |
| Arctic Ocean | 海水 | 0.006~0.07 ng·L-1 | 0.09~1.4 ng·L-1 | [135] |
| Resolute Bay, High Arctic, Canada | 海水 | 0.06~0.1 ng·L-1 | 0.1~0.2 ng·L-1 | [125] |
注:表格中海水均为海冰覆盖水域. ...
Production of methylated mercury, lead, and cadmium by marine bacteria as a significant natural source for atmospheric heavy metals in polar regions
2
1999
... 降雪是冰冻圈大气降水最重要的表现形式,是大气汞进入冰冻圈的重要环节[108].积雪亦是一种活跃的媒介,消融后将成为环境汞的重要来源[60,109-110].在南北极地区,南极(0.1~1 ng·L-1)与北极(1~5 ng·L-1)表层雪冰中的总汞浓度在冬季均很低;然而在春季,受大气汞亏损事件(Atmospheric Mercury Depletion Events, AMDEs)的影响,南北极雪冰中汞的浓度均显著增加[111-119].早期研究表明积雪中甲基汞与大气沉降密切相关,但在AMDEs期间甲基汞与无机汞表现出完全不同的行为模式[108,111].Constant等[120]调查了加拿大亚北极地区AMDEs时期雪冰中的甲基汞变化规律,结果表明甲基汞的来源与AMDEs无关.雪冰甲基汞与氯化物之间存在较强的相关性,表明沿海地区积雪甲基汞可能来自海盐气溶胶[108].海洋中浮游植物可以产生二甲基汞(DMeHg),由于二甲基汞具有较高的挥发性,它可以从海洋释放到大气中,被自由基氧化成单甲基汞,然后随海盐沉降到附近的雪面之上[108,121-123].值得注意的是新沉降的甲基汞在积雪中并不稳定,15%~56%的甲基汞会在夜间通过去甲基化或以其他方式损失[124]. ...
... 海冰中的甲基汞来源主要包括大气沉降和原位汞甲基化.海洋中的浮游植物生产的二甲基汞从海面释放,在大气中降解为甲基汞,最后又沉降到海冰或海水中,这是海冰甲基汞重要来源之一[121].Baya等[128]研究指出海冰覆盖和初级生产是促进海洋浮游植物生产二甲基汞的主要驱动因素.海冰甲基汞来源除了大气和海洋之外,海冰原位产甲基汞亦非常重要.大量研究已表明海冰中下部和冰水界面是已知的可能产生甲基汞的区位[28,129].因此,海冰中汞的原位甲基化使得海冰融化极有可能成为海洋生态系统新的甲基汞来源. ...
A long-path Fourier transform infrared study of the kinetics and mechanism for the hydroxyl radical-initiated oxidation of dimethylmercury
0
1983
Springtime changes in snow chemistry lead to new insights into mercury methylation in the Arctic
4
2010
... 降雪是冰冻圈大气降水最重要的表现形式,是大气汞进入冰冻圈的重要环节[108].积雪亦是一种活跃的媒介,消融后将成为环境汞的重要来源[60,109-110].在南北极地区,南极(0.1~1 ng·L-1)与北极(1~5 ng·L-1)表层雪冰中的总汞浓度在冬季均很低;然而在春季,受大气汞亏损事件(Atmospheric Mercury Depletion Events, AMDEs)的影响,南北极雪冰中汞的浓度均显著增加[111-119].早期研究表明积雪中甲基汞与大气沉降密切相关,但在AMDEs期间甲基汞与无机汞表现出完全不同的行为模式[108,111].Constant等[120]调查了加拿大亚北极地区AMDEs时期雪冰中的甲基汞变化规律,结果表明甲基汞的来源与AMDEs无关.雪冰甲基汞与氯化物之间存在较强的相关性,表明沿海地区积雪甲基汞可能来自海盐气溶胶[108].海洋中浮游植物可以产生二甲基汞(DMeHg),由于二甲基汞具有较高的挥发性,它可以从海洋释放到大气中,被自由基氧化成单甲基汞,然后随海盐沉降到附近的雪面之上[108,121-123].值得注意的是新沉降的甲基汞在积雪中并不稳定,15%~56%的甲基汞会在夜间通过去甲基化或以其他方式损失[124]. ...
... 积雪中甲基汞的来源在过去被归因于大气沉降,然而随着研究的不断深入,新的证据表明在低纬度苔原的积雪中可能存在着较强的汞甲基化活动[108,111,123-127].在融雪期间,雪冰融水中甲基汞占总汞的比例增大(从2.7%增加到7.6%)[126].导致这一现象的原因一方面由于甲基汞从无机/有机物中浸出,在雪冰融化时趋于集中;另一方面是由于雪冰内部可能发生甲基化作用,微生物活动和反应可能在积雪中产生了甲基汞[108,120,123-127].极地积雪的pH值通常为酸性,有机物以及微生物等存在于雪层中,因此在消融的积雪中存在微生物汞甲基化的可能[120].随着气候变暖,温度升高会促使积雪中微生物种群数量增长,从而导致微生物影响汞甲基化的强度大幅增加[25].Loseto等[60]估算出北极地区积雪中甲基汞产量约为1.5 mg·km-2·d-1,并指出北极康沃利斯岛陆地积雪融化是地表径流中最重要的甲基汞来源. ...
... ,123-127].极地积雪的pH值通常为酸性,有机物以及微生物等存在于雪层中,因此在消融的积雪中存在微生物汞甲基化的可能[120].随着气候变暖,温度升高会促使积雪中微生物种群数量增长,从而导致微生物影响汞甲基化的强度大幅增加[25].Loseto等[60]估算出北极地区积雪中甲基汞产量约为1.5 mg·km-2·d-1,并指出北极康沃利斯岛陆地积雪融化是地表径流中最重要的甲基汞来源. ...
... Concentrations of MeHg in the cryospheric environment
Table 2| 地点 | 样品类型 | 甲基汞 | 总汞 | 文献来源 |
|---|
| Devon Island, Canada | 多年冻土 | 0.3~3.1 ng·g-1 | — | [58] |
| Barrow, Alaska | 热融湖塘 | 0.3~2.3 ng·L-1 | 25.3~141.2 ng·L-1 | [23] |
| Tibetan Plateau, China | 多年冻土 | — | 5.6~39.3 ng·g-1 | [17] |
| Fennoscandia | 多年冻土 | 0.004~28 ng·g-1 | 1.1~210 ng·g-1 | [24] |
| Tussock tundra, Alaska | 多年冻土 | — | 56~226 ng·g-1 | [16] |
| Laohugou No.12, Tibetan Plateau, China | 雪冰 | 0.005~0.5 ng·L-1 | 24.4~172.1 ng·L-1 | [56] |
| Tibetan Plateau, China | 冰尘 | 0.4~1.7 ng·g-1 | 17.9~114.5 ng·g-1 | [19] |
| French Alps | 表层雪 | 0.002~0.3 ng·L-1 | 1.8~169.5 ng·L-1 | [57] |
| Spitsbergen Island, Norway | 表层雪 | 0.004~0.5 ng·L-1 | 1~90 ng·L-1 | [123] |
| Resolute Bay, High Arctic, Canada | 表层雪 | <0.015~0.2 ng·L-1 | 2.6~150 ng·L-1 | [125] |
| Hudson Bay, Canada | 表层雪 | 0.04~0.14 ng·L-1 | 1.6~5.1 ng·L-1 | [120] |
| Alberta, Canada | 表层雪 | 0.02~0.3 ng·L-1 | 0.8~9.0 ng·L-1 | [137] |
| Antarctic | 积雪 | <0.02~0.3 ng·L-1 | 1.0~30.6 ng·L-1 | [59] |
| McMurdo/Ross Island, Antarctic | 积雪 | — | 40~430 ng·L-1 | [114] |
| Casey Station, Antarctic | 海冰 | <0.009~0.09 ng·L-1 | 0.2~5.7 ng·L-1 | [129] |
| Antarctic | 海冰 | <0.02~0.2 ng·L-1 | 1.4~179 ng·L-1 | [59] |
| Beaufort Sea and McClure Strait, Arctic | 海冰 | <0.02~0.5 ng·L-1 | 0.1~12.2 ng·L-1 | [28] |
| Southern Ocean | 海水 | <0.02~0.1 ng·L-1 | 0. 2~23 ng·L-1 | [59] |
| Arctic Ocean | 海水 | 0.006~0.07 ng·L-1 | 0.09~1.4 ng·L-1 | [135] |
| Resolute Bay, High Arctic, Canada | 海水 | 0.06~0.1 ng·L-1 | 0.1~0.2 ng·L-1 | [125] |
注:表格中海水均为海冰覆盖水域. ...
Photodecomposition of methylmercury in an Arctic Alaskan lake
1
2006
... 降雪是冰冻圈大气降水最重要的表现形式,是大气汞进入冰冻圈的重要环节[108].积雪亦是一种活跃的媒介,消融后将成为环境汞的重要来源[60,109-110].在南北极地区,南极(0.1~1 ng·L-1)与北极(1~5 ng·L-1)表层雪冰中的总汞浓度在冬季均很低;然而在春季,受大气汞亏损事件(Atmospheric Mercury Depletion Events, AMDEs)的影响,南北极雪冰中汞的浓度均显著增加[111-119].早期研究表明积雪中甲基汞与大气沉降密切相关,但在AMDEs期间甲基汞与无机汞表现出完全不同的行为模式[108,111].Constant等[120]调查了加拿大亚北极地区AMDEs时期雪冰中的甲基汞变化规律,结果表明甲基汞的来源与AMDEs无关.雪冰甲基汞与氯化物之间存在较强的相关性,表明沿海地区积雪甲基汞可能来自海盐气溶胶[108].海洋中浮游植物可以产生二甲基汞(DMeHg),由于二甲基汞具有较高的挥发性,它可以从海洋释放到大气中,被自由基氧化成单甲基汞,然后随海盐沉降到附近的雪面之上[108,121-123].值得注意的是新沉降的甲基汞在积雪中并不稳定,15%~56%的甲基汞会在夜间通过去甲基化或以其他方式损失[124]. ...
Methylated mercury species in Canadian high Arctic marine surface waters and snowpacks
2
2007
... Concentrations of MeHg in the cryospheric environment
Table 2| 地点 | 样品类型 | 甲基汞 | 总汞 | 文献来源 |
|---|
| Devon Island, Canada | 多年冻土 | 0.3~3.1 ng·g-1 | — | [58] |
| Barrow, Alaska | 热融湖塘 | 0.3~2.3 ng·L-1 | 25.3~141.2 ng·L-1 | [23] |
| Tibetan Plateau, China | 多年冻土 | — | 5.6~39.3 ng·g-1 | [17] |
| Fennoscandia | 多年冻土 | 0.004~28 ng·g-1 | 1.1~210 ng·g-1 | [24] |
| Tussock tundra, Alaska | 多年冻土 | — | 56~226 ng·g-1 | [16] |
| Laohugou No.12, Tibetan Plateau, China | 雪冰 | 0.005~0.5 ng·L-1 | 24.4~172.1 ng·L-1 | [56] |
| Tibetan Plateau, China | 冰尘 | 0.4~1.7 ng·g-1 | 17.9~114.5 ng·g-1 | [19] |
| French Alps | 表层雪 | 0.002~0.3 ng·L-1 | 1.8~169.5 ng·L-1 | [57] |
| Spitsbergen Island, Norway | 表层雪 | 0.004~0.5 ng·L-1 | 1~90 ng·L-1 | [123] |
| Resolute Bay, High Arctic, Canada | 表层雪 | <0.015~0.2 ng·L-1 | 2.6~150 ng·L-1 | [125] |
| Hudson Bay, Canada | 表层雪 | 0.04~0.14 ng·L-1 | 1.6~5.1 ng·L-1 | [120] |
| Alberta, Canada | 表层雪 | 0.02~0.3 ng·L-1 | 0.8~9.0 ng·L-1 | [137] |
| Antarctic | 积雪 | <0.02~0.3 ng·L-1 | 1.0~30.6 ng·L-1 | [59] |
| McMurdo/Ross Island, Antarctic | 积雪 | — | 40~430 ng·L-1 | [114] |
| Casey Station, Antarctic | 海冰 | <0.009~0.09 ng·L-1 | 0.2~5.7 ng·L-1 | [129] |
| Antarctic | 海冰 | <0.02~0.2 ng·L-1 | 1.4~179 ng·L-1 | [59] |
| Beaufort Sea and McClure Strait, Arctic | 海冰 | <0.02~0.5 ng·L-1 | 0.1~12.2 ng·L-1 | [28] |
| Southern Ocean | 海水 | <0.02~0.1 ng·L-1 | 0. 2~23 ng·L-1 | [59] |
| Arctic Ocean | 海水 | 0.006~0.07 ng·L-1 | 0.09~1.4 ng·L-1 | [135] |
| Resolute Bay, High Arctic, Canada | 海水 | 0.06~0.1 ng·L-1 | 0.1~0.2 ng·L-1 | [125] |
注:表格中海水均为海冰覆盖水域. ...
... [
125]
注:表格中海水均为海冰覆盖水域. ...
Total mercury and methylmercury dynamics in upland–peatland watersheds during snowmelt
1
2008
... 积雪中甲基汞的来源在过去被归因于大气沉降,然而随着研究的不断深入,新的证据表明在低纬度苔原的积雪中可能存在着较强的汞甲基化活动[108,111,123-127].在融雪期间,雪冰融水中甲基汞占总汞的比例增大(从2.7%增加到7.6%)[126].导致这一现象的原因一方面由于甲基汞从无机/有机物中浸出,在雪冰融化时趋于集中;另一方面是由于雪冰内部可能发生甲基化作用,微生物活动和反应可能在积雪中产生了甲基汞[108,120,123-127].极地积雪的pH值通常为酸性,有机物以及微生物等存在于雪层中,因此在消融的积雪中存在微生物汞甲基化的可能[120].随着气候变暖,温度升高会促使积雪中微生物种群数量增长,从而导致微生物影响汞甲基化的强度大幅增加[25].Loseto等[60]估算出北极地区积雪中甲基汞产量约为1.5 mg·km-2·d-1,并指出北极康沃利斯岛陆地积雪融化是地表径流中最重要的甲基汞来源. ...
A mass budget for mercury and methylmercury in the Arctic Ocean
2
2016
... 积雪中甲基汞的来源在过去被归因于大气沉降,然而随着研究的不断深入,新的证据表明在低纬度苔原的积雪中可能存在着较强的汞甲基化活动[108,111,123-127].在融雪期间,雪冰融水中甲基汞占总汞的比例增大(从2.7%增加到7.6%)[126].导致这一现象的原因一方面由于甲基汞从无机/有机物中浸出,在雪冰融化时趋于集中;另一方面是由于雪冰内部可能发生甲基化作用,微生物活动和反应可能在积雪中产生了甲基汞[108,120,123-127].极地积雪的pH值通常为酸性,有机物以及微生物等存在于雪层中,因此在消融的积雪中存在微生物汞甲基化的可能[120].随着气候变暖,温度升高会促使积雪中微生物种群数量增长,从而导致微生物影响汞甲基化的强度大幅增加[25].Loseto等[60]估算出北极地区积雪中甲基汞产量约为1.5 mg·km-2·d-1,并指出北极康沃利斯岛陆地积雪融化是地表径流中最重要的甲基汞来源. ...
... -127].极地积雪的pH值通常为酸性,有机物以及微生物等存在于雪层中,因此在消融的积雪中存在微生物汞甲基化的可能[120].随着气候变暖,温度升高会促使积雪中微生物种群数量增长,从而导致微生物影响汞甲基化的强度大幅增加[25].Loseto等[60]估算出北极地区积雪中甲基汞产量约为1.5 mg·km-2·d-1,并指出北极康沃利斯岛陆地积雪融化是地表径流中最重要的甲基汞来源. ...
Determination of monomethylmercury and dimethylmercury in the Arctic marine boundary layer
1
2015
... 海冰中的甲基汞来源主要包括大气沉降和原位汞甲基化.海洋中的浮游植物生产的二甲基汞从海面释放,在大气中降解为甲基汞,最后又沉降到海冰或海水中,这是海冰甲基汞重要来源之一[121].Baya等[128]研究指出海冰覆盖和初级生产是促进海洋浮游植物生产二甲基汞的主要驱动因素.海冰甲基汞来源除了大气和海洋之外,海冰原位产甲基汞亦非常重要.大量研究已表明海冰中下部和冰水界面是已知的可能产生甲基汞的区位[28,129].因此,海冰中汞的原位甲基化使得海冰融化极有可能成为海洋生态系统新的甲基汞来源. ...
Mercury in the Southern Ocean
4
2011
... 海冰中的甲基汞来源主要包括大气沉降和原位汞甲基化.海洋中的浮游植物生产的二甲基汞从海面释放,在大气中降解为甲基汞,最后又沉降到海冰或海水中,这是海冰甲基汞重要来源之一[121].Baya等[128]研究指出海冰覆盖和初级生产是促进海洋浮游植物生产二甲基汞的主要驱动因素.海冰甲基汞来源除了大气和海洋之外,海冰原位产甲基汞亦非常重要.大量研究已表明海冰中下部和冰水界面是已知的可能产生甲基汞的区位[28,129].因此,海冰中汞的原位甲基化使得海冰融化极有可能成为海洋生态系统新的甲基汞来源. ...
... Cossa等[129]在南极地区开展了“积雪-海冰-卤水-海水”连续系统中的汞循环研究.结果表明,尽管海冰与海水的甲基汞浓度范围大致相同,但在整个系统中分布较为复杂,海冰底部的甲基汞浓度达到峰值.Beattie等[28]在北极海冰中亦观测到了类似的分布模式,海冰中下部甲基汞与总汞比值通常非常高.相比底层水体,海冰具有更高的微生物活性、丰度和多样性[130-133].大气汞通过干湿沉降的方式积累在海冰和海水中为微生物甲基化提供了底物[134-135],并且海冰内部存在局部厌氧条件,成为硫酸盐还原菌和铁还原菌繁衍的氧化还原区,这种现象非常有利于海冰汞甲基化的发生[131].此外海冰-海水界面通常具有丰富的藻类、叶绿素和有机质[132-133].在春季和夏季,由于冰雪融化和浮游植物生长,无机汞被浮游植物吸收利用,这些过程极有可能在海冰-海水界面发生甲基化[133-134].已有研究表明,在南极和北极海冰底部都观测到较高水平的甲基汞含量,这与原位产生甲基汞密切相关,因为海冰底部也有较高的细菌丰度和有机质含量,而且厌氧环境亦有利于汞甲基化细菌的生长繁殖[28,129]. ...
... ,129]. ...
... Concentrations of MeHg in the cryospheric environment
Table 2| 地点 | 样品类型 | 甲基汞 | 总汞 | 文献来源 |
|---|
| Devon Island, Canada | 多年冻土 | 0.3~3.1 ng·g-1 | — | [58] |
| Barrow, Alaska | 热融湖塘 | 0.3~2.3 ng·L-1 | 25.3~141.2 ng·L-1 | [23] |
| Tibetan Plateau, China | 多年冻土 | — | 5.6~39.3 ng·g-1 | [17] |
| Fennoscandia | 多年冻土 | 0.004~28 ng·g-1 | 1.1~210 ng·g-1 | [24] |
| Tussock tundra, Alaska | 多年冻土 | — | 56~226 ng·g-1 | [16] |
| Laohugou No.12, Tibetan Plateau, China | 雪冰 | 0.005~0.5 ng·L-1 | 24.4~172.1 ng·L-1 | [56] |
| Tibetan Plateau, China | 冰尘 | 0.4~1.7 ng·g-1 | 17.9~114.5 ng·g-1 | [19] |
| French Alps | 表层雪 | 0.002~0.3 ng·L-1 | 1.8~169.5 ng·L-1 | [57] |
| Spitsbergen Island, Norway | 表层雪 | 0.004~0.5 ng·L-1 | 1~90 ng·L-1 | [123] |
| Resolute Bay, High Arctic, Canada | 表层雪 | <0.015~0.2 ng·L-1 | 2.6~150 ng·L-1 | [125] |
| Hudson Bay, Canada | 表层雪 | 0.04~0.14 ng·L-1 | 1.6~5.1 ng·L-1 | [120] |
| Alberta, Canada | 表层雪 | 0.02~0.3 ng·L-1 | 0.8~9.0 ng·L-1 | [137] |
| Antarctic | 积雪 | <0.02~0.3 ng·L-1 | 1.0~30.6 ng·L-1 | [59] |
| McMurdo/Ross Island, Antarctic | 积雪 | — | 40~430 ng·L-1 | [114] |
| Casey Station, Antarctic | 海冰 | <0.009~0.09 ng·L-1 | 0.2~5.7 ng·L-1 | [129] |
| Antarctic | 海冰 | <0.02~0.2 ng·L-1 | 1.4~179 ng·L-1 | [59] |
| Beaufort Sea and McClure Strait, Arctic | 海冰 | <0.02~0.5 ng·L-1 | 0.1~12.2 ng·L-1 | [28] |
| Southern Ocean | 海水 | <0.02~0.1 ng·L-1 | 0. 2~23 ng·L-1 | [59] |
| Arctic Ocean | 海水 | 0.006~0.07 ng·L-1 | 0.09~1.4 ng·L-1 | [135] |
| Resolute Bay, High Arctic, Canada | 海水 | 0.06~0.1 ng·L-1 | 0.1~0.2 ng·L-1 | [125] |
注:表格中海水均为海冰覆盖水域. ...
Respiration and bacterial carbon dynamics in Arctic sea ice
1
2011
... Cossa等[129]在南极地区开展了“积雪-海冰-卤水-海水”连续系统中的汞循环研究.结果表明,尽管海冰与海水的甲基汞浓度范围大致相同,但在整个系统中分布较为复杂,海冰底部的甲基汞浓度达到峰值.Beattie等[28]在北极海冰中亦观测到了类似的分布模式,海冰中下部甲基汞与总汞比值通常非常高.相比底层水体,海冰具有更高的微生物活性、丰度和多样性[130-133].大气汞通过干湿沉降的方式积累在海冰和海水中为微生物甲基化提供了底物[134-135],并且海冰内部存在局部厌氧条件,成为硫酸盐还原菌和铁还原菌繁衍的氧化还原区,这种现象非常有利于海冰汞甲基化的发生[131].此外海冰-海水界面通常具有丰富的藻类、叶绿素和有机质[132-133].在春季和夏季,由于冰雪融化和浮游植物生长,无机汞被浮游植物吸收利用,这些过程极有可能在海冰-海水界面发生甲基化[133-134].已有研究表明,在南极和北极海冰底部都观测到较高水平的甲基汞含量,这与原位产生甲基汞密切相关,因为海冰底部也有较高的细菌丰度和有机质含量,而且厌氧环境亦有利于汞甲基化细菌的生长繁殖[28,129]. ...
Denitrification activity and oxygen dynamics in Arctic sea ice
1
2008
... Cossa等[129]在南极地区开展了“积雪-海冰-卤水-海水”连续系统中的汞循环研究.结果表明,尽管海冰与海水的甲基汞浓度范围大致相同,但在整个系统中分布较为复杂,海冰底部的甲基汞浓度达到峰值.Beattie等[28]在北极海冰中亦观测到了类似的分布模式,海冰中下部甲基汞与总汞比值通常非常高.相比底层水体,海冰具有更高的微生物活性、丰度和多样性[130-133].大气汞通过干湿沉降的方式积累在海冰和海水中为微生物甲基化提供了底物[134-135],并且海冰内部存在局部厌氧条件,成为硫酸盐还原菌和铁还原菌繁衍的氧化还原区,这种现象非常有利于海冰汞甲基化的发生[131].此外海冰-海水界面通常具有丰富的藻类、叶绿素和有机质[132-133].在春季和夏季,由于冰雪融化和浮游植物生长,无机汞被浮游植物吸收利用,这些过程极有可能在海冰-海水界面发生甲基化[133-134].已有研究表明,在南极和北极海冰底部都观测到较高水平的甲基汞含量,这与原位产生甲基汞密切相关,因为海冰底部也有较高的细菌丰度和有机质含量,而且厌氧环境亦有利于汞甲基化细菌的生长繁殖[28,129]. ...
Leads in Arctic pack ice enable early phytoplankton blooms below snow-covered sea ice
1
2017
... Cossa等[129]在南极地区开展了“积雪-海冰-卤水-海水”连续系统中的汞循环研究.结果表明,尽管海冰与海水的甲基汞浓度范围大致相同,但在整个系统中分布较为复杂,海冰底部的甲基汞浓度达到峰值.Beattie等[28]在北极海冰中亦观测到了类似的分布模式,海冰中下部甲基汞与总汞比值通常非常高.相比底层水体,海冰具有更高的微生物活性、丰度和多样性[130-133].大气汞通过干湿沉降的方式积累在海冰和海水中为微生物甲基化提供了底物[134-135],并且海冰内部存在局部厌氧条件,成为硫酸盐还原菌和铁还原菌繁衍的氧化还原区,这种现象非常有利于海冰汞甲基化的发生[131].此外海冰-海水界面通常具有丰富的藻类、叶绿素和有机质[132-133].在春季和夏季,由于冰雪融化和浮游植物生长,无机汞被浮游植物吸收利用,这些过程极有可能在海冰-海水界面发生甲基化[133-134].已有研究表明,在南极和北极海冰底部都观测到较高水平的甲基汞含量,这与原位产生甲基汞密切相关,因为海冰底部也有较高的细菌丰度和有机质含量,而且厌氧环境亦有利于汞甲基化细菌的生长繁殖[28,129]. ...
Distributions of phytoplankton blooms in the Southern Ocean
3
1993
... Cossa等[129]在南极地区开展了“积雪-海冰-卤水-海水”连续系统中的汞循环研究.结果表明,尽管海冰与海水的甲基汞浓度范围大致相同,但在整个系统中分布较为复杂,海冰底部的甲基汞浓度达到峰值.Beattie等[28]在北极海冰中亦观测到了类似的分布模式,海冰中下部甲基汞与总汞比值通常非常高.相比底层水体,海冰具有更高的微生物活性、丰度和多样性[130-133].大气汞通过干湿沉降的方式积累在海冰和海水中为微生物甲基化提供了底物[134-135],并且海冰内部存在局部厌氧条件,成为硫酸盐还原菌和铁还原菌繁衍的氧化还原区,这种现象非常有利于海冰汞甲基化的发生[131].此外海冰-海水界面通常具有丰富的藻类、叶绿素和有机质[132-133].在春季和夏季,由于冰雪融化和浮游植物生长,无机汞被浮游植物吸收利用,这些过程极有可能在海冰-海水界面发生甲基化[133-134].已有研究表明,在南极和北极海冰底部都观测到较高水平的甲基汞含量,这与原位产生甲基汞密切相关,因为海冰底部也有较高的细菌丰度和有机质含量,而且厌氧环境亦有利于汞甲基化细菌的生长繁殖[28,129]. ...
... -133].在春季和夏季,由于冰雪融化和浮游植物生长,无机汞被浮游植物吸收利用,这些过程极有可能在海冰-海水界面发生甲基化[133-134].已有研究表明,在南极和北极海冰底部都观测到较高水平的甲基汞含量,这与原位产生甲基汞密切相关,因为海冰底部也有较高的细菌丰度和有机质含量,而且厌氧环境亦有利于汞甲基化细菌的生长繁殖[28,129]. ...
... [133-134].已有研究表明,在南极和北极海冰底部都观测到较高水平的甲基汞含量,这与原位产生甲基汞密切相关,因为海冰底部也有较高的细菌丰度和有机质含量,而且厌氧环境亦有利于汞甲基化细菌的生长繁殖[28,129]. ...
Biogeochemistry: mercury methylation on ice
4
2016
... Cossa等[129]在南极地区开展了“积雪-海冰-卤水-海水”连续系统中的汞循环研究.结果表明,尽管海冰与海水的甲基汞浓度范围大致相同,但在整个系统中分布较为复杂,海冰底部的甲基汞浓度达到峰值.Beattie等[28]在北极海冰中亦观测到了类似的分布模式,海冰中下部甲基汞与总汞比值通常非常高.相比底层水体,海冰具有更高的微生物活性、丰度和多样性[130-133].大气汞通过干湿沉降的方式积累在海冰和海水中为微生物甲基化提供了底物[134-135],并且海冰内部存在局部厌氧条件,成为硫酸盐还原菌和铁还原菌繁衍的氧化还原区,这种现象非常有利于海冰汞甲基化的发生[131].此外海冰-海水界面通常具有丰富的藻类、叶绿素和有机质[132-133].在春季和夏季,由于冰雪融化和浮游植物生长,无机汞被浮游植物吸收利用,这些过程极有可能在海冰-海水界面发生甲基化[133-134].已有研究表明,在南极和北极海冰底部都观测到较高水平的甲基汞含量,这与原位产生甲基汞密切相关,因为海冰底部也有较高的细菌丰度和有机质含量,而且厌氧环境亦有利于汞甲基化细菌的生长繁殖[28,129]. ...
... -134].已有研究表明,在南极和北极海冰底部都观测到较高水平的甲基汞含量,这与原位产生甲基汞密切相关,因为海冰底部也有较高的细菌丰度和有机质含量,而且厌氧环境亦有利于汞甲基化细菌的生长繁殖[28,129]. ...
... 最新的研究不断揭示出潜在的新型海洋汞甲基化微生物,它们具有更强的耐氧性和更广泛的生境范围,这些都是有利于海冰汞甲基化的理想条件[59].Gionfriddo等[59]通过对南极海冰汞甲基化研究表明,南极海冰中存在甲基汞的富集现象,主要是由微嗜氧细菌-硝化刺菌属(Nitrospina)主导产生的,参与调控微生物汞甲基化的基因为hgcA-like.这与之前的认知有所不同,过去认为只有厌氧条件支持显著的汞甲基化过程,且汞甲基化微生物都属于厌氧细菌.因此,海冰汞的研究不仅揭示了更多潜在的新型海洋汞甲基化微生物,而且极大地拓宽了我们对汞甲基化微生物耐氧性的认识.气候变暖背景下,海冰变薄、季节性海冰区扩大、表层海洋分层加剧和浮游生物生态动力学的变化,这些过程都会造成海洋甲基汞产量增加[26,136].从海冰融化释放出来的大量甲基汞进入海洋生态系统之后会在食物链中富集和放大,将会对南北极水生食物链中高营养级的动物构成很大威胁[26,28,59,134].通过对北极多年海冰的融化情况的估算,每年输入北冰洋的总汞和甲基汞的通量估计分别为420 kg·a-1和42 kg·a-1[28].南极海冰融化亦会给海洋分别带来超过9 600 kg·a-1总汞和18 kg·a-1甲基汞,总汞的估算值要比北冰洋高出一个数量级[59,134].为方便后续研究,将冰冻圈总汞和甲基汞浓度水平相关信息汇总于表2. ...
... ,134].为方便后续研究,将冰冻圈总汞和甲基汞浓度水平相关信息汇总于表2. ...
Shallow methylmercury production in the marginal sea ice zone of the central Arctic Ocean
2
2015
... Cossa等[129]在南极地区开展了“积雪-海冰-卤水-海水”连续系统中的汞循环研究.结果表明,尽管海冰与海水的甲基汞浓度范围大致相同,但在整个系统中分布较为复杂,海冰底部的甲基汞浓度达到峰值.Beattie等[28]在北极海冰中亦观测到了类似的分布模式,海冰中下部甲基汞与总汞比值通常非常高.相比底层水体,海冰具有更高的微生物活性、丰度和多样性[130-133].大气汞通过干湿沉降的方式积累在海冰和海水中为微生物甲基化提供了底物[134-135],并且海冰内部存在局部厌氧条件,成为硫酸盐还原菌和铁还原菌繁衍的氧化还原区,这种现象非常有利于海冰汞甲基化的发生[131].此外海冰-海水界面通常具有丰富的藻类、叶绿素和有机质[132-133].在春季和夏季,由于冰雪融化和浮游植物生长,无机汞被浮游植物吸收利用,这些过程极有可能在海冰-海水界面发生甲基化[133-134].已有研究表明,在南极和北极海冰底部都观测到较高水平的甲基汞含量,这与原位产生甲基汞密切相关,因为海冰底部也有较高的细菌丰度和有机质含量,而且厌氧环境亦有利于汞甲基化细菌的生长繁殖[28,129]. ...
... Concentrations of MeHg in the cryospheric environment
Table 2| 地点 | 样品类型 | 甲基汞 | 总汞 | 文献来源 |
|---|
| Devon Island, Canada | 多年冻土 | 0.3~3.1 ng·g-1 | — | [58] |
| Barrow, Alaska | 热融湖塘 | 0.3~2.3 ng·L-1 | 25.3~141.2 ng·L-1 | [23] |
| Tibetan Plateau, China | 多年冻土 | — | 5.6~39.3 ng·g-1 | [17] |
| Fennoscandia | 多年冻土 | 0.004~28 ng·g-1 | 1.1~210 ng·g-1 | [24] |
| Tussock tundra, Alaska | 多年冻土 | — | 56~226 ng·g-1 | [16] |
| Laohugou No.12, Tibetan Plateau, China | 雪冰 | 0.005~0.5 ng·L-1 | 24.4~172.1 ng·L-1 | [56] |
| Tibetan Plateau, China | 冰尘 | 0.4~1.7 ng·g-1 | 17.9~114.5 ng·g-1 | [19] |
| French Alps | 表层雪 | 0.002~0.3 ng·L-1 | 1.8~169.5 ng·L-1 | [57] |
| Spitsbergen Island, Norway | 表层雪 | 0.004~0.5 ng·L-1 | 1~90 ng·L-1 | [123] |
| Resolute Bay, High Arctic, Canada | 表层雪 | <0.015~0.2 ng·L-1 | 2.6~150 ng·L-1 | [125] |
| Hudson Bay, Canada | 表层雪 | 0.04~0.14 ng·L-1 | 1.6~5.1 ng·L-1 | [120] |
| Alberta, Canada | 表层雪 | 0.02~0.3 ng·L-1 | 0.8~9.0 ng·L-1 | [137] |
| Antarctic | 积雪 | <0.02~0.3 ng·L-1 | 1.0~30.6 ng·L-1 | [59] |
| McMurdo/Ross Island, Antarctic | 积雪 | — | 40~430 ng·L-1 | [114] |
| Casey Station, Antarctic | 海冰 | <0.009~0.09 ng·L-1 | 0.2~5.7 ng·L-1 | [129] |
| Antarctic | 海冰 | <0.02~0.2 ng·L-1 | 1.4~179 ng·L-1 | [59] |
| Beaufort Sea and McClure Strait, Arctic | 海冰 | <0.02~0.5 ng·L-1 | 0.1~12.2 ng·L-1 | [28] |
| Southern Ocean | 海水 | <0.02~0.1 ng·L-1 | 0. 2~23 ng·L-1 | [59] |
| Arctic Ocean | 海水 | 0.006~0.07 ng·L-1 | 0.09~1.4 ng·L-1 | [135] |
| Resolute Bay, High Arctic, Canada | 海水 | 0.06~0.1 ng·L-1 | 0.1~0.2 ng·L-1 | [125] |
注:表格中海水均为海冰覆盖水域. ...
A mass balance inventory of mercury in the Arctic Ocean
1
2008
... 最新的研究不断揭示出潜在的新型海洋汞甲基化微生物,它们具有更强的耐氧性和更广泛的生境范围,这些都是有利于海冰汞甲基化的理想条件[59].Gionfriddo等[59]通过对南极海冰汞甲基化研究表明,南极海冰中存在甲基汞的富集现象,主要是由微嗜氧细菌-硝化刺菌属(Nitrospina)主导产生的,参与调控微生物汞甲基化的基因为hgcA-like.这与之前的认知有所不同,过去认为只有厌氧条件支持显著的汞甲基化过程,且汞甲基化微生物都属于厌氧细菌.因此,海冰汞的研究不仅揭示了更多潜在的新型海洋汞甲基化微生物,而且极大地拓宽了我们对汞甲基化微生物耐氧性的认识.气候变暖背景下,海冰变薄、季节性海冰区扩大、表层海洋分层加剧和浮游生物生态动力学的变化,这些过程都会造成海洋甲基汞产量增加[26,136].从海冰融化释放出来的大量甲基汞进入海洋生态系统之后会在食物链中富集和放大,将会对南北极水生食物链中高营养级的动物构成很大威胁[26,28,59,134].通过对北极多年海冰的融化情况的估算,每年输入北冰洋的总汞和甲基汞的通量估计分别为420 kg·a-1和42 kg·a-1[28].南极海冰融化亦会给海洋分别带来超过9 600 kg·a-1总汞和18 kg·a-1甲基汞,总汞的估算值要比北冰洋高出一个数量级[59,134].为方便后续研究,将冰冻圈总汞和甲基汞浓度水平相关信息汇总于表2. ...
Tailings ponds of the Athabasca Oil Sands Region, Alberta, Canada, are likely not significant sources of total mercury and methylmercury to nearby ground and surface waters
1
2019
... Concentrations of MeHg in the cryospheric environment
Table 2| 地点 | 样品类型 | 甲基汞 | 总汞 | 文献来源 |
|---|
| Devon Island, Canada | 多年冻土 | 0.3~3.1 ng·g-1 | — | [58] |
| Barrow, Alaska | 热融湖塘 | 0.3~2.3 ng·L-1 | 25.3~141.2 ng·L-1 | [23] |
| Tibetan Plateau, China | 多年冻土 | — | 5.6~39.3 ng·g-1 | [17] |
| Fennoscandia | 多年冻土 | 0.004~28 ng·g-1 | 1.1~210 ng·g-1 | [24] |
| Tussock tundra, Alaska | 多年冻土 | — | 56~226 ng·g-1 | [16] |
| Laohugou No.12, Tibetan Plateau, China | 雪冰 | 0.005~0.5 ng·L-1 | 24.4~172.1 ng·L-1 | [56] |
| Tibetan Plateau, China | 冰尘 | 0.4~1.7 ng·g-1 | 17.9~114.5 ng·g-1 | [19] |
| French Alps | 表层雪 | 0.002~0.3 ng·L-1 | 1.8~169.5 ng·L-1 | [57] |
| Spitsbergen Island, Norway | 表层雪 | 0.004~0.5 ng·L-1 | 1~90 ng·L-1 | [123] |
| Resolute Bay, High Arctic, Canada | 表层雪 | <0.015~0.2 ng·L-1 | 2.6~150 ng·L-1 | [125] |
| Hudson Bay, Canada | 表层雪 | 0.04~0.14 ng·L-1 | 1.6~5.1 ng·L-1 | [120] |
| Alberta, Canada | 表层雪 | 0.02~0.3 ng·L-1 | 0.8~9.0 ng·L-1 | [137] |
| Antarctic | 积雪 | <0.02~0.3 ng·L-1 | 1.0~30.6 ng·L-1 | [59] |
| McMurdo/Ross Island, Antarctic | 积雪 | — | 40~430 ng·L-1 | [114] |
| Casey Station, Antarctic | 海冰 | <0.009~0.09 ng·L-1 | 0.2~5.7 ng·L-1 | [129] |
| Antarctic | 海冰 | <0.02~0.2 ng·L-1 | 1.4~179 ng·L-1 | [59] |
| Beaufort Sea and McClure Strait, Arctic | 海冰 | <0.02~0.5 ng·L-1 | 0.1~12.2 ng·L-1 | [28] |
| Southern Ocean | 海水 | <0.02~0.1 ng·L-1 | 0. 2~23 ng·L-1 | [59] |
| Arctic Ocean | 海水 | 0.006~0.07 ng·L-1 | 0.09~1.4 ng·L-1 | [135] |
| Resolute Bay, High Arctic, Canada | 海水 | 0.06~0.1 ng·L-1 | 0.1~0.2 ng·L-1 | [125] |
注:表格中海水均为海冰覆盖水域. ...
Psychrophilic lifestyles: mechanisms of adaptation and biotechnological tools
1
2019
... 自工业革命以来,人类活动已深刻改变了全球汞循环过程,造成大量人为汞污染物进入冰冻圈.近期研究发现,嗜冷菌微生物已经进化出了特殊机制来克服低温环境,并积极参与到冰冻圈汞甲基化过程,将冰冻圈自然环境中无机汞转化为甲基汞[34,138-139].由于生物富集和累积作用,海豹、鲸鱼、海鸟及北极熊体内的汞含量非常高,这些处于食物链高层的捕食者体内的汞几乎完全是甲基汞形式[140-152].在北极生物群落中海鸟作为顶级捕食者,寿命长、分布广泛而且对汞吸收很敏感,因此海鸟经常被用作海洋生态系统中汞富集研究的生物指示剂[145].研究发现海鸟羽毛中的汞浓度已超过5 μg·g-1(dw)的毒性阈值,而这种高水平浓度通常被认为有害健康和影响生殖[144-145,153-154].南极磷虾是污染物向上层营养级生物转移的重要环节,磷虾体内总汞浓度范围为4.04~19.4 ng·g-1,甲基汞浓度范围为0.26~2.94 ng·g-1,依赖磷虾的消费者(如幼鱼)通过食物链大量富集和积累甲基汞[146-148].已有研究报道南极海洋食物链顶端的物种(如企鹅和哺乳动物)体内甲基汞富集系数高达多个数量级,在北极地区捕食性鱼类中甲基汞含量甚至比海水高出约一百万倍[149-150]. ...
Microbial ecology of the cryosphere (glacial and permafrost habitats): current knowledge
1
2019
... 自工业革命以来,人类活动已深刻改变了全球汞循环过程,造成大量人为汞污染物进入冰冻圈.近期研究发现,嗜冷菌微生物已经进化出了特殊机制来克服低温环境,并积极参与到冰冻圈汞甲基化过程,将冰冻圈自然环境中无机汞转化为甲基汞[34,138-139].由于生物富集和累积作用,海豹、鲸鱼、海鸟及北极熊体内的汞含量非常高,这些处于食物链高层的捕食者体内的汞几乎完全是甲基汞形式[140-152].在北极生物群落中海鸟作为顶级捕食者,寿命长、分布广泛而且对汞吸收很敏感,因此海鸟经常被用作海洋生态系统中汞富集研究的生物指示剂[145].研究发现海鸟羽毛中的汞浓度已超过5 μg·g-1(dw)的毒性阈值,而这种高水平浓度通常被认为有害健康和影响生殖[144-145,153-154].南极磷虾是污染物向上层营养级生物转移的重要环节,磷虾体内总汞浓度范围为4.04~19.4 ng·g-1,甲基汞浓度范围为0.26~2.94 ng·g-1,依赖磷虾的消费者(如幼鱼)通过食物链大量富集和积累甲基汞[146-148].已有研究报道南极海洋食物链顶端的物种(如企鹅和哺乳动物)体内甲基汞富集系数高达多个数量级,在北极地区捕食性鱼类中甲基汞含量甚至比海水高出约一百万倍[149-150]. ...
What are the toxicological effects of mercury in Arctic biota?
1
2013
... 自工业革命以来,人类活动已深刻改变了全球汞循环过程,造成大量人为汞污染物进入冰冻圈.近期研究发现,嗜冷菌微生物已经进化出了特殊机制来克服低温环境,并积极参与到冰冻圈汞甲基化过程,将冰冻圈自然环境中无机汞转化为甲基汞[34,138-139].由于生物富集和累积作用,海豹、鲸鱼、海鸟及北极熊体内的汞含量非常高,这些处于食物链高层的捕食者体内的汞几乎完全是甲基汞形式[140-152].在北极生物群落中海鸟作为顶级捕食者,寿命长、分布广泛而且对汞吸收很敏感,因此海鸟经常被用作海洋生态系统中汞富集研究的生物指示剂[145].研究发现海鸟羽毛中的汞浓度已超过5 μg·g-1(dw)的毒性阈值,而这种高水平浓度通常被认为有害健康和影响生殖[144-145,153-154].南极磷虾是污染物向上层营养级生物转移的重要环节,磷虾体内总汞浓度范围为4.04~19.4 ng·g-1,甲基汞浓度范围为0.26~2.94 ng·g-1,依赖磷虾的消费者(如幼鱼)通过食物链大量富集和积累甲基汞[146-148].已有研究报道南极海洋食物链顶端的物种(如企鹅和哺乳动物)体内甲基汞富集系数高达多个数量级,在北极地区捕食性鱼类中甲基汞含量甚至比海水高出约一百万倍[149-150]. ...
Mercury and other trace elements in a pelagic Arctic marine food web (Northwater Polynya, Baffin Bay)
0
2005
Lead, cadmium, mercury and selenium in Greenland marine animals
0
1996
Human health implications of environmental contaminants in Arctic Canada: a review
0
2005
Mercury hazards to fish, wildlife, and invertebrates: a synoptic review
1
1987
... 自工业革命以来,人类活动已深刻改变了全球汞循环过程,造成大量人为汞污染物进入冰冻圈.近期研究发现,嗜冷菌微生物已经进化出了特殊机制来克服低温环境,并积极参与到冰冻圈汞甲基化过程,将冰冻圈自然环境中无机汞转化为甲基汞[34,138-139].由于生物富集和累积作用,海豹、鲸鱼、海鸟及北极熊体内的汞含量非常高,这些处于食物链高层的捕食者体内的汞几乎完全是甲基汞形式[140-152].在北极生物群落中海鸟作为顶级捕食者,寿命长、分布广泛而且对汞吸收很敏感,因此海鸟经常被用作海洋生态系统中汞富集研究的生物指示剂[145].研究发现海鸟羽毛中的汞浓度已超过5 μg·g-1(dw)的毒性阈值,而这种高水平浓度通常被认为有害健康和影响生殖[144-145,153-154].南极磷虾是污染物向上层营养级生物转移的重要环节,磷虾体内总汞浓度范围为4.04~19.4 ng·g-1,甲基汞浓度范围为0.26~2.94 ng·g-1,依赖磷虾的消费者(如幼鱼)通过食物链大量富集和积累甲基汞[146-148].已有研究报道南极海洋食物链顶端的物种(如企鹅和哺乳动物)体内甲基汞富集系数高达多个数量级,在北极地区捕食性鱼类中甲基汞含量甚至比海水高出约一百万倍[149-150]. ...
Using blood and feathers to investigate large-scale Hg contamination in Arctic seabirds: a review
2
2019
... 自工业革命以来,人类活动已深刻改变了全球汞循环过程,造成大量人为汞污染物进入冰冻圈.近期研究发现,嗜冷菌微生物已经进化出了特殊机制来克服低温环境,并积极参与到冰冻圈汞甲基化过程,将冰冻圈自然环境中无机汞转化为甲基汞[34,138-139].由于生物富集和累积作用,海豹、鲸鱼、海鸟及北极熊体内的汞含量非常高,这些处于食物链高层的捕食者体内的汞几乎完全是甲基汞形式[140-152].在北极生物群落中海鸟作为顶级捕食者,寿命长、分布广泛而且对汞吸收很敏感,因此海鸟经常被用作海洋生态系统中汞富集研究的生物指示剂[145].研究发现海鸟羽毛中的汞浓度已超过5 μg·g-1(dw)的毒性阈值,而这种高水平浓度通常被认为有害健康和影响生殖[144-145,153-154].南极磷虾是污染物向上层营养级生物转移的重要环节,磷虾体内总汞浓度范围为4.04~19.4 ng·g-1,甲基汞浓度范围为0.26~2.94 ng·g-1,依赖磷虾的消费者(如幼鱼)通过食物链大量富集和积累甲基汞[146-148].已有研究报道南极海洋食物链顶端的物种(如企鹅和哺乳动物)体内甲基汞富集系数高达多个数量级,在北极地区捕食性鱼类中甲基汞含量甚至比海水高出约一百万倍[149-150]. ...
... -145,153-154].南极磷虾是污染物向上层营养级生物转移的重要环节,磷虾体内总汞浓度范围为4.04~19.4 ng·g-1,甲基汞浓度范围为0.26~2.94 ng·g-1,依赖磷虾的消费者(如幼鱼)通过食物链大量富集和积累甲基汞[146-148].已有研究报道南极海洋食物链顶端的物种(如企鹅和哺乳动物)体内甲基汞富集系数高达多个数量级,在北极地区捕食性鱼类中甲基汞含量甚至比海水高出约一百万倍[149-150]. ...
Persistent organic pollutants at the base of the Antarctic marine food web
1
2004
... 自工业革命以来,人类活动已深刻改变了全球汞循环过程,造成大量人为汞污染物进入冰冻圈.近期研究发现,嗜冷菌微生物已经进化出了特殊机制来克服低温环境,并积极参与到冰冻圈汞甲基化过程,将冰冻圈自然环境中无机汞转化为甲基汞[34,138-139].由于生物富集和累积作用,海豹、鲸鱼、海鸟及北极熊体内的汞含量非常高,这些处于食物链高层的捕食者体内的汞几乎完全是甲基汞形式[140-152].在北极生物群落中海鸟作为顶级捕食者,寿命长、分布广泛而且对汞吸收很敏感,因此海鸟经常被用作海洋生态系统中汞富集研究的生物指示剂[145].研究发现海鸟羽毛中的汞浓度已超过5 μg·g-1(dw)的毒性阈值,而这种高水平浓度通常被认为有害健康和影响生殖[144-145,153-154].南极磷虾是污染物向上层营养级生物转移的重要环节,磷虾体内总汞浓度范围为4.04~19.4 ng·g-1,甲基汞浓度范围为0.26~2.94 ng·g-1,依赖磷虾的消费者(如幼鱼)通过食物链大量富集和积累甲基汞[146-148].已有研究报道南极海洋食物链顶端的物种(如企鹅和哺乳动物)体内甲基汞富集系数高达多个数量级,在北极地区捕食性鱼类中甲基汞含量甚至比海水高出约一百万倍[149-150]. ...
Patterns of total mercury and methylmercury bioaccumulation in Antarctic krill (Euphausia superba) along the West Antarctic Peninsula
0
2019
Mercury and major essential elements in seals, penguins, and other representative fauna of the Antarctic
1
1993
... 自工业革命以来,人类活动已深刻改变了全球汞循环过程,造成大量人为汞污染物进入冰冻圈.近期研究发现,嗜冷菌微生物已经进化出了特殊机制来克服低温环境,并积极参与到冰冻圈汞甲基化过程,将冰冻圈自然环境中无机汞转化为甲基汞[34,138-139].由于生物富集和累积作用,海豹、鲸鱼、海鸟及北极熊体内的汞含量非常高,这些处于食物链高层的捕食者体内的汞几乎完全是甲基汞形式[140-152].在北极生物群落中海鸟作为顶级捕食者,寿命长、分布广泛而且对汞吸收很敏感,因此海鸟经常被用作海洋生态系统中汞富集研究的生物指示剂[145].研究发现海鸟羽毛中的汞浓度已超过5 μg·g-1(dw)的毒性阈值,而这种高水平浓度通常被认为有害健康和影响生殖[144-145,153-154].南极磷虾是污染物向上层营养级生物转移的重要环节,磷虾体内总汞浓度范围为4.04~19.4 ng·g-1,甲基汞浓度范围为0.26~2.94 ng·g-1,依赖磷虾的消费者(如幼鱼)通过食物链大量富集和积累甲基汞[146-148].已有研究报道南极海洋食物链顶端的物种(如企鹅和哺乳动物)体内甲基汞富集系数高达多个数量级,在北极地区捕食性鱼类中甲基汞含量甚至比海水高出约一百万倍[149-150]. ...
Assumption and elimination of trace elements in Adélie penguins from Antarctica: a preliminary study
1
2002
... 自工业革命以来,人类活动已深刻改变了全球汞循环过程,造成大量人为汞污染物进入冰冻圈.近期研究发现,嗜冷菌微生物已经进化出了特殊机制来克服低温环境,并积极参与到冰冻圈汞甲基化过程,将冰冻圈自然环境中无机汞转化为甲基汞[34,138-139].由于生物富集和累积作用,海豹、鲸鱼、海鸟及北极熊体内的汞含量非常高,这些处于食物链高层的捕食者体内的汞几乎完全是甲基汞形式[140-152].在北极生物群落中海鸟作为顶级捕食者,寿命长、分布广泛而且对汞吸收很敏感,因此海鸟经常被用作海洋生态系统中汞富集研究的生物指示剂[145].研究发现海鸟羽毛中的汞浓度已超过5 μg·g-1(dw)的毒性阈值,而这种高水平浓度通常被认为有害健康和影响生殖[144-145,153-154].南极磷虾是污染物向上层营养级生物转移的重要环节,磷虾体内总汞浓度范围为4.04~19.4 ng·g-1,甲基汞浓度范围为0.26~2.94 ng·g-1,依赖磷虾的消费者(如幼鱼)通过食物链大量富集和积累甲基汞[146-148].已有研究报道南极海洋食物链顶端的物种(如企鹅和哺乳动物)体内甲基汞富集系数高达多个数量级,在北极地区捕食性鱼类中甲基汞含量甚至比海水高出约一百万倍[149-150]. ...
Biomagnification of mercury in an Antarctic marine coastal food web
1
1998
... 自工业革命以来,人类活动已深刻改变了全球汞循环过程,造成大量人为汞污染物进入冰冻圈.近期研究发现,嗜冷菌微生物已经进化出了特殊机制来克服低温环境,并积极参与到冰冻圈汞甲基化过程,将冰冻圈自然环境中无机汞转化为甲基汞[34,138-139].由于生物富集和累积作用,海豹、鲸鱼、海鸟及北极熊体内的汞含量非常高,这些处于食物链高层的捕食者体内的汞几乎完全是甲基汞形式[140-152].在北极生物群落中海鸟作为顶级捕食者,寿命长、分布广泛而且对汞吸收很敏感,因此海鸟经常被用作海洋生态系统中汞富集研究的生物指示剂[145].研究发现海鸟羽毛中的汞浓度已超过5 μg·g-1(dw)的毒性阈值,而这种高水平浓度通常被认为有害健康和影响生殖[144-145,153-154].南极磷虾是污染物向上层营养级生物转移的重要环节,磷虾体内总汞浓度范围为4.04~19.4 ng·g-1,甲基汞浓度范围为0.26~2.94 ng·g-1,依赖磷虾的消费者(如幼鱼)通过食物链大量富集和积累甲基汞[146-148].已有研究报道南极海洋食物链顶端的物种(如企鹅和哺乳动物)体内甲基汞富集系数高达多个数量级,在北极地区捕食性鱼类中甲基汞含量甚至比海水高出约一百万倍[149-150]. ...
Penguins as bioindicators of mercury contamination in the southern Indian Ocean: geographical and temporal trends
0
2016
A review of metal accumulation and toxicity in wild mammals: I. Mercury
1
1986
... 自工业革命以来,人类活动已深刻改变了全球汞循环过程,造成大量人为汞污染物进入冰冻圈.近期研究发现,嗜冷菌微生物已经进化出了特殊机制来克服低温环境,并积极参与到冰冻圈汞甲基化过程,将冰冻圈自然环境中无机汞转化为甲基汞[34,138-139].由于生物富集和累积作用,海豹、鲸鱼、海鸟及北极熊体内的汞含量非常高,这些处于食物链高层的捕食者体内的汞几乎完全是甲基汞形式[140-152].在北极生物群落中海鸟作为顶级捕食者,寿命长、分布广泛而且对汞吸收很敏感,因此海鸟经常被用作海洋生态系统中汞富集研究的生物指示剂[145].研究发现海鸟羽毛中的汞浓度已超过5 μg·g-1(dw)的毒性阈值,而这种高水平浓度通常被认为有害健康和影响生殖[144-145,153-154].南极磷虾是污染物向上层营养级生物转移的重要环节,磷虾体内总汞浓度范围为4.04~19.4 ng·g-1,甲基汞浓度范围为0.26~2.94 ng·g-1,依赖磷虾的消费者(如幼鱼)通过食物链大量富集和积累甲基汞[146-148].已有研究报道南极海洋食物链顶端的物种(如企鹅和哺乳动物)体内甲基汞富集系数高达多个数量级,在北极地区捕食性鱼类中甲基汞含量甚至比海水高出约一百万倍[149-150]. ...
Feather mercury levels in seabirds at South Georgia: influence of trophic position, sex and age
1
2002
... 自工业革命以来,人类活动已深刻改变了全球汞循环过程,造成大量人为汞污染物进入冰冻圈.近期研究发现,嗜冷菌微生物已经进化出了特殊机制来克服低温环境,并积极参与到冰冻圈汞甲基化过程,将冰冻圈自然环境中无机汞转化为甲基汞[34,138-139].由于生物富集和累积作用,海豹、鲸鱼、海鸟及北极熊体内的汞含量非常高,这些处于食物链高层的捕食者体内的汞几乎完全是甲基汞形式[140-152].在北极生物群落中海鸟作为顶级捕食者,寿命长、分布广泛而且对汞吸收很敏感,因此海鸟经常被用作海洋生态系统中汞富集研究的生物指示剂[145].研究发现海鸟羽毛中的汞浓度已超过5 μg·g-1(dw)的毒性阈值,而这种高水平浓度通常被认为有害健康和影响生殖[144-145,153-154].南极磷虾是污染物向上层营养级生物转移的重要环节,磷虾体内总汞浓度范围为4.04~19.4 ng·g-1,甲基汞浓度范围为0.26~2.94 ng·g-1,依赖磷虾的消费者(如幼鱼)通过食物链大量富集和积累甲基汞[146-148].已有研究报道南极海洋食物链顶端的物种(如企鹅和哺乳动物)体内甲基汞富集系数高达多个数量级,在北极地区捕食性鱼类中甲基汞含量甚至比海水高出约一百万倍[149-150]. ...
Seasonal variation of mercury contamination in Arctic seabirds: a pan-arctic assessment
1
2021
... 自工业革命以来,人类活动已深刻改变了全球汞循环过程,造成大量人为汞污染物进入冰冻圈.近期研究发现,嗜冷菌微生物已经进化出了特殊机制来克服低温环境,并积极参与到冰冻圈汞甲基化过程,将冰冻圈自然环境中无机汞转化为甲基汞[34,138-139].由于生物富集和累积作用,海豹、鲸鱼、海鸟及北极熊体内的汞含量非常高,这些处于食物链高层的捕食者体内的汞几乎完全是甲基汞形式[140-152].在北极生物群落中海鸟作为顶级捕食者,寿命长、分布广泛而且对汞吸收很敏感,因此海鸟经常被用作海洋生态系统中汞富集研究的生物指示剂[145].研究发现海鸟羽毛中的汞浓度已超过5 μg·g-1(dw)的毒性阈值,而这种高水平浓度通常被认为有害健康和影响生殖[144-145,153-154].南极磷虾是污染物向上层营养级生物转移的重要环节,磷虾体内总汞浓度范围为4.04~19.4 ng·g-1,甲基汞浓度范围为0.26~2.94 ng·g-1,依赖磷虾的消费者(如幼鱼)通过食物链大量富集和积累甲基汞[146-148].已有研究报道南极海洋食物链顶端的物种(如企鹅和哺乳动物)体内甲基汞富集系数高达多个数量级,在北极地区捕食性鱼类中甲基汞含量甚至比海水高出约一百万倍[149-150]. ...
Methylmercury exposure and adverse cardiovascular effects in Faroese whaling men
1
2009
... 冰冻圈甲基汞富集作用同样对生活在该区域的人类健康构成重大威胁.北极地区居住着土著居民(如因纽特人),他们依赖于捕获鱼类和海洋哺乳动物为食,通过饮食摄入是当地居民暴露甲基汞的主要途径[3,155-157].北极因纽特人原住民由于常年捕食海洋生物,人体内的甲基汞含量都远超过了毒理学阈值[104,158].由于甲基汞对成人和儿童的毒性效应有所不同,处于发育中的儿童中枢神经系统比成年人对甲基汞更为敏感,而孕妇长期处于低剂量甲基汞暴露将会对婴儿的智力发育产生严重影响[33].研究表明,北极一些地区50%以上的母亲和育龄妇女的血液中的汞含量已超过美国环境保护署建议的标准浓度(5.8 μg·L-1),且在格陵兰岛部分地区近乎90%的育龄妇女血液中的汞含量都超标,这些都表明我们迫切需要重视冰冻圈甲基汞暴露风险[33,157-158]. ...
A new era of mercury hazards
0
1998
Toxicological effects of methylmercury
2
C
... 冰冻圈甲基汞富集作用同样对生活在该区域的人类健康构成重大威胁.北极地区居住着土著居民(如因纽特人),他们依赖于捕获鱼类和海洋哺乳动物为食,通过饮食摄入是当地居民暴露甲基汞的主要途径[3,155-157].北极因纽特人原住民由于常年捕食海洋生物,人体内的甲基汞含量都远超过了毒理学阈值[104,158].由于甲基汞对成人和儿童的毒性效应有所不同,处于发育中的儿童中枢神经系统比成年人对甲基汞更为敏感,而孕妇长期处于低剂量甲基汞暴露将会对婴儿的智力发育产生严重影响[33].研究表明,北极一些地区50%以上的母亲和育龄妇女的血液中的汞含量已超过美国环境保护署建议的标准浓度(5.8 μg·L-1),且在格陵兰岛部分地区近乎90%的育龄妇女血液中的汞含量都超标,这些都表明我们迫切需要重视冰冻圈甲基汞暴露风险[33,157-158]. ...
... ,157-158]. ...
Arctic Monitoring and Assessment Programme 2011: mercury in the Arctic
2
2011
... 冰冻圈甲基汞富集作用同样对生活在该区域的人类健康构成重大威胁.北极地区居住着土著居民(如因纽特人),他们依赖于捕获鱼类和海洋哺乳动物为食,通过饮食摄入是当地居民暴露甲基汞的主要途径[3,155-157].北极因纽特人原住民由于常年捕食海洋生物,人体内的甲基汞含量都远超过了毒理学阈值[104,158].由于甲基汞对成人和儿童的毒性效应有所不同,处于发育中的儿童中枢神经系统比成年人对甲基汞更为敏感,而孕妇长期处于低剂量甲基汞暴露将会对婴儿的智力发育产生严重影响[33].研究表明,北极一些地区50%以上的母亲和育龄妇女的血液中的汞含量已超过美国环境保护署建议的标准浓度(5.8 μg·L-1),且在格陵兰岛部分地区近乎90%的育龄妇女血液中的汞含量都超标,这些都表明我们迫切需要重视冰冻圈甲基汞暴露风险[33,157-158]. ...
... -158]. ...


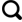



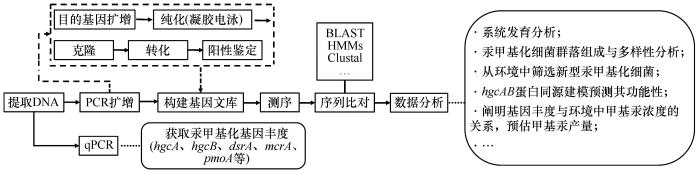

 甘公网安备 62010202000676号
甘公网安备 62010202000676号
{kind=link}
{kind=link}
{kind=link}
{kind=link}