The emerging field of geogenomics: constraining geological problems with genetic data
6
2014
... 系统发育是指某一个生物类群的形成和发展过程[1 -2 ] ,可以通过物种系统发生关系来推测祖先类群形成的时间和分布模式,是现代生物地理学的根基[2 -3 ] .地理分布上截然不同的物种和种群是生物自然演化和响应气候或构造环境变化的结果,其遗传信息往往留有地质历史时期环境变化或构造活动的烙印,因此,重建生物类群的演化历史是回溯过去,认识地质时期环境与构造演化的契机[1 ] .近年来,随着DNA测序技术的发展,已经产生了大量生物类群的遗传数据,并可以预见,这些遗传数据将呈指数级增长,在不久的将来有可能获取整个生命之树(the tree of life)所有分支单元的基因组数据.因此,现代生物学与地理学、地质学、古生物学等进行交叉研究,通过整合大规模不同生物类群的遗传数据,推断其历史分布区演化,并试图将这些分布模式与影响其演化的地质过程联系起来,并试图检验或约束地质构造或环境变化历史[1 ] .近年来,科学家通过大量遗传数据的整合研究探讨了晚中新世以来安第斯山脉隆升、河流改组与亚马逊雨林生物多样性变化之间的联系[4 -5 ] ;通过南北美洲之间以及大西洋和太平洋之间生物类群演化特征来推测中美洲地峡的关闭时间[6 -7 ] ;对中国92%被子植物属的系统树和物种分布信息综合研究,表明中国约66%的被子植物在中新世期间才开始出现,中国东部是被子植物的“博物馆”,而西部尤其是青藏高原的大部分地区更像是草本植物的“摇篮”,并探讨了青藏高原隆升和亚洲季风演化对中国被子植物时空分化产生的影响[8 ] ;对中国-日本森林亚区(水杉植物区系)和中国喜马拉雅森林植物亚区(杜鹃植物区系)的综合研究表明这两个区系有着相似或相同的起源时间,都是伴随着中新世季风气候的演化而发展起来的,青藏高原隆升及其山脉走向对杜鹃植物区系的物种多样性演化产生重大影响有关[9 ] . ...
... [1 ].近年来,随着DNA测序技术的发展,已经产生了大量生物类群的遗传数据,并可以预见,这些遗传数据将呈指数级增长,在不久的将来有可能获取整个生命之树(the tree of life)所有分支单元的基因组数据.因此,现代生物学与地理学、地质学、古生物学等进行交叉研究,通过整合大规模不同生物类群的遗传数据,推断其历史分布区演化,并试图将这些分布模式与影响其演化的地质过程联系起来,并试图检验或约束地质构造或环境变化历史[1 ] .近年来,科学家通过大量遗传数据的整合研究探讨了晚中新世以来安第斯山脉隆升、河流改组与亚马逊雨林生物多样性变化之间的联系[4 -5 ] ;通过南北美洲之间以及大西洋和太平洋之间生物类群演化特征来推测中美洲地峡的关闭时间[6 -7 ] ;对中国92%被子植物属的系统树和物种分布信息综合研究,表明中国约66%的被子植物在中新世期间才开始出现,中国东部是被子植物的“博物馆”,而西部尤其是青藏高原的大部分地区更像是草本植物的“摇篮”,并探讨了青藏高原隆升和亚洲季风演化对中国被子植物时空分化产生的影响[8 ] ;对中国-日本森林亚区(水杉植物区系)和中国喜马拉雅森林植物亚区(杜鹃植物区系)的综合研究表明这两个区系有着相似或相同的起源时间,都是伴随着中新世季风气候的演化而发展起来的,青藏高原隆升及其山脉走向对杜鹃植物区系的物种多样性演化产生重大影响有关[9 ] . ...
... [1 ].近年来,科学家通过大量遗传数据的整合研究探讨了晚中新世以来安第斯山脉隆升、河流改组与亚马逊雨林生物多样性变化之间的联系[4 -5 ] ;通过南北美洲之间以及大西洋和太平洋之间生物类群演化特征来推测中美洲地峡的关闭时间[6 -7 ] ;对中国92%被子植物属的系统树和物种分布信息综合研究,表明中国约66%的被子植物在中新世期间才开始出现,中国东部是被子植物的“博物馆”,而西部尤其是青藏高原的大部分地区更像是草本植物的“摇篮”,并探讨了青藏高原隆升和亚洲季风演化对中国被子植物时空分化产生的影响[8 ] ;对中国-日本森林亚区(水杉植物区系)和中国喜马拉雅森林植物亚区(杜鹃植物区系)的综合研究表明这两个区系有着相似或相同的起源时间,都是伴随着中新世季风气候的演化而发展起来的,青藏高原隆升及其山脉走向对杜鹃植物区系的物种多样性演化产生重大影响有关[9 ] . ...
... 青藏高原及周边地区物种丰富,是全球生物多样性热点地区之一(图1 )[10 -11 ] .超过1 500属和12 000种植物生活在青藏高原地区,其中3 673种是该区域的特有种子植物[12 -14 ] .该地区植物区系独特,是全球海拔最高、高寒生物多样性最丰富的地区,被称为植物区系的“第三极”[15 ] .此外,青藏高原地区还生活着148种特有哺乳动物[16 ] ,167种特有鸟类[17 ] 以及六足动物亚门的许多特有种类[18 -19 ] .青藏高原平均海拔超过4 500 m,环境极端恶劣,低温、严寒、干旱、低氧、高紫外线辐射等是制约高寒区生物生存的关键因素,同时也是促使该区域生物特有种分化形成的主要原因[20 -21 ] .地质时期构造活动和气候变化可以形成地理隔离障碍促使物种分化和多样性增加,也可以通过减少生物扩散障碍使得物种分布区扩大,不同区域间的生物交流增加[22 -25 ] .青藏高原高寒区物种的形成和演化过程受到青藏高原构造抬升及其相应气候变化的影响[25 -26 ] .伴随着高原海拔的增长,物种就地演化或低海拔物种向高海拔迁移并适应高寒生境,或寒冷地区物种通过长距离扩散进入正在生长的高原[26 -27 ] ,导致该区域高寒生物多样性增加.高寒区生物多样性快速增长的时间通常同步于或略晚于高山或高原形成的时间[22 ,25 ,27 ] ,因此,高寒区生物类群的多样性演化过程和分化时间对我们认知构造活动期次[28 -29 ] ,推断高原隆升历史及其气候效应[1 ,30 -31 ] 等具有启示意义,甚至能够揭示以前所没有注意到的地质事件[22 ] .近年来,青藏高寒区生物地理学研究取得长足发展,为我们提供了一个独特的视角来探讨区域生物多样性演化、地质活动和气候变化之间的关系[25 ] . ...
... 生物地理学主要研究生物的时空分布格局以及性状演化过程及其形成原因和机制.从1852年开始,生物地理学家就开始寻找和总结动植物类群的分布模式,推断其历史分布区演化,并试图将这些分布模式与影响其演化的地质过程联系起来
[35 ] .随着DNA测序技术的发展,研究者对标本进行DNA测序,基于宽松分子钟模型或使用年代和鉴定准确的化石记录来校正分子钟,使用最大似然法和贝叶斯法构建系统树,估算物种演化的分支时间和演化历史
[18 ] ,通过现生类群的分布格局来推断祖先类群的地理分布.扩散-灭绝-进化分支(dispersal-extinction-cladogenesis)模型是目前最常使用的模型,使用最大似然法和贝叶斯法推断物种的扩散和灭绝历史,用隔离分化或支序发生来解释不同区域间类群分布格局
[26 ,36 ] .同时,利用比较系统发育学方法还可以估算物种多样化速率,推断物种分布格局和多样性演化历史,探讨构造事件和气候变化对多样性形成和演化过程的影响
[26 ] .
图2 生物地理学与构造、气候演化研究范式(根据文献[<xref ref-type="bibr" rid="R1">1</xref>]修改) Conceptual illustration of how biogeography can constrain geologic or climatic scenarios(Revised from Reference [<xref ref-type="bibr" rid="R1">1</xref>]) Fig.2 ![]()
青藏高原平均海拔超过4 500 m,是全球中低纬冰川分布最为集中的区域[37 ] ,被誉为世界“第三极”[38 -39 ] ,高寒生物区分布广泛,对气候变化反映敏感[40 ] .近年来,全球变暖加剧,青藏高原及周边地区升温尤其明显[41 -42 ] ,高寒区生物生存压力加大.对青藏高寒区生物开展生物地理学研究有助于探讨青藏高寒区生物多样性起源演化,并预测未来多样性演化对全球变化的响应.此外,最近的研究表明青藏高原东南缘高寒植物多样性在早渐新世就开始积累,是现存高寒生物区中起源最早的[26 ] ,远比安第斯山脉(约3.5 Ma)[43 ] 、阿尔卑斯山脉(<1.75 Ma)[44 ] 、新西兰南阿尔卑斯山脉(<5 Ma)[45 ] 、东非高山(8~7 Ma)[46 ] 高寒植物群起源地早.青藏高寒区生物多样性演化、物种扩散、分化与灭绝受到高原阶段性隆升及其气候效应的影响,对其开展研究能够进一步理解高原隆升过程及其如何对生物多样性演化产生影响. ...
... Conceptual illustration of how biogeography can constrain geologic or climatic scenarios(Revised from Reference [
1 ])
Fig.2 ![]()
青藏高原平均海拔超过4 500 m,是全球中低纬冰川分布最为集中的区域[37 ] ,被誉为世界“第三极”[38 -39 ] ,高寒生物区分布广泛,对气候变化反映敏感[40 ] .近年来,全球变暖加剧,青藏高原及周边地区升温尤其明显[41 -42 ] ,高寒区生物生存压力加大.对青藏高寒区生物开展生物地理学研究有助于探讨青藏高寒区生物多样性起源演化,并预测未来多样性演化对全球变化的响应.此外,最近的研究表明青藏高原东南缘高寒植物多样性在早渐新世就开始积累,是现存高寒生物区中起源最早的[26 ] ,远比安第斯山脉(约3.5 Ma)[43 ] 、阿尔卑斯山脉(<1.75 Ma)[44 ] 、新西兰南阿尔卑斯山脉(<5 Ma)[45 ] 、东非高山(8~7 Ma)[46 ] 高寒植物群起源地早.青藏高寒区生物多样性演化、物种扩散、分化与灭绝受到高原阶段性隆升及其气候效应的影响,对其开展研究能够进一步理解高原隆升过程及其如何对生物多样性演化产生影响. ...
2
2006
... 系统发育是指某一个生物类群的形成和发展过程[1 -2 ] ,可以通过物种系统发生关系来推测祖先类群形成的时间和分布模式,是现代生物地理学的根基[2 -3 ] .地理分布上截然不同的物种和种群是生物自然演化和响应气候或构造环境变化的结果,其遗传信息往往留有地质历史时期环境变化或构造活动的烙印,因此,重建生物类群的演化历史是回溯过去,认识地质时期环境与构造演化的契机[1 ] .近年来,随着DNA测序技术的发展,已经产生了大量生物类群的遗传数据,并可以预见,这些遗传数据将呈指数级增长,在不久的将来有可能获取整个生命之树(the tree of life)所有分支单元的基因组数据.因此,现代生物学与地理学、地质学、古生物学等进行交叉研究,通过整合大规模不同生物类群的遗传数据,推断其历史分布区演化,并试图将这些分布模式与影响其演化的地质过程联系起来,并试图检验或约束地质构造或环境变化历史[1 ] .近年来,科学家通过大量遗传数据的整合研究探讨了晚中新世以来安第斯山脉隆升、河流改组与亚马逊雨林生物多样性变化之间的联系[4 -5 ] ;通过南北美洲之间以及大西洋和太平洋之间生物类群演化特征来推测中美洲地峡的关闭时间[6 -7 ] ;对中国92%被子植物属的系统树和物种分布信息综合研究,表明中国约66%的被子植物在中新世期间才开始出现,中国东部是被子植物的“博物馆”,而西部尤其是青藏高原的大部分地区更像是草本植物的“摇篮”,并探讨了青藏高原隆升和亚洲季风演化对中国被子植物时空分化产生的影响[8 ] ;对中国-日本森林亚区(水杉植物区系)和中国喜马拉雅森林植物亚区(杜鹃植物区系)的综合研究表明这两个区系有着相似或相同的起源时间,都是伴随着中新世季风气候的演化而发展起来的,青藏高原隆升及其山脉走向对杜鹃植物区系的物种多样性演化产生重大影响有关[9 ] . ...
... [2 -3 ].地理分布上截然不同的物种和种群是生物自然演化和响应气候或构造环境变化的结果,其遗传信息往往留有地质历史时期环境变化或构造活动的烙印,因此,重建生物类群的演化历史是回溯过去,认识地质时期环境与构造演化的契机[1 ] .近年来,随着DNA测序技术的发展,已经产生了大量生物类群的遗传数据,并可以预见,这些遗传数据将呈指数级增长,在不久的将来有可能获取整个生命之树(the tree of life)所有分支单元的基因组数据.因此,现代生物学与地理学、地质学、古生物学等进行交叉研究,通过整合大规模不同生物类群的遗传数据,推断其历史分布区演化,并试图将这些分布模式与影响其演化的地质过程联系起来,并试图检验或约束地质构造或环境变化历史[1 ] .近年来,科学家通过大量遗传数据的整合研究探讨了晚中新世以来安第斯山脉隆升、河流改组与亚马逊雨林生物多样性变化之间的联系[4 -5 ] ;通过南北美洲之间以及大西洋和太平洋之间生物类群演化特征来推测中美洲地峡的关闭时间[6 -7 ] ;对中国92%被子植物属的系统树和物种分布信息综合研究,表明中国约66%的被子植物在中新世期间才开始出现,中国东部是被子植物的“博物馆”,而西部尤其是青藏高原的大部分地区更像是草本植物的“摇篮”,并探讨了青藏高原隆升和亚洲季风演化对中国被子植物时空分化产生的影响[8 ] ;对中国-日本森林亚区(水杉植物区系)和中国喜马拉雅森林植物亚区(杜鹃植物区系)的综合研究表明这两个区系有着相似或相同的起源时间,都是伴随着中新世季风气候的演化而发展起来的,青藏高原隆升及其山脉走向对杜鹃植物区系的物种多样性演化产生重大影响有关[9 ] . ...
Phylogenetic methods in biogeography
1
2011
... 系统发育是指某一个生物类群的形成和发展过程[1 -2 ] ,可以通过物种系统发生关系来推测祖先类群形成的时间和分布模式,是现代生物地理学的根基[2 -3 ] .地理分布上截然不同的物种和种群是生物自然演化和响应气候或构造环境变化的结果,其遗传信息往往留有地质历史时期环境变化或构造活动的烙印,因此,重建生物类群的演化历史是回溯过去,认识地质时期环境与构造演化的契机[1 ] .近年来,随着DNA测序技术的发展,已经产生了大量生物类群的遗传数据,并可以预见,这些遗传数据将呈指数级增长,在不久的将来有可能获取整个生命之树(the tree of life)所有分支单元的基因组数据.因此,现代生物学与地理学、地质学、古生物学等进行交叉研究,通过整合大规模不同生物类群的遗传数据,推断其历史分布区演化,并试图将这些分布模式与影响其演化的地质过程联系起来,并试图检验或约束地质构造或环境变化历史[1 ] .近年来,科学家通过大量遗传数据的整合研究探讨了晚中新世以来安第斯山脉隆升、河流改组与亚马逊雨林生物多样性变化之间的联系[4 -5 ] ;通过南北美洲之间以及大西洋和太平洋之间生物类群演化特征来推测中美洲地峡的关闭时间[6 -7 ] ;对中国92%被子植物属的系统树和物种分布信息综合研究,表明中国约66%的被子植物在中新世期间才开始出现,中国东部是被子植物的“博物馆”,而西部尤其是青藏高原的大部分地区更像是草本植物的“摇篮”,并探讨了青藏高原隆升和亚洲季风演化对中国被子植物时空分化产生的影响[8 ] ;对中国-日本森林亚区(水杉植物区系)和中国喜马拉雅森林植物亚区(杜鹃植物区系)的综合研究表明这两个区系有着相似或相同的起源时间,都是伴随着中新世季风气候的演化而发展起来的,青藏高原隆升及其山脉走向对杜鹃植物区系的物种多样性演化产生重大影响有关[9 ] . ...
Amazonia through time: Andean uplift, climate change, landscape evolution, and biodiversity
1
2010
... 系统发育是指某一个生物类群的形成和发展过程[1 -2 ] ,可以通过物种系统发生关系来推测祖先类群形成的时间和分布模式,是现代生物地理学的根基[2 -3 ] .地理分布上截然不同的物种和种群是生物自然演化和响应气候或构造环境变化的结果,其遗传信息往往留有地质历史时期环境变化或构造活动的烙印,因此,重建生物类群的演化历史是回溯过去,认识地质时期环境与构造演化的契机[1 ] .近年来,随着DNA测序技术的发展,已经产生了大量生物类群的遗传数据,并可以预见,这些遗传数据将呈指数级增长,在不久的将来有可能获取整个生命之树(the tree of life)所有分支单元的基因组数据.因此,现代生物学与地理学、地质学、古生物学等进行交叉研究,通过整合大规模不同生物类群的遗传数据,推断其历史分布区演化,并试图将这些分布模式与影响其演化的地质过程联系起来,并试图检验或约束地质构造或环境变化历史[1 ] .近年来,科学家通过大量遗传数据的整合研究探讨了晚中新世以来安第斯山脉隆升、河流改组与亚马逊雨林生物多样性变化之间的联系[4 -5 ] ;通过南北美洲之间以及大西洋和太平洋之间生物类群演化特征来推测中美洲地峡的关闭时间[6 -7 ] ;对中国92%被子植物属的系统树和物种分布信息综合研究,表明中国约66%的被子植物在中新世期间才开始出现,中国东部是被子植物的“博物馆”,而西部尤其是青藏高原的大部分地区更像是草本植物的“摇篮”,并探讨了青藏高原隆升和亚洲季风演化对中国被子植物时空分化产生的影响[8 ] ;对中国-日本森林亚区(水杉植物区系)和中国喜马拉雅森林植物亚区(杜鹃植物区系)的综合研究表明这两个区系有着相似或相同的起源时间,都是伴随着中新世季风气候的演化而发展起来的,青藏高原隆升及其山脉走向对杜鹃植物区系的物种多样性演化产生重大影响有关[9 ] . ...
Quaternary climate changes as speciation drivers in the Amazon floodplains
1
2020
... 系统发育是指某一个生物类群的形成和发展过程[1 -2 ] ,可以通过物种系统发生关系来推测祖先类群形成的时间和分布模式,是现代生物地理学的根基[2 -3 ] .地理分布上截然不同的物种和种群是生物自然演化和响应气候或构造环境变化的结果,其遗传信息往往留有地质历史时期环境变化或构造活动的烙印,因此,重建生物类群的演化历史是回溯过去,认识地质时期环境与构造演化的契机[1 ] .近年来,随着DNA测序技术的发展,已经产生了大量生物类群的遗传数据,并可以预见,这些遗传数据将呈指数级增长,在不久的将来有可能获取整个生命之树(the tree of life)所有分支单元的基因组数据.因此,现代生物学与地理学、地质学、古生物学等进行交叉研究,通过整合大规模不同生物类群的遗传数据,推断其历史分布区演化,并试图将这些分布模式与影响其演化的地质过程联系起来,并试图检验或约束地质构造或环境变化历史[1 ] .近年来,科学家通过大量遗传数据的整合研究探讨了晚中新世以来安第斯山脉隆升、河流改组与亚马逊雨林生物多样性变化之间的联系[4 -5 ] ;通过南北美洲之间以及大西洋和太平洋之间生物类群演化特征来推测中美洲地峡的关闭时间[6 -7 ] ;对中国92%被子植物属的系统树和物种分布信息综合研究,表明中国约66%的被子植物在中新世期间才开始出现,中国东部是被子植物的“博物馆”,而西部尤其是青藏高原的大部分地区更像是草本植物的“摇篮”,并探讨了青藏高原隆升和亚洲季风演化对中国被子植物时空分化产生的影响[8 ] ;对中国-日本森林亚区(水杉植物区系)和中国喜马拉雅森林植物亚区(杜鹃植物区系)的综合研究表明这两个区系有着相似或相同的起源时间,都是伴随着中新世季风气候的演化而发展起来的,青藏高原隆升及其山脉走向对杜鹃植物区系的物种多样性演化产生重大影响有关[9 ] . ...
Biological evidence supports an early and complex emergence of the Isthmus of Panama
1
2015
... 系统发育是指某一个生物类群的形成和发展过程[1 -2 ] ,可以通过物种系统发生关系来推测祖先类群形成的时间和分布模式,是现代生物地理学的根基[2 -3 ] .地理分布上截然不同的物种和种群是生物自然演化和响应气候或构造环境变化的结果,其遗传信息往往留有地质历史时期环境变化或构造活动的烙印,因此,重建生物类群的演化历史是回溯过去,认识地质时期环境与构造演化的契机[1 ] .近年来,随着DNA测序技术的发展,已经产生了大量生物类群的遗传数据,并可以预见,这些遗传数据将呈指数级增长,在不久的将来有可能获取整个生命之树(the tree of life)所有分支单元的基因组数据.因此,现代生物学与地理学、地质学、古生物学等进行交叉研究,通过整合大规模不同生物类群的遗传数据,推断其历史分布区演化,并试图将这些分布模式与影响其演化的地质过程联系起来,并试图检验或约束地质构造或环境变化历史[1 ] .近年来,科学家通过大量遗传数据的整合研究探讨了晚中新世以来安第斯山脉隆升、河流改组与亚马逊雨林生物多样性变化之间的联系[4 -5 ] ;通过南北美洲之间以及大西洋和太平洋之间生物类群演化特征来推测中美洲地峡的关闭时间[6 -7 ] ;对中国92%被子植物属的系统树和物种分布信息综合研究,表明中国约66%的被子植物在中新世期间才开始出现,中国东部是被子植物的“博物馆”,而西部尤其是青藏高原的大部分地区更像是草本植物的“摇篮”,并探讨了青藏高原隆升和亚洲季风演化对中国被子植物时空分化产生的影响[8 ] ;对中国-日本森林亚区(水杉植物区系)和中国喜马拉雅森林植物亚区(杜鹃植物区系)的综合研究表明这两个区系有着相似或相同的起源时间,都是伴随着中新世季风气候的演化而发展起来的,青藏高原隆升及其山脉走向对杜鹃植物区系的物种多样性演化产生重大影响有关[9 ] . ...
Formation of the Isthmus of Panama
1
2016
... 系统发育是指某一个生物类群的形成和发展过程[1 -2 ] ,可以通过物种系统发生关系来推测祖先类群形成的时间和分布模式,是现代生物地理学的根基[2 -3 ] .地理分布上截然不同的物种和种群是生物自然演化和响应气候或构造环境变化的结果,其遗传信息往往留有地质历史时期环境变化或构造活动的烙印,因此,重建生物类群的演化历史是回溯过去,认识地质时期环境与构造演化的契机[1 ] .近年来,随着DNA测序技术的发展,已经产生了大量生物类群的遗传数据,并可以预见,这些遗传数据将呈指数级增长,在不久的将来有可能获取整个生命之树(the tree of life)所有分支单元的基因组数据.因此,现代生物学与地理学、地质学、古生物学等进行交叉研究,通过整合大规模不同生物类群的遗传数据,推断其历史分布区演化,并试图将这些分布模式与影响其演化的地质过程联系起来,并试图检验或约束地质构造或环境变化历史[1 ] .近年来,科学家通过大量遗传数据的整合研究探讨了晚中新世以来安第斯山脉隆升、河流改组与亚马逊雨林生物多样性变化之间的联系[4 -5 ] ;通过南北美洲之间以及大西洋和太平洋之间生物类群演化特征来推测中美洲地峡的关闭时间[6 -7 ] ;对中国92%被子植物属的系统树和物种分布信息综合研究,表明中国约66%的被子植物在中新世期间才开始出现,中国东部是被子植物的“博物馆”,而西部尤其是青藏高原的大部分地区更像是草本植物的“摇篮”,并探讨了青藏高原隆升和亚洲季风演化对中国被子植物时空分化产生的影响[8 ] ;对中国-日本森林亚区(水杉植物区系)和中国喜马拉雅森林植物亚区(杜鹃植物区系)的综合研究表明这两个区系有着相似或相同的起源时间,都是伴随着中新世季风气候的演化而发展起来的,青藏高原隆升及其山脉走向对杜鹃植物区系的物种多样性演化产生重大影响有关[9 ] . ...
Evolutionary history of the angiosperm flora of China
1
2018
... 系统发育是指某一个生物类群的形成和发展过程[1 -2 ] ,可以通过物种系统发生关系来推测祖先类群形成的时间和分布模式,是现代生物地理学的根基[2 -3 ] .地理分布上截然不同的物种和种群是生物自然演化和响应气候或构造环境变化的结果,其遗传信息往往留有地质历史时期环境变化或构造活动的烙印,因此,重建生物类群的演化历史是回溯过去,认识地质时期环境与构造演化的契机[1 ] .近年来,随着DNA测序技术的发展,已经产生了大量生物类群的遗传数据,并可以预见,这些遗传数据将呈指数级增长,在不久的将来有可能获取整个生命之树(the tree of life)所有分支单元的基因组数据.因此,现代生物学与地理学、地质学、古生物学等进行交叉研究,通过整合大规模不同生物类群的遗传数据,推断其历史分布区演化,并试图将这些分布模式与影响其演化的地质过程联系起来,并试图检验或约束地质构造或环境变化历史[1 ] .近年来,科学家通过大量遗传数据的整合研究探讨了晚中新世以来安第斯山脉隆升、河流改组与亚马逊雨林生物多样性变化之间的联系[4 -5 ] ;通过南北美洲之间以及大西洋和太平洋之间生物类群演化特征来推测中美洲地峡的关闭时间[6 -7 ] ;对中国92%被子植物属的系统树和物种分布信息综合研究,表明中国约66%的被子植物在中新世期间才开始出现,中国东部是被子植物的“博物馆”,而西部尤其是青藏高原的大部分地区更像是草本植物的“摇篮”,并探讨了青藏高原隆升和亚洲季风演化对中国被子植物时空分化产生的影响[8 ] ;对中国-日本森林亚区(水杉植物区系)和中国喜马拉雅森林植物亚区(杜鹃植物区系)的综合研究表明这两个区系有着相似或相同的起源时间,都是伴随着中新世季风气候的演化而发展起来的,青藏高原隆升及其山脉走向对杜鹃植物区系的物种多样性演化产生重大影响有关[9 ] . ...
Is the East Asian flora ancient or not?
1
2018
... 系统发育是指某一个生物类群的形成和发展过程[1 -2 ] ,可以通过物种系统发生关系来推测祖先类群形成的时间和分布模式,是现代生物地理学的根基[2 -3 ] .地理分布上截然不同的物种和种群是生物自然演化和响应气候或构造环境变化的结果,其遗传信息往往留有地质历史时期环境变化或构造活动的烙印,因此,重建生物类群的演化历史是回溯过去,认识地质时期环境与构造演化的契机[1 ] .近年来,随着DNA测序技术的发展,已经产生了大量生物类群的遗传数据,并可以预见,这些遗传数据将呈指数级增长,在不久的将来有可能获取整个生命之树(the tree of life)所有分支单元的基因组数据.因此,现代生物学与地理学、地质学、古生物学等进行交叉研究,通过整合大规模不同生物类群的遗传数据,推断其历史分布区演化,并试图将这些分布模式与影响其演化的地质过程联系起来,并试图检验或约束地质构造或环境变化历史[1 ] .近年来,科学家通过大量遗传数据的整合研究探讨了晚中新世以来安第斯山脉隆升、河流改组与亚马逊雨林生物多样性变化之间的联系[4 -5 ] ;通过南北美洲之间以及大西洋和太平洋之间生物类群演化特征来推测中美洲地峡的关闭时间[6 -7 ] ;对中国92%被子植物属的系统树和物种分布信息综合研究,表明中国约66%的被子植物在中新世期间才开始出现,中国东部是被子植物的“博物馆”,而西部尤其是青藏高原的大部分地区更像是草本植物的“摇篮”,并探讨了青藏高原隆升和亚洲季风演化对中国被子植物时空分化产生的影响[8 ] ;对中国-日本森林亚区(水杉植物区系)和中国喜马拉雅森林植物亚区(杜鹃植物区系)的综合研究表明这两个区系有着相似或相同的起源时间,都是伴随着中新世季风气候的演化而发展起来的,青藏高原隆升及其山脉走向对杜鹃植物区系的物种多样性演化产生重大影响有关[9 ] . ...
The role of the uplift of the Qinghai‐Tibetan Plateau for the evolution of Tibetan biotas
3
2015
... 青藏高原及周边地区物种丰富,是全球生物多样性热点地区之一(图1 )[10 -11 ] .超过1 500属和12 000种植物生活在青藏高原地区,其中3 673种是该区域的特有种子植物[12 -14 ] .该地区植物区系独特,是全球海拔最高、高寒生物多样性最丰富的地区,被称为植物区系的“第三极”[15 ] .此外,青藏高原地区还生活着148种特有哺乳动物[16 ] ,167种特有鸟类[17 ] 以及六足动物亚门的许多特有种类[18 -19 ] .青藏高原平均海拔超过4 500 m,环境极端恶劣,低温、严寒、干旱、低氧、高紫外线辐射等是制约高寒区生物生存的关键因素,同时也是促使该区域生物特有种分化形成的主要原因[20 -21 ] .地质时期构造活动和气候变化可以形成地理隔离障碍促使物种分化和多样性增加,也可以通过减少生物扩散障碍使得物种分布区扩大,不同区域间的生物交流增加[22 -25 ] .青藏高原高寒区物种的形成和演化过程受到青藏高原构造抬升及其相应气候变化的影响[25 -26 ] .伴随着高原海拔的增长,物种就地演化或低海拔物种向高海拔迁移并适应高寒生境,或寒冷地区物种通过长距离扩散进入正在生长的高原[26 -27 ] ,导致该区域高寒生物多样性增加.高寒区生物多样性快速增长的时间通常同步于或略晚于高山或高原形成的时间[22 ,25 ,27 ] ,因此,高寒区生物类群的多样性演化过程和分化时间对我们认知构造活动期次[28 -29 ] ,推断高原隆升历史及其气候效应[1 ,30 -31 ] 等具有启示意义,甚至能够揭示以前所没有注意到的地质事件[22 ] .近年来,青藏高寒区生物地理学研究取得长足发展,为我们提供了一个独特的视角来探讨区域生物多样性演化、地质活动和气候变化之间的关系[25 ] . ...
... 青藏高原及周边生物多样性热点地区(根据文献[
10 ]修改)
①中亚山地;②喜马拉雅山脉;③中缅地区;④横断山区 ...
... ①中亚山地;②喜马拉雅山脉;③中缅地区;④横断山区
Maps of the four hotspots of biodiversity of the Tibetan Plateau (Revised from Reference [<xref ref-type="bibr" rid="R10">10</xref>]) ①Mountains of Central Asia;②Himalaya;③China-Myanmar Region;④Hengduan Mountains ...
Biodiversity hotspots for conservation priorities
1
2000
... 青藏高原及周边地区物种丰富,是全球生物多样性热点地区之一(图1 )[10 -11 ] .超过1 500属和12 000种植物生活在青藏高原地区,其中3 673种是该区域的特有种子植物[12 -14 ] .该地区植物区系独特,是全球海拔最高、高寒生物多样性最丰富的地区,被称为植物区系的“第三极”[15 ] .此外,青藏高原地区还生活着148种特有哺乳动物[16 ] ,167种特有鸟类[17 ] 以及六足动物亚门的许多特有种类[18 -19 ] .青藏高原平均海拔超过4 500 m,环境极端恶劣,低温、严寒、干旱、低氧、高紫外线辐射等是制约高寒区生物生存的关键因素,同时也是促使该区域生物特有种分化形成的主要原因[20 -21 ] .地质时期构造活动和气候变化可以形成地理隔离障碍促使物种分化和多样性增加,也可以通过减少生物扩散障碍使得物种分布区扩大,不同区域间的生物交流增加[22 -25 ] .青藏高原高寒区物种的形成和演化过程受到青藏高原构造抬升及其相应气候变化的影响[25 -26 ] .伴随着高原海拔的增长,物种就地演化或低海拔物种向高海拔迁移并适应高寒生境,或寒冷地区物种通过长距离扩散进入正在生长的高原[26 -27 ] ,导致该区域高寒生物多样性增加.高寒区生物多样性快速增长的时间通常同步于或略晚于高山或高原形成的时间[22 ,25 ,27 ] ,因此,高寒区生物类群的多样性演化过程和分化时间对我们认知构造活动期次[28 -29 ] ,推断高原隆升历史及其气候效应[1 ,30 -31 ] 等具有启示意义,甚至能够揭示以前所没有注意到的地质事件[22 ] .近年来,青藏高寒区生物地理学研究取得长足发展,为我们提供了一个独特的视角来探讨区域生物多样性演化、地质活动和气候变化之间的关系[25 ] . ...
1
1987
... 青藏高原及周边地区物种丰富,是全球生物多样性热点地区之一(图1 )[10 -11 ] .超过1 500属和12 000种植物生活在青藏高原地区,其中3 673种是该区域的特有种子植物[12 -14 ] .该地区植物区系独特,是全球海拔最高、高寒生物多样性最丰富的地区,被称为植物区系的“第三极”[15 ] .此外,青藏高原地区还生活着148种特有哺乳动物[16 ] ,167种特有鸟类[17 ] 以及六足动物亚门的许多特有种类[18 -19 ] .青藏高原平均海拔超过4 500 m,环境极端恶劣,低温、严寒、干旱、低氧、高紫外线辐射等是制约高寒区生物生存的关键因素,同时也是促使该区域生物特有种分化形成的主要原因[20 -21 ] .地质时期构造活动和气候变化可以形成地理隔离障碍促使物种分化和多样性增加,也可以通过减少生物扩散障碍使得物种分布区扩大,不同区域间的生物交流增加[22 -25 ] .青藏高原高寒区物种的形成和演化过程受到青藏高原构造抬升及其相应气候变化的影响[25 -26 ] .伴随着高原海拔的增长,物种就地演化或低海拔物种向高海拔迁移并适应高寒生境,或寒冷地区物种通过长距离扩散进入正在生长的高原[26 -27 ] ,导致该区域高寒生物多样性增加.高寒区生物多样性快速增长的时间通常同步于或略晚于高山或高原形成的时间[22 ,25 ,27 ] ,因此,高寒区生物类群的多样性演化过程和分化时间对我们认知构造活动期次[28 -29 ] ,推断高原隆升历史及其气候效应[1 ,30 -31 ] 等具有启示意义,甚至能够揭示以前所没有注意到的地质事件[22 ] .近年来,青藏高寒区生物地理学研究取得长足发展,为我们提供了一个独特的视角来探讨区域生物多样性演化、地质活动和气候变化之间的关系[25 ] . ...
1
1987
... 青藏高原及周边地区物种丰富,是全球生物多样性热点地区之一(图1 )[10 -11 ] .超过1 500属和12 000种植物生活在青藏高原地区,其中3 673种是该区域的特有种子植物[12 -14 ] .该地区植物区系独特,是全球海拔最高、高寒生物多样性最丰富的地区,被称为植物区系的“第三极”[15 ] .此外,青藏高原地区还生活着148种特有哺乳动物[16 ] ,167种特有鸟类[17 ] 以及六足动物亚门的许多特有种类[18 -19 ] .青藏高原平均海拔超过4 500 m,环境极端恶劣,低温、严寒、干旱、低氧、高紫外线辐射等是制约高寒区生物生存的关键因素,同时也是促使该区域生物特有种分化形成的主要原因[20 -21 ] .地质时期构造活动和气候变化可以形成地理隔离障碍促使物种分化和多样性增加,也可以通过减少生物扩散障碍使得物种分布区扩大,不同区域间的生物交流增加[22 -25 ] .青藏高原高寒区物种的形成和演化过程受到青藏高原构造抬升及其相应气候变化的影响[25 -26 ] .伴随着高原海拔的增长,物种就地演化或低海拔物种向高海拔迁移并适应高寒生境,或寒冷地区物种通过长距离扩散进入正在生长的高原[26 -27 ] ,导致该区域高寒生物多样性增加.高寒区生物多样性快速增长的时间通常同步于或略晚于高山或高原形成的时间[22 ,25 ,27 ] ,因此,高寒区生物类群的多样性演化过程和分化时间对我们认知构造活动期次[28 -29 ] ,推断高原隆升历史及其气候效应[1 ,30 -31 ] 等具有启示意义,甚至能够揭示以前所没有注意到的地质事件[22 ] .近年来,青藏高寒区生物地理学研究取得长足发展,为我们提供了一个独特的视角来探讨区域生物多样性演化、地质活动和气候变化之间的关系[25 ] . ...
Testing multiple hypotheses for the high endemic plant diversity of the Tibetan Plateau
1
2019
... 青藏高原及周边地区物种丰富,是全球生物多样性热点地区之一(图1 )[10 -11 ] .超过1 500属和12 000种植物生活在青藏高原地区,其中3 673种是该区域的特有种子植物[12 -14 ] .该地区植物区系独特,是全球海拔最高、高寒生物多样性最丰富的地区,被称为植物区系的“第三极”[15 ] .此外,青藏高原地区还生活着148种特有哺乳动物[16 ] ,167种特有鸟类[17 ] 以及六足动物亚门的许多特有种类[18 -19 ] .青藏高原平均海拔超过4 500 m,环境极端恶劣,低温、严寒、干旱、低氧、高紫外线辐射等是制约高寒区生物生存的关键因素,同时也是促使该区域生物特有种分化形成的主要原因[20 -21 ] .地质时期构造活动和气候变化可以形成地理隔离障碍促使物种分化和多样性增加,也可以通过减少生物扩散障碍使得物种分布区扩大,不同区域间的生物交流增加[22 -25 ] .青藏高原高寒区物种的形成和演化过程受到青藏高原构造抬升及其相应气候变化的影响[25 -26 ] .伴随着高原海拔的增长,物种就地演化或低海拔物种向高海拔迁移并适应高寒生境,或寒冷地区物种通过长距离扩散进入正在生长的高原[26 -27 ] ,导致该区域高寒生物多样性增加.高寒区生物多样性快速增长的时间通常同步于或略晚于高山或高原形成的时间[22 ,25 ,27 ] ,因此,高寒区生物类群的多样性演化过程和分化时间对我们认知构造活动期次[28 -29 ] ,推断高原隆升历史及其气候效应[1 ,30 -31 ] 等具有启示意义,甚至能够揭示以前所没有注意到的地质事件[22 ] .近年来,青藏高寒区生物地理学研究取得长足发展,为我们提供了一个独特的视角来探讨区域生物多样性演化、地质活动和气候变化之间的关系[25 ] . ...
青藏高原高寒地区种子植物区系的研究
1
1995
... 青藏高原及周边地区物种丰富,是全球生物多样性热点地区之一(图1 )[10 -11 ] .超过1 500属和12 000种植物生活在青藏高原地区,其中3 673种是该区域的特有种子植物[12 -14 ] .该地区植物区系独特,是全球海拔最高、高寒生物多样性最丰富的地区,被称为植物区系的“第三极”[15 ] .此外,青藏高原地区还生活着148种特有哺乳动物[16 ] ,167种特有鸟类[17 ] 以及六足动物亚门的许多特有种类[18 -19 ] .青藏高原平均海拔超过4 500 m,环境极端恶劣,低温、严寒、干旱、低氧、高紫外线辐射等是制约高寒区生物生存的关键因素,同时也是促使该区域生物特有种分化形成的主要原因[20 -21 ] .地质时期构造活动和气候变化可以形成地理隔离障碍促使物种分化和多样性增加,也可以通过减少生物扩散障碍使得物种分布区扩大,不同区域间的生物交流增加[22 -25 ] .青藏高原高寒区物种的形成和演化过程受到青藏高原构造抬升及其相应气候变化的影响[25 -26 ] .伴随着高原海拔的增长,物种就地演化或低海拔物种向高海拔迁移并适应高寒生境,或寒冷地区物种通过长距离扩散进入正在生长的高原[26 -27 ] ,导致该区域高寒生物多样性增加.高寒区生物多样性快速增长的时间通常同步于或略晚于高山或高原形成的时间[22 ,25 ,27 ] ,因此,高寒区生物类群的多样性演化过程和分化时间对我们认知构造活动期次[28 -29 ] ,推断高原隆升历史及其气候效应[1 ,30 -31 ] 等具有启示意义,甚至能够揭示以前所没有注意到的地质事件[22 ] .近年来,青藏高寒区生物地理学研究取得长足发展,为我们提供了一个独特的视角来探讨区域生物多样性演化、地质活动和气候变化之间的关系[25 ] . ...
青藏高原高寒地区种子植物区系的研究
1
1995
... 青藏高原及周边地区物种丰富,是全球生物多样性热点地区之一(图1 )[10 -11 ] .超过1 500属和12 000种植物生活在青藏高原地区,其中3 673种是该区域的特有种子植物[12 -14 ] .该地区植物区系独特,是全球海拔最高、高寒生物多样性最丰富的地区,被称为植物区系的“第三极”[15 ] .此外,青藏高原地区还生活着148种特有哺乳动物[16 ] ,167种特有鸟类[17 ] 以及六足动物亚门的许多特有种类[18 -19 ] .青藏高原平均海拔超过4 500 m,环境极端恶劣,低温、严寒、干旱、低氧、高紫外线辐射等是制约高寒区生物生存的关键因素,同时也是促使该区域生物特有种分化形成的主要原因[20 -21 ] .地质时期构造活动和气候变化可以形成地理隔离障碍促使物种分化和多样性增加,也可以通过减少生物扩散障碍使得物种分布区扩大,不同区域间的生物交流增加[22 -25 ] .青藏高原高寒区物种的形成和演化过程受到青藏高原构造抬升及其相应气候变化的影响[25 -26 ] .伴随着高原海拔的增长,物种就地演化或低海拔物种向高海拔迁移并适应高寒生境,或寒冷地区物种通过长距离扩散进入正在生长的高原[26 -27 ] ,导致该区域高寒生物多样性增加.高寒区生物多样性快速增长的时间通常同步于或略晚于高山或高原形成的时间[22 ,25 ,27 ] ,因此,高寒区生物类群的多样性演化过程和分化时间对我们认知构造活动期次[28 -29 ] ,推断高原隆升历史及其气候效应[1 ,30 -31 ] 等具有启示意义,甚至能够揭示以前所没有注意到的地质事件[22 ] .近年来,青藏高寒区生物地理学研究取得长足发展,为我们提供了一个独特的视角来探讨区域生物多样性演化、地质活动和气候变化之间的关系[25 ] . ...
Quantitative analysis of land mammal zoogeographical regions in China and adjacent regions
1
2004
... 青藏高原及周边地区物种丰富,是全球生物多样性热点地区之一(图1 )[10 -11 ] .超过1 500属和12 000种植物生活在青藏高原地区,其中3 673种是该区域的特有种子植物[12 -14 ] .该地区植物区系独特,是全球海拔最高、高寒生物多样性最丰富的地区,被称为植物区系的“第三极”[15 ] .此外,青藏高原地区还生活着148种特有哺乳动物[16 ] ,167种特有鸟类[17 ] 以及六足动物亚门的许多特有种类[18 -19 ] .青藏高原平均海拔超过4 500 m,环境极端恶劣,低温、严寒、干旱、低氧、高紫外线辐射等是制约高寒区生物生存的关键因素,同时也是促使该区域生物特有种分化形成的主要原因[20 -21 ] .地质时期构造活动和气候变化可以形成地理隔离障碍促使物种分化和多样性增加,也可以通过减少生物扩散障碍使得物种分布区扩大,不同区域间的生物交流增加[22 -25 ] .青藏高原高寒区物种的形成和演化过程受到青藏高原构造抬升及其相应气候变化的影响[25 -26 ] .伴随着高原海拔的增长,物种就地演化或低海拔物种向高海拔迁移并适应高寒生境,或寒冷地区物种通过长距离扩散进入正在生长的高原[26 -27 ] ,导致该区域高寒生物多样性增加.高寒区生物多样性快速增长的时间通常同步于或略晚于高山或高原形成的时间[22 ,25 ,27 ] ,因此,高寒区生物类群的多样性演化过程和分化时间对我们认知构造活动期次[28 -29 ] ,推断高原隆升历史及其气候效应[1 ,30 -31 ] 等具有启示意义,甚至能够揭示以前所没有注意到的地质事件[22 ] .近年来,青藏高寒区生物地理学研究取得长足发展,为我们提供了一个独特的视角来探讨区域生物多样性演化、地质活动和气候变化之间的关系[25 ] . ...
Mobile hotspots and refugia of avian diversity in the mountains of south-west China under past and contemporary global climate change
1
2017
... 青藏高原及周边地区物种丰富,是全球生物多样性热点地区之一(图1 )[10 -11 ] .超过1 500属和12 000种植物生活在青藏高原地区,其中3 673种是该区域的特有种子植物[12 -14 ] .该地区植物区系独特,是全球海拔最高、高寒生物多样性最丰富的地区,被称为植物区系的“第三极”[15 ] .此外,青藏高原地区还生活着148种特有哺乳动物[16 ] ,167种特有鸟类[17 ] 以及六足动物亚门的许多特有种类[18 -19 ] .青藏高原平均海拔超过4 500 m,环境极端恶劣,低温、严寒、干旱、低氧、高紫外线辐射等是制约高寒区生物生存的关键因素,同时也是促使该区域生物特有种分化形成的主要原因[20 -21 ] .地质时期构造活动和气候变化可以形成地理隔离障碍促使物种分化和多样性增加,也可以通过减少生物扩散障碍使得物种分布区扩大,不同区域间的生物交流增加[22 -25 ] .青藏高原高寒区物种的形成和演化过程受到青藏高原构造抬升及其相应气候变化的影响[25 -26 ] .伴随着高原海拔的增长,物种就地演化或低海拔物种向高海拔迁移并适应高寒生境,或寒冷地区物种通过长距离扩散进入正在生长的高原[26 -27 ] ,导致该区域高寒生物多样性增加.高寒区生物多样性快速增长的时间通常同步于或略晚于高山或高原形成的时间[22 ,25 ,27 ] ,因此,高寒区生物类群的多样性演化过程和分化时间对我们认知构造活动期次[28 -29 ] ,推断高原隆升历史及其气候效应[1 ,30 -31 ] 等具有启示意义,甚至能够揭示以前所没有注意到的地质事件[22 ] .近年来,青藏高寒区生物地理学研究取得长足发展,为我们提供了一个独特的视角来探讨区域生物多样性演化、地质活动和气候变化之间的关系[25 ] . ...
青藏高原琵甲族部分属的系统发育与历史生物地理学
4
2020
... 青藏高原及周边地区物种丰富,是全球生物多样性热点地区之一(图1 )[10 -11 ] .超过1 500属和12 000种植物生活在青藏高原地区,其中3 673种是该区域的特有种子植物[12 -14 ] .该地区植物区系独特,是全球海拔最高、高寒生物多样性最丰富的地区,被称为植物区系的“第三极”[15 ] .此外,青藏高原地区还生活着148种特有哺乳动物[16 ] ,167种特有鸟类[17 ] 以及六足动物亚门的许多特有种类[18 -19 ] .青藏高原平均海拔超过4 500 m,环境极端恶劣,低温、严寒、干旱、低氧、高紫外线辐射等是制约高寒区生物生存的关键因素,同时也是促使该区域生物特有种分化形成的主要原因[20 -21 ] .地质时期构造活动和气候变化可以形成地理隔离障碍促使物种分化和多样性增加,也可以通过减少生物扩散障碍使得物种分布区扩大,不同区域间的生物交流增加[22 -25 ] .青藏高原高寒区物种的形成和演化过程受到青藏高原构造抬升及其相应气候变化的影响[25 -26 ] .伴随着高原海拔的增长,物种就地演化或低海拔物种向高海拔迁移并适应高寒生境,或寒冷地区物种通过长距离扩散进入正在生长的高原[26 -27 ] ,导致该区域高寒生物多样性增加.高寒区生物多样性快速增长的时间通常同步于或略晚于高山或高原形成的时间[22 ,25 ,27 ] ,因此,高寒区生物类群的多样性演化过程和分化时间对我们认知构造活动期次[28 -29 ] ,推断高原隆升历史及其气候效应[1 ,30 -31 ] 等具有启示意义,甚至能够揭示以前所没有注意到的地质事件[22 ] .近年来,青藏高寒区生物地理学研究取得长足发展,为我们提供了一个独特的视角来探讨区域生物多样性演化、地质活动和气候变化之间的关系[25 ] . ...
... 生物地理学主要研究生物的时空分布格局以及性状演化过程及其形成原因和机制.从1852年开始,生物地理学家就开始寻找和总结动植物类群的分布模式,推断其历史分布区演化,并试图将这些分布模式与影响其演化的地质过程联系起来[35 ] .随着DNA测序技术的发展,研究者对标本进行DNA测序,基于宽松分子钟模型或使用年代和鉴定准确的化石记录来校正分子钟,使用最大似然法和贝叶斯法构建系统树,估算物种演化的分支时间和演化历史[18 ] ,通过现生类群的分布格局来推断祖先类群的地理分布.扩散-灭绝-进化分支(dispersal-extinction-cladogenesis)模型是目前最常使用的模型,使用最大似然法和贝叶斯法推断物种的扩散和灭绝历史,用隔离分化或支序发生来解释不同区域间类群分布格局[26 ,36 ] .同时,利用比较系统发育学方法还可以估算物种多样化速率,推断物种分布格局和多样性演化历史,探讨构造事件和气候变化对多样性形成和演化过程的影响[26 ] . ...
... 小琵甲属(Gnaptorina )是琵甲族的高原特有属,已知的34种全部分布在青藏高原及周边地区海拔3 000~4 500 m的区域,该属昆虫后翅愈合,不能飞翔,是对高海拔极端环境的一种适应性状,小琵甲属生物地理学研究表明其祖先类群在始新世起源于青藏高原东南缘,高原隆升和河流切割造成的地理隔离阻断了小琵甲属物种或种群之间的基因交流,加速特有种的快速辐射演化[18 -19 ] ;隙蛛(Coelotinae )是北半球温带和亚热带地区常见蜘蛛种类,青藏高原东南缘晚始新世形成的高海拔地貌阻碍了南方隙蛛向北扩散,但是随之形成的复杂地貌创造的局部地理隔离也促使南方隙蛛在此阶段快速辐射演化,渐新世以来高原中部和北部的隆起导致北方隙蛛在早期多样化过程中经历了灭绝事件[59 ] . ...
... Biological evolution in the alpine region of Tibetan Plateau
Table 1 种属 分布区域 适应高海拔类群分化时间或多样性增加时间 文献来源 Sinopodophyllum hexandrum 青藏高原东部 在约(6.52±1.89) Ma期间出现在青藏高原东部,但是直到3~2 Ma以来才演化出适应高山环境的形态 [47 ] Nannoglottis 青藏高原 在23~32 Ma期间出现在青藏高原,随后多样性增高,其高山灌丛和针叶林生活类型分支时间约为3.4 Ma [48 ] Reaumuria soongarica 西北干旱区、青藏高原北部 高原内部分布类群演化时间2.0~6.3 Ma,高原东北缘与高原西北类群分化时间1.1~3.5 Ma [49 ] Orchidinae 青藏高原及喜马拉雅山脉 青藏高原特有种大多是在过去6 Ma里进化而来 [50 ] Oxyria sinensis 青藏高原东南部 高山类群出现时间7.54~24.36 Ma,种内多样化增加时间1.74~0.86 Ma [51 ] Juniperus microsperma 西藏波密 早中新世21.6~14 Ma期间与其他种群分化 [52 ] Rhododendron 北半球 横断山区概属多样性在晚中新世(约8 Ma)快速增加 [53 ] Roscoea, Cautleya 喜马拉雅-横断山区 在早渐新世(~32 Ma)两属出现,Roscoea在横断山区与喜马拉雅山脉的两个分支大概在渐新世-中新世之交分化 [54 ] Nanorana parkeri 青藏高原南部 高原东部和西部类群分化时间1.4~3.7 Ma [55 ] Paini 南亚、东南亚、青藏高原 在渐新世早期棘蛙族高原-喜马拉雅类群与中国第二阶梯上生活的类群分化,随后在在26.1~12.5 Ma高原生活类群与喜马拉雅山脉生活类群分化,高原内部生活类群在晚中新世到上新世期间分化速率加快 [22 ] Bufo tibetanus 青藏高原、四川、甘肃、陕西等 高原种类形成于上新世期间 [56 ] Phrynocephalus 亚洲干旱区 在7.2~13.0 Ma期间演化出胎生类群,3.2~7.3 Ma期间种间多样性增加 [57 ] Phrynocephalus erythrurus 羌塘盆地 在3.67 Ma左右北部类群与南部类群发生分化,约2.76 Ma北部类群内部进一步分化 [58 ] Gnaptorina 青藏高原及周边高海拔地区 始新世起源于青藏高原东南缘 [18 -19 ] Coelotinae 北半球温带、亚热带 晚始新世高原东南缘隆升阻碍南方隙蛛向北扩散,渐新世高原中部和北部隆起导致北方隙蛛早期多样化进程中的灭绝事件 [59 ] Montifringilla, Onychostruthus, Pyrgilauda 青藏高原 雪雀的共同祖先在约14 Ma就应经生活在青藏高原,并进化出适应高海拔极端环境的性状 [21 ] Parus major 亚洲、欧洲 蒙古分布类群与其他种类分化时间为2.8~0.7 Ma,喜马拉雅山脉东部与中国中东部类群分化时间为1.9~0.4 Ma [60 ] Pseudopodoces humilis 青藏高原、柴达木盆地周边高山 在9.9~7.7 Ma期间地山雀就与与其他山雀分化开来,并出现适应高海拔性状 [61 ] Phylloscopus, Seicercus 亚洲 喜马拉雅山脉分布类群在7 Ma加速进化 [62 -63 ] Perdix 西藏、甘肃、青海、四川、新疆 高原类群出现于3.63 (4.52~2.75) Ma [64 ] Pantholops hodgsonii 西藏、新疆、青海 上新世-更新世就已经出现在青藏高原 [65 ] Procapra picticaudata 西藏、四川、甘肃、青海、新疆 种内分化时间约2.2~4.4 Ma [66 ] Rhinopithecus 四川、甘肃、陕西、贵州、云南、缅甸、越南 北方类群川金丝猴、黔金丝猴与喜马拉雅类群滇金丝猴、缅甸金丝猴分化时间约1.6 Ma [67 ] Ochotona 北半球 分布于青藏高原的高山类群和灌丛草原类群与低海拔泛北极分布类群分化时间大约为2.4 Ma [68 ] Apodemus draco 四川、云南、甘肃、青海、陕西、湖北、湖南、福建、台湾 2.2 Ma (3.8~0.87 Ma)分化形成,后来由于高原的阶段性隆升以及冰川作用种系及遗传结构更加复杂 [69 -70 ] Eothenomys 东亚 高原种类与低海拔种类分裂时间约2.7 Ma [71 ] Schizothoracinae 青藏高原及周边地区 在晚中新世晚期或上新世裂腹鱼亚科鱼类就已经出现在青藏高原及周边水系中,在晚更新世期间种内分化加速,不同海拔之间的类群产生等级分化,种间多样性增加,第四纪以来青藏高原隆升导致的水系改组是推动裂腹鱼亚科鱼类演化的原动力 [31 ,76 -78 ]
3 讨论 最近,青藏高原古地貌重建工作取得了丰硕的成果(图3 ),但是各种指标重建结果往往存在矛盾(图4 )[25 ,79 -83 ] .同位素重建结果表明高原南部在古近纪早期就存在安第斯型的巨大山系[84 -85 ] ;古生物研究发现高原腹地从中始新世到晚渐新世都存在东西向的大峡谷,该峡谷底部海拔在2 000 m左右,两侧为高大的冈底斯山脉和羌塘山脉[82 ,86 -88 ] ,动植物如巨犀(Paraceratherium )[89 ] 、攀鲈(Eoanabas )[90 -91 ] 、臭椿(Ailanthus )[88 ] 、椿榆(Cedrelospermum )[92 ] 、兔耳果(Lagokarpos )[93 ] 等在古近纪都可以通过青藏地区峡谷与周边动植物区系交流,新近纪之前的青藏地区虽然地形复杂,但是尚未隆升成今天的高原;横断山区某些区域则在晚始新世-早渐新世之交就已经获得现今的海拔高度[94 -95 ] ;高原北部和喜马拉雅山脉则在新近纪形成现在的地貌格局[25 ,96 -97 ] .古生物研究也发现青藏高原在晚上新世就已经存在高寒生境和冰缘环境,成为第四纪冰期动物忍耐寒冷环境的“训练营”,第四纪冰期来临之后这些适应寒冷环境的生物才扩散至北半球高纬度地区[34 ,98 -99 ] .因此,高原及周边地区高寒生物类群的起源时间表明在渐新世青藏高原部分区域就已经出现高寒生态系统,晚中新世以来青藏高原地貌与气候继续发生剧烈变化,促使这些物种的祖先类群迁入或就地演化积累适应高寒环境的基因和形态,并最终演化成适应高寒生境的新物种[25 ] .高寒区物种在晚上新世的加速分化可能与高原巨大山系和水系、冰川活动等造成的地理隔离有关[26 ] .随着第四纪冰期的到来,青藏高原及周边地区的高寒生物区也开始与北半球高纬度地区存在密切联系[34 ,98 -99 ] .同时,演化生物学研究也发现在第四纪冰期期间并未形成覆盖整个青藏高原的大冰盖,高原面上仍存在很多生物避难所[26 ] . ...
青藏高原琵甲族部分属的系统发育与历史生物地理学
4
2020
... 青藏高原及周边地区物种丰富,是全球生物多样性热点地区之一(图1 )[10 -11 ] .超过1 500属和12 000种植物生活在青藏高原地区,其中3 673种是该区域的特有种子植物[12 -14 ] .该地区植物区系独特,是全球海拔最高、高寒生物多样性最丰富的地区,被称为植物区系的“第三极”[15 ] .此外,青藏高原地区还生活着148种特有哺乳动物[16 ] ,167种特有鸟类[17 ] 以及六足动物亚门的许多特有种类[18 -19 ] .青藏高原平均海拔超过4 500 m,环境极端恶劣,低温、严寒、干旱、低氧、高紫外线辐射等是制约高寒区生物生存的关键因素,同时也是促使该区域生物特有种分化形成的主要原因[20 -21 ] .地质时期构造活动和气候变化可以形成地理隔离障碍促使物种分化和多样性增加,也可以通过减少生物扩散障碍使得物种分布区扩大,不同区域间的生物交流增加[22 -25 ] .青藏高原高寒区物种的形成和演化过程受到青藏高原构造抬升及其相应气候变化的影响[25 -26 ] .伴随着高原海拔的增长,物种就地演化或低海拔物种向高海拔迁移并适应高寒生境,或寒冷地区物种通过长距离扩散进入正在生长的高原[26 -27 ] ,导致该区域高寒生物多样性增加.高寒区生物多样性快速增长的时间通常同步于或略晚于高山或高原形成的时间[22 ,25 ,27 ] ,因此,高寒区生物类群的多样性演化过程和分化时间对我们认知构造活动期次[28 -29 ] ,推断高原隆升历史及其气候效应[1 ,30 -31 ] 等具有启示意义,甚至能够揭示以前所没有注意到的地质事件[22 ] .近年来,青藏高寒区生物地理学研究取得长足发展,为我们提供了一个独特的视角来探讨区域生物多样性演化、地质活动和气候变化之间的关系[25 ] . ...
... 生物地理学主要研究生物的时空分布格局以及性状演化过程及其形成原因和机制.从1852年开始,生物地理学家就开始寻找和总结动植物类群的分布模式,推断其历史分布区演化,并试图将这些分布模式与影响其演化的地质过程联系起来[35 ] .随着DNA测序技术的发展,研究者对标本进行DNA测序,基于宽松分子钟模型或使用年代和鉴定准确的化石记录来校正分子钟,使用最大似然法和贝叶斯法构建系统树,估算物种演化的分支时间和演化历史[18 ] ,通过现生类群的分布格局来推断祖先类群的地理分布.扩散-灭绝-进化分支(dispersal-extinction-cladogenesis)模型是目前最常使用的模型,使用最大似然法和贝叶斯法推断物种的扩散和灭绝历史,用隔离分化或支序发生来解释不同区域间类群分布格局[26 ,36 ] .同时,利用比较系统发育学方法还可以估算物种多样化速率,推断物种分布格局和多样性演化历史,探讨构造事件和气候变化对多样性形成和演化过程的影响[26 ] . ...
... 小琵甲属(Gnaptorina )是琵甲族的高原特有属,已知的34种全部分布在青藏高原及周边地区海拔3 000~4 500 m的区域,该属昆虫后翅愈合,不能飞翔,是对高海拔极端环境的一种适应性状,小琵甲属生物地理学研究表明其祖先类群在始新世起源于青藏高原东南缘,高原隆升和河流切割造成的地理隔离阻断了小琵甲属物种或种群之间的基因交流,加速特有种的快速辐射演化[18 -19 ] ;隙蛛(Coelotinae )是北半球温带和亚热带地区常见蜘蛛种类,青藏高原东南缘晚始新世形成的高海拔地貌阻碍了南方隙蛛向北扩散,但是随之形成的复杂地貌创造的局部地理隔离也促使南方隙蛛在此阶段快速辐射演化,渐新世以来高原中部和北部的隆起导致北方隙蛛在早期多样化过程中经历了灭绝事件[59 ] . ...
... Biological evolution in the alpine region of Tibetan Plateau
Table 1 种属 分布区域 适应高海拔类群分化时间或多样性增加时间 文献来源 Sinopodophyllum hexandrum 青藏高原东部 在约(6.52±1.89) Ma期间出现在青藏高原东部,但是直到3~2 Ma以来才演化出适应高山环境的形态 [47 ] Nannoglottis 青藏高原 在23~32 Ma期间出现在青藏高原,随后多样性增高,其高山灌丛和针叶林生活类型分支时间约为3.4 Ma [48 ] Reaumuria soongarica 西北干旱区、青藏高原北部 高原内部分布类群演化时间2.0~6.3 Ma,高原东北缘与高原西北类群分化时间1.1~3.5 Ma [49 ] Orchidinae 青藏高原及喜马拉雅山脉 青藏高原特有种大多是在过去6 Ma里进化而来 [50 ] Oxyria sinensis 青藏高原东南部 高山类群出现时间7.54~24.36 Ma,种内多样化增加时间1.74~0.86 Ma [51 ] Juniperus microsperma 西藏波密 早中新世21.6~14 Ma期间与其他种群分化 [52 ] Rhododendron 北半球 横断山区概属多样性在晚中新世(约8 Ma)快速增加 [53 ] Roscoea, Cautleya 喜马拉雅-横断山区 在早渐新世(~32 Ma)两属出现,Roscoea在横断山区与喜马拉雅山脉的两个分支大概在渐新世-中新世之交分化 [54 ] Nanorana parkeri 青藏高原南部 高原东部和西部类群分化时间1.4~3.7 Ma [55 ] Paini 南亚、东南亚、青藏高原 在渐新世早期棘蛙族高原-喜马拉雅类群与中国第二阶梯上生活的类群分化,随后在在26.1~12.5 Ma高原生活类群与喜马拉雅山脉生活类群分化,高原内部生活类群在晚中新世到上新世期间分化速率加快 [22 ] Bufo tibetanus 青藏高原、四川、甘肃、陕西等 高原种类形成于上新世期间 [56 ] Phrynocephalus 亚洲干旱区 在7.2~13.0 Ma期间演化出胎生类群,3.2~7.3 Ma期间种间多样性增加 [57 ] Phrynocephalus erythrurus 羌塘盆地 在3.67 Ma左右北部类群与南部类群发生分化,约2.76 Ma北部类群内部进一步分化 [58 ] Gnaptorina 青藏高原及周边高海拔地区 始新世起源于青藏高原东南缘 [18 -19 ] Coelotinae 北半球温带、亚热带 晚始新世高原东南缘隆升阻碍南方隙蛛向北扩散,渐新世高原中部和北部隆起导致北方隙蛛早期多样化进程中的灭绝事件 [59 ] Montifringilla, Onychostruthus, Pyrgilauda 青藏高原 雪雀的共同祖先在约14 Ma就应经生活在青藏高原,并进化出适应高海拔极端环境的性状 [21 ] Parus major 亚洲、欧洲 蒙古分布类群与其他种类分化时间为2.8~0.7 Ma,喜马拉雅山脉东部与中国中东部类群分化时间为1.9~0.4 Ma [60 ] Pseudopodoces humilis 青藏高原、柴达木盆地周边高山 在9.9~7.7 Ma期间地山雀就与与其他山雀分化开来,并出现适应高海拔性状 [61 ] Phylloscopus, Seicercus 亚洲 喜马拉雅山脉分布类群在7 Ma加速进化 [62 -63 ] Perdix 西藏、甘肃、青海、四川、新疆 高原类群出现于3.63 (4.52~2.75) Ma [64 ] Pantholops hodgsonii 西藏、新疆、青海 上新世-更新世就已经出现在青藏高原 [65 ] Procapra picticaudata 西藏、四川、甘肃、青海、新疆 种内分化时间约2.2~4.4 Ma [66 ] Rhinopithecus 四川、甘肃、陕西、贵州、云南、缅甸、越南 北方类群川金丝猴、黔金丝猴与喜马拉雅类群滇金丝猴、缅甸金丝猴分化时间约1.6 Ma [67 ] Ochotona 北半球 分布于青藏高原的高山类群和灌丛草原类群与低海拔泛北极分布类群分化时间大约为2.4 Ma [68 ] Apodemus draco 四川、云南、甘肃、青海、陕西、湖北、湖南、福建、台湾 2.2 Ma (3.8~0.87 Ma)分化形成,后来由于高原的阶段性隆升以及冰川作用种系及遗传结构更加复杂 [69 -70 ] Eothenomys 东亚 高原种类与低海拔种类分裂时间约2.7 Ma [71 ] Schizothoracinae 青藏高原及周边地区 在晚中新世晚期或上新世裂腹鱼亚科鱼类就已经出现在青藏高原及周边水系中,在晚更新世期间种内分化加速,不同海拔之间的类群产生等级分化,种间多样性增加,第四纪以来青藏高原隆升导致的水系改组是推动裂腹鱼亚科鱼类演化的原动力 [31 ,76 -78 ]
3 讨论 最近,青藏高原古地貌重建工作取得了丰硕的成果(图3 ),但是各种指标重建结果往往存在矛盾(图4 )[25 ,79 -83 ] .同位素重建结果表明高原南部在古近纪早期就存在安第斯型的巨大山系[84 -85 ] ;古生物研究发现高原腹地从中始新世到晚渐新世都存在东西向的大峡谷,该峡谷底部海拔在2 000 m左右,两侧为高大的冈底斯山脉和羌塘山脉[82 ,86 -88 ] ,动植物如巨犀(Paraceratherium )[89 ] 、攀鲈(Eoanabas )[90 -91 ] 、臭椿(Ailanthus )[88 ] 、椿榆(Cedrelospermum )[92 ] 、兔耳果(Lagokarpos )[93 ] 等在古近纪都可以通过青藏地区峡谷与周边动植物区系交流,新近纪之前的青藏地区虽然地形复杂,但是尚未隆升成今天的高原;横断山区某些区域则在晚始新世-早渐新世之交就已经获得现今的海拔高度[94 -95 ] ;高原北部和喜马拉雅山脉则在新近纪形成现在的地貌格局[25 ,96 -97 ] .古生物研究也发现青藏高原在晚上新世就已经存在高寒生境和冰缘环境,成为第四纪冰期动物忍耐寒冷环境的“训练营”,第四纪冰期来临之后这些适应寒冷环境的生物才扩散至北半球高纬度地区[34 ,98 -99 ] .因此,高原及周边地区高寒生物类群的起源时间表明在渐新世青藏高原部分区域就已经出现高寒生态系统,晚中新世以来青藏高原地貌与气候继续发生剧烈变化,促使这些物种的祖先类群迁入或就地演化积累适应高寒环境的基因和形态,并最终演化成适应高寒生境的新物种[25 ] .高寒区物种在晚上新世的加速分化可能与高原巨大山系和水系、冰川活动等造成的地理隔离有关[26 ] .随着第四纪冰期的到来,青藏高原及周边地区的高寒生物区也开始与北半球高纬度地区存在密切联系[34 ,98 -99 ] .同时,演化生物学研究也发现在第四纪冰期期间并未形成覆盖整个青藏高原的大冰盖,高原面上仍存在很多生物避难所[26 ] . ...
Phylogenetics, historical biogeography and molecular species delimitation of Gnaptorina Reitter (Coleoptera: Tenebrionidae: Blaptini)
3
2021
... 青藏高原及周边地区物种丰富,是全球生物多样性热点地区之一(图1 )[10 -11 ] .超过1 500属和12 000种植物生活在青藏高原地区,其中3 673种是该区域的特有种子植物[12 -14 ] .该地区植物区系独特,是全球海拔最高、高寒生物多样性最丰富的地区,被称为植物区系的“第三极”[15 ] .此外,青藏高原地区还生活着148种特有哺乳动物[16 ] ,167种特有鸟类[17 ] 以及六足动物亚门的许多特有种类[18 -19 ] .青藏高原平均海拔超过4 500 m,环境极端恶劣,低温、严寒、干旱、低氧、高紫外线辐射等是制约高寒区生物生存的关键因素,同时也是促使该区域生物特有种分化形成的主要原因[20 -21 ] .地质时期构造活动和气候变化可以形成地理隔离障碍促使物种分化和多样性增加,也可以通过减少生物扩散障碍使得物种分布区扩大,不同区域间的生物交流增加[22 -25 ] .青藏高原高寒区物种的形成和演化过程受到青藏高原构造抬升及其相应气候变化的影响[25 -26 ] .伴随着高原海拔的增长,物种就地演化或低海拔物种向高海拔迁移并适应高寒生境,或寒冷地区物种通过长距离扩散进入正在生长的高原[26 -27 ] ,导致该区域高寒生物多样性增加.高寒区生物多样性快速增长的时间通常同步于或略晚于高山或高原形成的时间[22 ,25 ,27 ] ,因此,高寒区生物类群的多样性演化过程和分化时间对我们认知构造活动期次[28 -29 ] ,推断高原隆升历史及其气候效应[1 ,30 -31 ] 等具有启示意义,甚至能够揭示以前所没有注意到的地质事件[22 ] .近年来,青藏高寒区生物地理学研究取得长足发展,为我们提供了一个独特的视角来探讨区域生物多样性演化、地质活动和气候变化之间的关系[25 ] . ...
... 小琵甲属(Gnaptorina )是琵甲族的高原特有属,已知的34种全部分布在青藏高原及周边地区海拔3 000~4 500 m的区域,该属昆虫后翅愈合,不能飞翔,是对高海拔极端环境的一种适应性状,小琵甲属生物地理学研究表明其祖先类群在始新世起源于青藏高原东南缘,高原隆升和河流切割造成的地理隔离阻断了小琵甲属物种或种群之间的基因交流,加速特有种的快速辐射演化[18 -19 ] ;隙蛛(Coelotinae )是北半球温带和亚热带地区常见蜘蛛种类,青藏高原东南缘晚始新世形成的高海拔地貌阻碍了南方隙蛛向北扩散,但是随之形成的复杂地貌创造的局部地理隔离也促使南方隙蛛在此阶段快速辐射演化,渐新世以来高原中部和北部的隆起导致北方隙蛛在早期多样化过程中经历了灭绝事件[59 ] . ...
... Biological evolution in the alpine region of Tibetan Plateau
Table 1 种属 分布区域 适应高海拔类群分化时间或多样性增加时间 文献来源 Sinopodophyllum hexandrum 青藏高原东部 在约(6.52±1.89) Ma期间出现在青藏高原东部,但是直到3~2 Ma以来才演化出适应高山环境的形态 [47 ] Nannoglottis 青藏高原 在23~32 Ma期间出现在青藏高原,随后多样性增高,其高山灌丛和针叶林生活类型分支时间约为3.4 Ma [48 ] Reaumuria soongarica 西北干旱区、青藏高原北部 高原内部分布类群演化时间2.0~6.3 Ma,高原东北缘与高原西北类群分化时间1.1~3.5 Ma [49 ] Orchidinae 青藏高原及喜马拉雅山脉 青藏高原特有种大多是在过去6 Ma里进化而来 [50 ] Oxyria sinensis 青藏高原东南部 高山类群出现时间7.54~24.36 Ma,种内多样化增加时间1.74~0.86 Ma [51 ] Juniperus microsperma 西藏波密 早中新世21.6~14 Ma期间与其他种群分化 [52 ] Rhododendron 北半球 横断山区概属多样性在晚中新世(约8 Ma)快速增加 [53 ] Roscoea, Cautleya 喜马拉雅-横断山区 在早渐新世(~32 Ma)两属出现,Roscoea在横断山区与喜马拉雅山脉的两个分支大概在渐新世-中新世之交分化 [54 ] Nanorana parkeri 青藏高原南部 高原东部和西部类群分化时间1.4~3.7 Ma [55 ] Paini 南亚、东南亚、青藏高原 在渐新世早期棘蛙族高原-喜马拉雅类群与中国第二阶梯上生活的类群分化,随后在在26.1~12.5 Ma高原生活类群与喜马拉雅山脉生活类群分化,高原内部生活类群在晚中新世到上新世期间分化速率加快 [22 ] Bufo tibetanus 青藏高原、四川、甘肃、陕西等 高原种类形成于上新世期间 [56 ] Phrynocephalus 亚洲干旱区 在7.2~13.0 Ma期间演化出胎生类群,3.2~7.3 Ma期间种间多样性增加 [57 ] Phrynocephalus erythrurus 羌塘盆地 在3.67 Ma左右北部类群与南部类群发生分化,约2.76 Ma北部类群内部进一步分化 [58 ] Gnaptorina 青藏高原及周边高海拔地区 始新世起源于青藏高原东南缘 [18 -19 ] Coelotinae 北半球温带、亚热带 晚始新世高原东南缘隆升阻碍南方隙蛛向北扩散,渐新世高原中部和北部隆起导致北方隙蛛早期多样化进程中的灭绝事件 [59 ] Montifringilla, Onychostruthus, Pyrgilauda 青藏高原 雪雀的共同祖先在约14 Ma就应经生活在青藏高原,并进化出适应高海拔极端环境的性状 [21 ] Parus major 亚洲、欧洲 蒙古分布类群与其他种类分化时间为2.8~0.7 Ma,喜马拉雅山脉东部与中国中东部类群分化时间为1.9~0.4 Ma [60 ] Pseudopodoces humilis 青藏高原、柴达木盆地周边高山 在9.9~7.7 Ma期间地山雀就与与其他山雀分化开来,并出现适应高海拔性状 [61 ] Phylloscopus, Seicercus 亚洲 喜马拉雅山脉分布类群在7 Ma加速进化 [62 -63 ] Perdix 西藏、甘肃、青海、四川、新疆 高原类群出现于3.63 (4.52~2.75) Ma [64 ] Pantholops hodgsonii 西藏、新疆、青海 上新世-更新世就已经出现在青藏高原 [65 ] Procapra picticaudata 西藏、四川、甘肃、青海、新疆 种内分化时间约2.2~4.4 Ma [66 ] Rhinopithecus 四川、甘肃、陕西、贵州、云南、缅甸、越南 北方类群川金丝猴、黔金丝猴与喜马拉雅类群滇金丝猴、缅甸金丝猴分化时间约1.6 Ma [67 ] Ochotona 北半球 分布于青藏高原的高山类群和灌丛草原类群与低海拔泛北极分布类群分化时间大约为2.4 Ma [68 ] Apodemus draco 四川、云南、甘肃、青海、陕西、湖北、湖南、福建、台湾 2.2 Ma (3.8~0.87 Ma)分化形成,后来由于高原的阶段性隆升以及冰川作用种系及遗传结构更加复杂 [69 -70 ] Eothenomys 东亚 高原种类与低海拔种类分裂时间约2.7 Ma [71 ] Schizothoracinae 青藏高原及周边地区 在晚中新世晚期或上新世裂腹鱼亚科鱼类就已经出现在青藏高原及周边水系中,在晚更新世期间种内分化加速,不同海拔之间的类群产生等级分化,种间多样性增加,第四纪以来青藏高原隆升导致的水系改组是推动裂腹鱼亚科鱼类演化的原动力 [31 ,76 -78 ]
3 讨论 最近,青藏高原古地貌重建工作取得了丰硕的成果(图3 ),但是各种指标重建结果往往存在矛盾(图4 )[25 ,79 -83 ] .同位素重建结果表明高原南部在古近纪早期就存在安第斯型的巨大山系[84 -85 ] ;古生物研究发现高原腹地从中始新世到晚渐新世都存在东西向的大峡谷,该峡谷底部海拔在2 000 m左右,两侧为高大的冈底斯山脉和羌塘山脉[82 ,86 -88 ] ,动植物如巨犀(Paraceratherium )[89 ] 、攀鲈(Eoanabas )[90 -91 ] 、臭椿(Ailanthus )[88 ] 、椿榆(Cedrelospermum )[92 ] 、兔耳果(Lagokarpos )[93 ] 等在古近纪都可以通过青藏地区峡谷与周边动植物区系交流,新近纪之前的青藏地区虽然地形复杂,但是尚未隆升成今天的高原;横断山区某些区域则在晚始新世-早渐新世之交就已经获得现今的海拔高度[94 -95 ] ;高原北部和喜马拉雅山脉则在新近纪形成现在的地貌格局[25 ,96 -97 ] .古生物研究也发现青藏高原在晚上新世就已经存在高寒生境和冰缘环境,成为第四纪冰期动物忍耐寒冷环境的“训练营”,第四纪冰期来临之后这些适应寒冷环境的生物才扩散至北半球高纬度地区[34 ,98 -99 ] .因此,高原及周边地区高寒生物类群的起源时间表明在渐新世青藏高原部分区域就已经出现高寒生态系统,晚中新世以来青藏高原地貌与气候继续发生剧烈变化,促使这些物种的祖先类群迁入或就地演化积累适应高寒环境的基因和形态,并最终演化成适应高寒生境的新物种[25 ] .高寒区物种在晚上新世的加速分化可能与高原巨大山系和水系、冰川活动等造成的地理隔离有关[26 ] .随着第四纪冰期的到来,青藏高原及周边地区的高寒生物区也开始与北半球高纬度地区存在密切联系[34 ,98 -99 ] .同时,演化生物学研究也发现在第四纪冰期期间并未形成覆盖整个青藏高原的大冰盖,高原面上仍存在很多生物避难所[26 ] . ...
Evolution of Neogene mammals in Eurasia: environmental forcing and biotic interactions
2
2014
... 青藏高原及周边地区物种丰富,是全球生物多样性热点地区之一(图1 )[10 -11 ] .超过1 500属和12 000种植物生活在青藏高原地区,其中3 673种是该区域的特有种子植物[12 -14 ] .该地区植物区系独特,是全球海拔最高、高寒生物多样性最丰富的地区,被称为植物区系的“第三极”[15 ] .此外,青藏高原地区还生活着148种特有哺乳动物[16 ] ,167种特有鸟类[17 ] 以及六足动物亚门的许多特有种类[18 -19 ] .青藏高原平均海拔超过4 500 m,环境极端恶劣,低温、严寒、干旱、低氧、高紫外线辐射等是制约高寒区生物生存的关键因素,同时也是促使该区域生物特有种分化形成的主要原因[20 -21 ] .地质时期构造活动和气候变化可以形成地理隔离障碍促使物种分化和多样性增加,也可以通过减少生物扩散障碍使得物种分布区扩大,不同区域间的生物交流增加[22 -25 ] .青藏高原高寒区物种的形成和演化过程受到青藏高原构造抬升及其相应气候变化的影响[25 -26 ] .伴随着高原海拔的增长,物种就地演化或低海拔物种向高海拔迁移并适应高寒生境,或寒冷地区物种通过长距离扩散进入正在生长的高原[26 -27 ] ,导致该区域高寒生物多样性增加.高寒区生物多样性快速增长的时间通常同步于或略晚于高山或高原形成的时间[22 ,25 ,27 ] ,因此,高寒区生物类群的多样性演化过程和分化时间对我们认知构造活动期次[28 -29 ] ,推断高原隆升历史及其气候效应[1 ,30 -31 ] 等具有启示意义,甚至能够揭示以前所没有注意到的地质事件[22 ] .近年来,青藏高寒区生物地理学研究取得长足发展,为我们提供了一个独特的视角来探讨区域生物多样性演化、地质活动和气候变化之间的关系[25 ] . ...
... 高寒生物区是分布于树线和雪线之间的独特生态系统,遍布全球各大洲高山和高原之上[26 ,32 ] .高寒生物区自然环境极端恶劣,物种为适应环境进化出独特的抗寒、抗旱、耐低温、抗紫外线辐射、抗缺氧等性状,并在这些极端环境中经过悠久的演化历史最终存活下来[21 ,26 ] ,是获取抗性基因的重要宝库.高寒区生物是适应极端环境的产物,对环境变化非常敏感.近年来全球变暖加剧了高寒区生物灭绝风险,使得该区域成为全球生物多样性保护研究中重点关注的区域.同时,高寒区生物是山脉或高原隆升到一定海拔后的产物[33 ] ,研究高寒区生物多样性的起源和演化过程,不仅可以认识高寒区生物多样性的形成过程,还可以预测未来气候变化对高寒区生物多样性的影响,同时也能够推断高山、高原地貌和气候的演化历史[20 ,23 -24 ,34 ] . ...
The evolution of ancestral and species-specific adaptations in snowfinches at the Qinghai-Tibet Plateau
4
2021
... 青藏高原及周边地区物种丰富,是全球生物多样性热点地区之一(图1 )[10 -11 ] .超过1 500属和12 000种植物生活在青藏高原地区,其中3 673种是该区域的特有种子植物[12 -14 ] .该地区植物区系独特,是全球海拔最高、高寒生物多样性最丰富的地区,被称为植物区系的“第三极”[15 ] .此外,青藏高原地区还生活着148种特有哺乳动物[16 ] ,167种特有鸟类[17 ] 以及六足动物亚门的许多特有种类[18 -19 ] .青藏高原平均海拔超过4 500 m,环境极端恶劣,低温、严寒、干旱、低氧、高紫外线辐射等是制约高寒区生物生存的关键因素,同时也是促使该区域生物特有种分化形成的主要原因[20 -21 ] .地质时期构造活动和气候变化可以形成地理隔离障碍促使物种分化和多样性增加,也可以通过减少生物扩散障碍使得物种分布区扩大,不同区域间的生物交流增加[22 -25 ] .青藏高原高寒区物种的形成和演化过程受到青藏高原构造抬升及其相应气候变化的影响[25 -26 ] .伴随着高原海拔的增长,物种就地演化或低海拔物种向高海拔迁移并适应高寒生境,或寒冷地区物种通过长距离扩散进入正在生长的高原[26 -27 ] ,导致该区域高寒生物多样性增加.高寒区生物多样性快速增长的时间通常同步于或略晚于高山或高原形成的时间[22 ,25 ,27 ] ,因此,高寒区生物类群的多样性演化过程和分化时间对我们认知构造活动期次[28 -29 ] ,推断高原隆升历史及其气候效应[1 ,30 -31 ] 等具有启示意义,甚至能够揭示以前所没有注意到的地质事件[22 ] .近年来,青藏高寒区生物地理学研究取得长足发展,为我们提供了一个独特的视角来探讨区域生物多样性演化、地质活动和气候变化之间的关系[25 ] . ...
... 高寒生物区是分布于树线和雪线之间的独特生态系统,遍布全球各大洲高山和高原之上[26 ,32 ] .高寒生物区自然环境极端恶劣,物种为适应环境进化出独特的抗寒、抗旱、耐低温、抗紫外线辐射、抗缺氧等性状,并在这些极端环境中经过悠久的演化历史最终存活下来[21 ,26 ] ,是获取抗性基因的重要宝库.高寒区生物是适应极端环境的产物,对环境变化非常敏感.近年来全球变暖加剧了高寒区生物灭绝风险,使得该区域成为全球生物多样性保护研究中重点关注的区域.同时,高寒区生物是山脉或高原隆升到一定海拔后的产物[33 ] ,研究高寒区生物多样性的起源和演化过程,不仅可以认识高寒区生物多样性的形成过程,还可以预测未来气候变化对高寒区生物多样性的影响,同时也能够推断高山、高原地貌和气候的演化历史[20 ,23 -24 ,34 ] . ...
... 鸟类中的雪雀(Montifringilla , Onychostruthus , Pyrgilauda )是在青藏高原低温、缺氧、强紫外线极端环境下繁衍壮大的类群,其全基因组测序与蛋白质功能研究发现雪雀的共同祖先在约14 Ma就应经生活在青藏高原,并进化出适应高海拔极端环境的性状[21 ] .大山雀(Parus major )在北半球广泛分布,第四纪以来高原周边地区构造隆升以及全球气候变干等形成地理隔离,使得高原分布类群与周边地区分布的姐妹类群发生分化,多样性升高[60 ] .地山雀(Pseudopodoces humilis )在青藏高原分布的海拔较高,一般超过4 000 m,系统发育研究发现在9.9~7.7 Ma期间地山雀就与其他山雀分化开来,并进化出适应高海拔生境的形态和习性[61 ] .对喜马拉雅山脉鸣禽的研究表明,山脉隆升和季风演化等原因导致柳莺属(Phylloscopus )和鹟莺属(Seicercus )在7 Ma左右开始加速分化[62 -63 ] .山鹑属(Perdix )广泛分布于中国西北和西南地区,其中高原山鹑(P. hodgsoniae )分布于青藏高原海拔2 500~5 000 m范围内,系统发育研究结果表明其适应高原生境的类群出现于4.52~2.75 Ma期间[64 ] . ...
... Biological evolution in the alpine region of Tibetan Plateau
Table 1 种属 分布区域 适应高海拔类群分化时间或多样性增加时间 文献来源 Sinopodophyllum hexandrum 青藏高原东部 在约(6.52±1.89) Ma期间出现在青藏高原东部,但是直到3~2 Ma以来才演化出适应高山环境的形态 [47 ] Nannoglottis 青藏高原 在23~32 Ma期间出现在青藏高原,随后多样性增高,其高山灌丛和针叶林生活类型分支时间约为3.4 Ma [48 ] Reaumuria soongarica 西北干旱区、青藏高原北部 高原内部分布类群演化时间2.0~6.3 Ma,高原东北缘与高原西北类群分化时间1.1~3.5 Ma [49 ] Orchidinae 青藏高原及喜马拉雅山脉 青藏高原特有种大多是在过去6 Ma里进化而来 [50 ] Oxyria sinensis 青藏高原东南部 高山类群出现时间7.54~24.36 Ma,种内多样化增加时间1.74~0.86 Ma [51 ] Juniperus microsperma 西藏波密 早中新世21.6~14 Ma期间与其他种群分化 [52 ] Rhododendron 北半球 横断山区概属多样性在晚中新世(约8 Ma)快速增加 [53 ] Roscoea, Cautleya 喜马拉雅-横断山区 在早渐新世(~32 Ma)两属出现,Roscoea在横断山区与喜马拉雅山脉的两个分支大概在渐新世-中新世之交分化 [54 ] Nanorana parkeri 青藏高原南部 高原东部和西部类群分化时间1.4~3.7 Ma [55 ] Paini 南亚、东南亚、青藏高原 在渐新世早期棘蛙族高原-喜马拉雅类群与中国第二阶梯上生活的类群分化,随后在在26.1~12.5 Ma高原生活类群与喜马拉雅山脉生活类群分化,高原内部生活类群在晚中新世到上新世期间分化速率加快 [22 ] Bufo tibetanus 青藏高原、四川、甘肃、陕西等 高原种类形成于上新世期间 [56 ] Phrynocephalus 亚洲干旱区 在7.2~13.0 Ma期间演化出胎生类群,3.2~7.3 Ma期间种间多样性增加 [57 ] Phrynocephalus erythrurus 羌塘盆地 在3.67 Ma左右北部类群与南部类群发生分化,约2.76 Ma北部类群内部进一步分化 [58 ] Gnaptorina 青藏高原及周边高海拔地区 始新世起源于青藏高原东南缘 [18 -19 ] Coelotinae 北半球温带、亚热带 晚始新世高原东南缘隆升阻碍南方隙蛛向北扩散,渐新世高原中部和北部隆起导致北方隙蛛早期多样化进程中的灭绝事件 [59 ] Montifringilla, Onychostruthus, Pyrgilauda 青藏高原 雪雀的共同祖先在约14 Ma就应经生活在青藏高原,并进化出适应高海拔极端环境的性状 [21 ] Parus major 亚洲、欧洲 蒙古分布类群与其他种类分化时间为2.8~0.7 Ma,喜马拉雅山脉东部与中国中东部类群分化时间为1.9~0.4 Ma [60 ] Pseudopodoces humilis 青藏高原、柴达木盆地周边高山 在9.9~7.7 Ma期间地山雀就与与其他山雀分化开来,并出现适应高海拔性状 [61 ] Phylloscopus, Seicercus 亚洲 喜马拉雅山脉分布类群在7 Ma加速进化 [62 -63 ] Perdix 西藏、甘肃、青海、四川、新疆 高原类群出现于3.63 (4.52~2.75) Ma [64 ] Pantholops hodgsonii 西藏、新疆、青海 上新世-更新世就已经出现在青藏高原 [65 ] Procapra picticaudata 西藏、四川、甘肃、青海、新疆 种内分化时间约2.2~4.4 Ma [66 ] Rhinopithecus 四川、甘肃、陕西、贵州、云南、缅甸、越南 北方类群川金丝猴、黔金丝猴与喜马拉雅类群滇金丝猴、缅甸金丝猴分化时间约1.6 Ma [67 ] Ochotona 北半球 分布于青藏高原的高山类群和灌丛草原类群与低海拔泛北极分布类群分化时间大约为2.4 Ma [68 ] Apodemus draco 四川、云南、甘肃、青海、陕西、湖北、湖南、福建、台湾 2.2 Ma (3.8~0.87 Ma)分化形成,后来由于高原的阶段性隆升以及冰川作用种系及遗传结构更加复杂 [69 -70 ] Eothenomys 东亚 高原种类与低海拔种类分裂时间约2.7 Ma [71 ] Schizothoracinae 青藏高原及周边地区 在晚中新世晚期或上新世裂腹鱼亚科鱼类就已经出现在青藏高原及周边水系中,在晚更新世期间种内分化加速,不同海拔之间的类群产生等级分化,种间多样性增加,第四纪以来青藏高原隆升导致的水系改组是推动裂腹鱼亚科鱼类演化的原动力 [31 ,76 -78 ]
3 讨论 最近,青藏高原古地貌重建工作取得了丰硕的成果(图3 ),但是各种指标重建结果往往存在矛盾(图4 )[25 ,79 -83 ] .同位素重建结果表明高原南部在古近纪早期就存在安第斯型的巨大山系[84 -85 ] ;古生物研究发现高原腹地从中始新世到晚渐新世都存在东西向的大峡谷,该峡谷底部海拔在2 000 m左右,两侧为高大的冈底斯山脉和羌塘山脉[82 ,86 -88 ] ,动植物如巨犀(Paraceratherium )[89 ] 、攀鲈(Eoanabas )[90 -91 ] 、臭椿(Ailanthus )[88 ] 、椿榆(Cedrelospermum )[92 ] 、兔耳果(Lagokarpos )[93 ] 等在古近纪都可以通过青藏地区峡谷与周边动植物区系交流,新近纪之前的青藏地区虽然地形复杂,但是尚未隆升成今天的高原;横断山区某些区域则在晚始新世-早渐新世之交就已经获得现今的海拔高度[94 -95 ] ;高原北部和喜马拉雅山脉则在新近纪形成现在的地貌格局[25 ,96 -97 ] .古生物研究也发现青藏高原在晚上新世就已经存在高寒生境和冰缘环境,成为第四纪冰期动物忍耐寒冷环境的“训练营”,第四纪冰期来临之后这些适应寒冷环境的生物才扩散至北半球高纬度地区[34 ,98 -99 ] .因此,高原及周边地区高寒生物类群的起源时间表明在渐新世青藏高原部分区域就已经出现高寒生态系统,晚中新世以来青藏高原地貌与气候继续发生剧烈变化,促使这些物种的祖先类群迁入或就地演化积累适应高寒环境的基因和形态,并最终演化成适应高寒生境的新物种[25 ] .高寒区物种在晚上新世的加速分化可能与高原巨大山系和水系、冰川活动等造成的地理隔离有关[26 ] .随着第四纪冰期的到来,青藏高原及周边地区的高寒生物区也开始与北半球高纬度地区存在密切联系[34 ,98 -99 ] .同时,演化生物学研究也发现在第四纪冰期期间并未形成覆盖整个青藏高原的大冰盖,高原面上仍存在很多生物避难所[26 ] . ...
Spiny frogs (Paini) illuminate the history of the Himalayan region and Southeast Asia
5
2010
... 青藏高原及周边地区物种丰富,是全球生物多样性热点地区之一(图1 )[10 -11 ] .超过1 500属和12 000种植物生活在青藏高原地区,其中3 673种是该区域的特有种子植物[12 -14 ] .该地区植物区系独特,是全球海拔最高、高寒生物多样性最丰富的地区,被称为植物区系的“第三极”[15 ] .此外,青藏高原地区还生活着148种特有哺乳动物[16 ] ,167种特有鸟类[17 ] 以及六足动物亚门的许多特有种类[18 -19 ] .青藏高原平均海拔超过4 500 m,环境极端恶劣,低温、严寒、干旱、低氧、高紫外线辐射等是制约高寒区生物生存的关键因素,同时也是促使该区域生物特有种分化形成的主要原因[20 -21 ] .地质时期构造活动和气候变化可以形成地理隔离障碍促使物种分化和多样性增加,也可以通过减少生物扩散障碍使得物种分布区扩大,不同区域间的生物交流增加[22 -25 ] .青藏高原高寒区物种的形成和演化过程受到青藏高原构造抬升及其相应气候变化的影响[25 -26 ] .伴随着高原海拔的增长,物种就地演化或低海拔物种向高海拔迁移并适应高寒生境,或寒冷地区物种通过长距离扩散进入正在生长的高原[26 -27 ] ,导致该区域高寒生物多样性增加.高寒区生物多样性快速增长的时间通常同步于或略晚于高山或高原形成的时间[22 ,25 ,27 ] ,因此,高寒区生物类群的多样性演化过程和分化时间对我们认知构造活动期次[28 -29 ] ,推断高原隆升历史及其气候效应[1 ,30 -31 ] 等具有启示意义,甚至能够揭示以前所没有注意到的地质事件[22 ] .近年来,青藏高寒区生物地理学研究取得长足发展,为我们提供了一个独特的视角来探讨区域生物多样性演化、地质活动和气候变化之间的关系[25 ] . ...
... [22 ,25 ,27 ],因此,高寒区生物类群的多样性演化过程和分化时间对我们认知构造活动期次[28 -29 ] ,推断高原隆升历史及其气候效应[1 ,30 -31 ] 等具有启示意义,甚至能够揭示以前所没有注意到的地质事件[22 ] .近年来,青藏高寒区生物地理学研究取得长足发展,为我们提供了一个独特的视角来探讨区域生物多样性演化、地质活动和气候变化之间的关系[25 ] . ...
... [22 ].近年来,青藏高寒区生物地理学研究取得长足发展,为我们提供了一个独特的视角来探讨区域生物多样性演化、地质活动和气候变化之间的关系[25 ] . ...
... 两栖动物中的高山倭蛙(Nanorana parkeri )分布于青藏高原南部海拔2 850~4 700 m之间的高山地区,其高原东部和西部类群分化时间约为3.7~1.4 Ma[55 ] .棘蛙族(Paini)物种多分布于深山急流中,结合其分子进化历史和生物地理分析发现在渐新世早期棘蛙族高原-喜马拉雅类群与中国第二阶梯上生活的类群分化,随后在26.1~12.5 Ma高原生活类群与喜马拉雅山脉生活类群分化,高原内部生活类群在晚中新世到上新世期间分化速率加快[22 ] .生活于青藏高原南部海拔2 850~4 700 m区域的西藏蟾蜍(Bufo tibetanus ),系统发育学研究表明其在上新世就演化出适应高原生活的性状[56 ] .爬行类动物沙蜥属(Phrynocephalus )一般分布于海拔超过2 200 m地区,为适应高原恶劣环境在约13.0~7.2 Ma进化出胎生类群,在7.3~3.2 Ma期间沙蜥属种间多样性快速增加[57 ] .分布于羌塘高原海拔4 500~5 000 m区域的红尾沙蜥(P.erythrurus )在3.67 Ma左右北部类群与南部类群发生分化,约2.76 Ma北部类群内部进一步分化,可能与晚上新世以来唐古拉山脉、乌兰乌拉山脉以及北麓河构造活动形成地理隔离有关[58 ] . ...
... Biological evolution in the alpine region of Tibetan Plateau
Table 1 种属 分布区域 适应高海拔类群分化时间或多样性增加时间 文献来源 Sinopodophyllum hexandrum 青藏高原东部 在约(6.52±1.89) Ma期间出现在青藏高原东部,但是直到3~2 Ma以来才演化出适应高山环境的形态 [47 ] Nannoglottis 青藏高原 在23~32 Ma期间出现在青藏高原,随后多样性增高,其高山灌丛和针叶林生活类型分支时间约为3.4 Ma [48 ] Reaumuria soongarica 西北干旱区、青藏高原北部 高原内部分布类群演化时间2.0~6.3 Ma,高原东北缘与高原西北类群分化时间1.1~3.5 Ma [49 ] Orchidinae 青藏高原及喜马拉雅山脉 青藏高原特有种大多是在过去6 Ma里进化而来 [50 ] Oxyria sinensis 青藏高原东南部 高山类群出现时间7.54~24.36 Ma,种内多样化增加时间1.74~0.86 Ma [51 ] Juniperus microsperma 西藏波密 早中新世21.6~14 Ma期间与其他种群分化 [52 ] Rhododendron 北半球 横断山区概属多样性在晚中新世(约8 Ma)快速增加 [53 ] Roscoea, Cautleya 喜马拉雅-横断山区 在早渐新世(~32 Ma)两属出现,Roscoea在横断山区与喜马拉雅山脉的两个分支大概在渐新世-中新世之交分化 [54 ] Nanorana parkeri 青藏高原南部 高原东部和西部类群分化时间1.4~3.7 Ma [55 ] Paini 南亚、东南亚、青藏高原 在渐新世早期棘蛙族高原-喜马拉雅类群与中国第二阶梯上生活的类群分化,随后在在26.1~12.5 Ma高原生活类群与喜马拉雅山脉生活类群分化,高原内部生活类群在晚中新世到上新世期间分化速率加快 [22 ] Bufo tibetanus 青藏高原、四川、甘肃、陕西等 高原种类形成于上新世期间 [56 ] Phrynocephalus 亚洲干旱区 在7.2~13.0 Ma期间演化出胎生类群,3.2~7.3 Ma期间种间多样性增加 [57 ] Phrynocephalus erythrurus 羌塘盆地 在3.67 Ma左右北部类群与南部类群发生分化,约2.76 Ma北部类群内部进一步分化 [58 ] Gnaptorina 青藏高原及周边高海拔地区 始新世起源于青藏高原东南缘 [18 -19 ] Coelotinae 北半球温带、亚热带 晚始新世高原东南缘隆升阻碍南方隙蛛向北扩散,渐新世高原中部和北部隆起导致北方隙蛛早期多样化进程中的灭绝事件 [59 ] Montifringilla, Onychostruthus, Pyrgilauda 青藏高原 雪雀的共同祖先在约14 Ma就应经生活在青藏高原,并进化出适应高海拔极端环境的性状 [21 ] Parus major 亚洲、欧洲 蒙古分布类群与其他种类分化时间为2.8~0.7 Ma,喜马拉雅山脉东部与中国中东部类群分化时间为1.9~0.4 Ma [60 ] Pseudopodoces humilis 青藏高原、柴达木盆地周边高山 在9.9~7.7 Ma期间地山雀就与与其他山雀分化开来,并出现适应高海拔性状 [61 ] Phylloscopus, Seicercus 亚洲 喜马拉雅山脉分布类群在7 Ma加速进化 [62 -63 ] Perdix 西藏、甘肃、青海、四川、新疆 高原类群出现于3.63 (4.52~2.75) Ma [64 ] Pantholops hodgsonii 西藏、新疆、青海 上新世-更新世就已经出现在青藏高原 [65 ] Procapra picticaudata 西藏、四川、甘肃、青海、新疆 种内分化时间约2.2~4.4 Ma [66 ] Rhinopithecus 四川、甘肃、陕西、贵州、云南、缅甸、越南 北方类群川金丝猴、黔金丝猴与喜马拉雅类群滇金丝猴、缅甸金丝猴分化时间约1.6 Ma [67 ] Ochotona 北半球 分布于青藏高原的高山类群和灌丛草原类群与低海拔泛北极分布类群分化时间大约为2.4 Ma [68 ] Apodemus draco 四川、云南、甘肃、青海、陕西、湖北、湖南、福建、台湾 2.2 Ma (3.8~0.87 Ma)分化形成,后来由于高原的阶段性隆升以及冰川作用种系及遗传结构更加复杂 [69 -70 ] Eothenomys 东亚 高原种类与低海拔种类分裂时间约2.7 Ma [71 ] Schizothoracinae 青藏高原及周边地区 在晚中新世晚期或上新世裂腹鱼亚科鱼类就已经出现在青藏高原及周边水系中,在晚更新世期间种内分化加速,不同海拔之间的类群产生等级分化,种间多样性增加,第四纪以来青藏高原隆升导致的水系改组是推动裂腹鱼亚科鱼类演化的原动力 [31 ,76 -78 ]
3 讨论 最近,青藏高原古地貌重建工作取得了丰硕的成果(图3 ),但是各种指标重建结果往往存在矛盾(图4 )[25 ,79 -83 ] .同位素重建结果表明高原南部在古近纪早期就存在安第斯型的巨大山系[84 -85 ] ;古生物研究发现高原腹地从中始新世到晚渐新世都存在东西向的大峡谷,该峡谷底部海拔在2 000 m左右,两侧为高大的冈底斯山脉和羌塘山脉[82 ,86 -88 ] ,动植物如巨犀(Paraceratherium )[89 ] 、攀鲈(Eoanabas )[90 -91 ] 、臭椿(Ailanthus )[88 ] 、椿榆(Cedrelospermum )[92 ] 、兔耳果(Lagokarpos )[93 ] 等在古近纪都可以通过青藏地区峡谷与周边动植物区系交流,新近纪之前的青藏地区虽然地形复杂,但是尚未隆升成今天的高原;横断山区某些区域则在晚始新世-早渐新世之交就已经获得现今的海拔高度[94 -95 ] ;高原北部和喜马拉雅山脉则在新近纪形成现在的地貌格局[25 ,96 -97 ] .古生物研究也发现青藏高原在晚上新世就已经存在高寒生境和冰缘环境,成为第四纪冰期动物忍耐寒冷环境的“训练营”,第四纪冰期来临之后这些适应寒冷环境的生物才扩散至北半球高纬度地区[34 ,98 -99 ] .因此,高原及周边地区高寒生物类群的起源时间表明在渐新世青藏高原部分区域就已经出现高寒生态系统,晚中新世以来青藏高原地貌与气候继续发生剧烈变化,促使这些物种的祖先类群迁入或就地演化积累适应高寒环境的基因和形态,并最终演化成适应高寒生境的新物种[25 ] .高寒区物种在晚上新世的加速分化可能与高原巨大山系和水系、冰川活动等造成的地理隔离有关[26 ] .随着第四纪冰期的到来,青藏高原及周边地区的高寒生物区也开始与北半球高纬度地区存在密切联系[34 ,98 -99 ] .同时,演化生物学研究也发现在第四纪冰期期间并未形成覆盖整个青藏高原的大冰盖,高原面上仍存在很多生物避难所[26 ] . ...
Geological and climatic influences on mountain biodiversity
1
2018
... 高寒生物区是分布于树线和雪线之间的独特生态系统,遍布全球各大洲高山和高原之上[26 ,32 ] .高寒生物区自然环境极端恶劣,物种为适应环境进化出独特的抗寒、抗旱、耐低温、抗紫外线辐射、抗缺氧等性状,并在这些极端环境中经过悠久的演化历史最终存活下来[21 ,26 ] ,是获取抗性基因的重要宝库.高寒区生物是适应极端环境的产物,对环境变化非常敏感.近年来全球变暖加剧了高寒区生物灭绝风险,使得该区域成为全球生物多样性保护研究中重点关注的区域.同时,高寒区生物是山脉或高原隆升到一定海拔后的产物[33 ] ,研究高寒区生物多样性的起源和演化过程,不仅可以认识高寒区生物多样性的形成过程,还可以预测未来气候变化对高寒区生物多样性的影响,同时也能够推断高山、高原地貌和气候的演化历史[20 ,23 -24 ,34 ] . ...
Building mountain biodiversity: geological and evolutionary processes
1
2019
... 高寒生物区是分布于树线和雪线之间的独特生态系统,遍布全球各大洲高山和高原之上[26 ,32 ] .高寒生物区自然环境极端恶劣,物种为适应环境进化出独特的抗寒、抗旱、耐低温、抗紫外线辐射、抗缺氧等性状,并在这些极端环境中经过悠久的演化历史最终存活下来[21 ,26 ] ,是获取抗性基因的重要宝库.高寒区生物是适应极端环境的产物,对环境变化非常敏感.近年来全球变暖加剧了高寒区生物灭绝风险,使得该区域成为全球生物多样性保护研究中重点关注的区域.同时,高寒区生物是山脉或高原隆升到一定海拔后的产物[33 ] ,研究高寒区生物多样性的起源和演化过程,不仅可以认识高寒区生物多样性的形成过程,还可以预测未来气候变化对高寒区生物多样性的影响,同时也能够推断高山、高原地貌和气候的演化历史[20 ,23 -24 ,34 ] . ...
晚新生代天水盆地孢粉记录的气候变化与青藏高原隆升
13
2016
... 青藏高原及周边地区物种丰富,是全球生物多样性热点地区之一(图1 )[10 -11 ] .超过1 500属和12 000种植物生活在青藏高原地区,其中3 673种是该区域的特有种子植物[12 -14 ] .该地区植物区系独特,是全球海拔最高、高寒生物多样性最丰富的地区,被称为植物区系的“第三极”[15 ] .此外,青藏高原地区还生活着148种特有哺乳动物[16 ] ,167种特有鸟类[17 ] 以及六足动物亚门的许多特有种类[18 -19 ] .青藏高原平均海拔超过4 500 m,环境极端恶劣,低温、严寒、干旱、低氧、高紫外线辐射等是制约高寒区生物生存的关键因素,同时也是促使该区域生物特有种分化形成的主要原因[20 -21 ] .地质时期构造活动和气候变化可以形成地理隔离障碍促使物种分化和多样性增加,也可以通过减少生物扩散障碍使得物种分布区扩大,不同区域间的生物交流增加[22 -25 ] .青藏高原高寒区物种的形成和演化过程受到青藏高原构造抬升及其相应气候变化的影响[25 -26 ] .伴随着高原海拔的增长,物种就地演化或低海拔物种向高海拔迁移并适应高寒生境,或寒冷地区物种通过长距离扩散进入正在生长的高原[26 -27 ] ,导致该区域高寒生物多样性增加.高寒区生物多样性快速增长的时间通常同步于或略晚于高山或高原形成的时间[22 ,25 ,27 ] ,因此,高寒区生物类群的多样性演化过程和分化时间对我们认知构造活动期次[28 -29 ] ,推断高原隆升历史及其气候效应[1 ,30 -31 ] 等具有启示意义,甚至能够揭示以前所没有注意到的地质事件[22 ] .近年来,青藏高寒区生物地理学研究取得长足发展,为我们提供了一个独特的视角来探讨区域生物多样性演化、地质活动和气候变化之间的关系[25 ] . ...
... [25 -26 ].伴随着高原海拔的增长,物种就地演化或低海拔物种向高海拔迁移并适应高寒生境,或寒冷地区物种通过长距离扩散进入正在生长的高原[26 -27 ] ,导致该区域高寒生物多样性增加.高寒区生物多样性快速增长的时间通常同步于或略晚于高山或高原形成的时间[22 ,25 ,27 ] ,因此,高寒区生物类群的多样性演化过程和分化时间对我们认知构造活动期次[28 -29 ] ,推断高原隆升历史及其气候效应[1 ,30 -31 ] 等具有启示意义,甚至能够揭示以前所没有注意到的地质事件[22 ] .近年来,青藏高寒区生物地理学研究取得长足发展,为我们提供了一个独特的视角来探讨区域生物多样性演化、地质活动和气候变化之间的关系[25 ] . ...
... ,25 ,27 ],因此,高寒区生物类群的多样性演化过程和分化时间对我们认知构造活动期次[28 -29 ] ,推断高原隆升历史及其气候效应[1 ,30 -31 ] 等具有启示意义,甚至能够揭示以前所没有注意到的地质事件[22 ] .近年来,青藏高寒区生物地理学研究取得长足发展,为我们提供了一个独特的视角来探讨区域生物多样性演化、地质活动和气候变化之间的关系[25 ] . ...
... [25 ]. ...
... 最近,青藏高原古地貌重建工作取得了丰硕的成果(图3 ),但是各种指标重建结果往往存在矛盾(图4 )[25 ,79 -83 ] .同位素重建结果表明高原南部在古近纪早期就存在安第斯型的巨大山系[84 -85 ] ;古生物研究发现高原腹地从中始新世到晚渐新世都存在东西向的大峡谷,该峡谷底部海拔在2 000 m左右,两侧为高大的冈底斯山脉和羌塘山脉[82 ,86 -88 ] ,动植物如巨犀(Paraceratherium )[89 ] 、攀鲈(Eoanabas )[90 -91 ] 、臭椿(Ailanthus )[88 ] 、椿榆(Cedrelospermum )[92 ] 、兔耳果(Lagokarpos )[93 ] 等在古近纪都可以通过青藏地区峡谷与周边动植物区系交流,新近纪之前的青藏地区虽然地形复杂,但是尚未隆升成今天的高原;横断山区某些区域则在晚始新世-早渐新世之交就已经获得现今的海拔高度[94 -95 ] ;高原北部和喜马拉雅山脉则在新近纪形成现在的地貌格局[25 ,96 -97 ] .古生物研究也发现青藏高原在晚上新世就已经存在高寒生境和冰缘环境,成为第四纪冰期动物忍耐寒冷环境的“训练营”,第四纪冰期来临之后这些适应寒冷环境的生物才扩散至北半球高纬度地区[34 ,98 -99 ] .因此,高原及周边地区高寒生物类群的起源时间表明在渐新世青藏高原部分区域就已经出现高寒生态系统,晚中新世以来青藏高原地貌与气候继续发生剧烈变化,促使这些物种的祖先类群迁入或就地演化积累适应高寒环境的基因和形态,并最终演化成适应高寒生境的新物种[25 ] .高寒区物种在晚上新世的加速分化可能与高原巨大山系和水系、冰川活动等造成的地理隔离有关[26 ] .随着第四纪冰期的到来,青藏高原及周边地区的高寒生物区也开始与北半球高纬度地区存在密切联系[34 ,98 -99 ] .同时,演化生物学研究也发现在第四纪冰期期间并未形成覆盖整个青藏高原的大冰盖,高原面上仍存在很多生物避难所[26 ] . ...
... [25 ,96 -97 ].古生物研究也发现青藏高原在晚上新世就已经存在高寒生境和冰缘环境,成为第四纪冰期动物忍耐寒冷环境的“训练营”,第四纪冰期来临之后这些适应寒冷环境的生物才扩散至北半球高纬度地区[34 ,98 -99 ] .因此,高原及周边地区高寒生物类群的起源时间表明在渐新世青藏高原部分区域就已经出现高寒生态系统,晚中新世以来青藏高原地貌与气候继续发生剧烈变化,促使这些物种的祖先类群迁入或就地演化积累适应高寒环境的基因和形态,并最终演化成适应高寒生境的新物种[25 ] .高寒区物种在晚上新世的加速分化可能与高原巨大山系和水系、冰川活动等造成的地理隔离有关[26 ] .随着第四纪冰期的到来,青藏高原及周边地区的高寒生物区也开始与北半球高纬度地区存在密切联系[34 ,98 -99 ] .同时,演化生物学研究也发现在第四纪冰期期间并未形成覆盖整个青藏高原的大冰盖,高原面上仍存在很多生物避难所[26 ] . ...
... [25 ].高寒区物种在晚上新世的加速分化可能与高原巨大山系和水系、冰川活动等造成的地理隔离有关[26 ] .随着第四纪冰期的到来,青藏高原及周边地区的高寒生物区也开始与北半球高纬度地区存在密切联系[34 ,98 -99 ] .同时,演化生物学研究也发现在第四纪冰期期间并未形成覆盖整个青藏高原的大冰盖,高原面上仍存在很多生物避难所[26 ] . ...
... 青藏高原新生代古高程定量重建研究点(根据文献[
25 ]修改)
Quantitative reconstruction paleoelevation sites on Tibetan Plateau (Revised from Reference [<xref ref-type="bibr" rid="R25">25</xref>]) Fig.3 ![]()
图4 青藏高原南北剖面古高程变化示意图(根据文献[<xref ref-type="bibr" rid="R25">25</xref>]、[<xref ref-type="bibr" rid="R34">34</xref>]、[<xref ref-type="bibr" rid="R82">82</xref>]、[<xref ref-type="bibr" rid="R87">87</xref>]、[<xref ref-type="bibr" rid="R100">100</xref>]修改) North-south paleoelevation profiles on the Tibetan Plateau (Revised from References [25],[34],[82],[87],[100]) Fig.4 ![]()
随着DNA测序技术和比较系统学方法的发展,大量涌现的青藏高寒区生物地理学研究为我们理解高原隆升过程及其气候效应提供了生物演化方面的证据[25 ] .但是,演化生物学家通常仅对单一特定类群的系统发育过程开展研究,很少进行系统的总结工作[25 ] ,对某一类群遗传数据的解释也往往与最新的青藏高原隆升和气候变化研究进展脱节,不能很好地将最新的关于高原隆升和气候变化的研究与生物类群演化联系在一起[101 ] ,同时也没有认识到青藏高原的隆升过程是阶段性的,没有对青藏高原的不同组成部分进行区分[26 ,53 ,81 ] .地质学研究大多认为高原地貌格局形成于中新世之前,但是综合近年来的生物地理学研究结果可以发现,青藏高寒生物类群的分化时间多集中在晚中新世,种内分化加速和多样性增加主要发生在晚上新世到第四纪期间(图5 ),这些不同类群在分化时间上的巧合可能反映青藏高寒区生物演化受到晚中新世以来构造和气候事件的影响,但是并未引起地质学家足够的重视[25 ] . ...
... Quantitative reconstruction paleoelevation sites on Tibetan Plateau (Revised from Reference [
25 ])
Fig.3 ![]()
图4 青藏高原南北剖面古高程变化示意图(根据文献[<xref ref-type="bibr" rid="R25">25</xref>]、[<xref ref-type="bibr" rid="R34">34</xref>]、[<xref ref-type="bibr" rid="R82">82</xref>]、[<xref ref-type="bibr" rid="R87">87</xref>]、[<xref ref-type="bibr" rid="R100">100</xref>]修改) North-south paleoelevation profiles on the Tibetan Plateau (Revised from References [25],[34],[82],[87],[100]) Fig.4 ![]()
随着DNA测序技术和比较系统学方法的发展,大量涌现的青藏高寒区生物地理学研究为我们理解高原隆升过程及其气候效应提供了生物演化方面的证据[25 ] .但是,演化生物学家通常仅对单一特定类群的系统发育过程开展研究,很少进行系统的总结工作[25 ] ,对某一类群遗传数据的解释也往往与最新的青藏高原隆升和气候变化研究进展脱节,不能很好地将最新的关于高原隆升和气候变化的研究与生物类群演化联系在一起[101 ] ,同时也没有认识到青藏高原的隆升过程是阶段性的,没有对青藏高原的不同组成部分进行区分[26 ,53 ,81 ] .地质学研究大多认为高原地貌格局形成于中新世之前,但是综合近年来的生物地理学研究结果可以发现,青藏高寒生物类群的分化时间多集中在晚中新世,种内分化加速和多样性增加主要发生在晚上新世到第四纪期间(图5 ),这些不同类群在分化时间上的巧合可能反映青藏高寒区生物演化受到晚中新世以来构造和气候事件的影响,但是并未引起地质学家足够的重视[25 ] . ...
... 青藏高原南北剖面古高程变化示意图(根据文献[
25 ]、[
34 ]、[
82 ]、[
87 ]、[
100 ]修改)
North-south paleoelevation profiles on the Tibetan Plateau (Revised from References [25],[34],[82],[87],[100]) Fig.4 ![]()
随着DNA测序技术和比较系统学方法的发展,大量涌现的青藏高寒区生物地理学研究为我们理解高原隆升过程及其气候效应提供了生物演化方面的证据[25 ] .但是,演化生物学家通常仅对单一特定类群的系统发育过程开展研究,很少进行系统的总结工作[25 ] ,对某一类群遗传数据的解释也往往与最新的青藏高原隆升和气候变化研究进展脱节,不能很好地将最新的关于高原隆升和气候变化的研究与生物类群演化联系在一起[101 ] ,同时也没有认识到青藏高原的隆升过程是阶段性的,没有对青藏高原的不同组成部分进行区分[26 ,53 ,81 ] .地质学研究大多认为高原地貌格局形成于中新世之前,但是综合近年来的生物地理学研究结果可以发现,青藏高寒生物类群的分化时间多集中在晚中新世,种内分化加速和多样性增加主要发生在晚上新世到第四纪期间(图5 ),这些不同类群在分化时间上的巧合可能反映青藏高寒区生物演化受到晚中新世以来构造和气候事件的影响,但是并未引起地质学家足够的重视[25 ] . ...
... 随着DNA测序技术和比较系统学方法的发展,大量涌现的青藏高寒区生物地理学研究为我们理解高原隆升过程及其气候效应提供了生物演化方面的证据[25 ] .但是,演化生物学家通常仅对单一特定类群的系统发育过程开展研究,很少进行系统的总结工作[25 ] ,对某一类群遗传数据的解释也往往与最新的青藏高原隆升和气候变化研究进展脱节,不能很好地将最新的关于高原隆升和气候变化的研究与生物类群演化联系在一起[101 ] ,同时也没有认识到青藏高原的隆升过程是阶段性的,没有对青藏高原的不同组成部分进行区分[26 ,53 ,81 ] .地质学研究大多认为高原地貌格局形成于中新世之前,但是综合近年来的生物地理学研究结果可以发现,青藏高寒生物类群的分化时间多集中在晚中新世,种内分化加速和多样性增加主要发生在晚上新世到第四纪期间(图5 ),这些不同类群在分化时间上的巧合可能反映青藏高寒区生物演化受到晚中新世以来构造和气候事件的影响,但是并未引起地质学家足够的重视[25 ] . ...
... [25 ],对某一类群遗传数据的解释也往往与最新的青藏高原隆升和气候变化研究进展脱节,不能很好地将最新的关于高原隆升和气候变化的研究与生物类群演化联系在一起[101 ] ,同时也没有认识到青藏高原的隆升过程是阶段性的,没有对青藏高原的不同组成部分进行区分[26 ,53 ,81 ] .地质学研究大多认为高原地貌格局形成于中新世之前,但是综合近年来的生物地理学研究结果可以发现,青藏高寒生物类群的分化时间多集中在晚中新世,种内分化加速和多样性增加主要发生在晚上新世到第四纪期间(图5 ),这些不同类群在分化时间上的巧合可能反映青藏高寒区生物演化受到晚中新世以来构造和气候事件的影响,但是并未引起地质学家足够的重视[25 ] . ...
... [25 ]. ...
晚新生代天水盆地孢粉记录的气候变化与青藏高原隆升
13
2016
... 青藏高原及周边地区物种丰富,是全球生物多样性热点地区之一(图1 )[10 -11 ] .超过1 500属和12 000种植物生活在青藏高原地区,其中3 673种是该区域的特有种子植物[12 -14 ] .该地区植物区系独特,是全球海拔最高、高寒生物多样性最丰富的地区,被称为植物区系的“第三极”[15 ] .此外,青藏高原地区还生活着148种特有哺乳动物[16 ] ,167种特有鸟类[17 ] 以及六足动物亚门的许多特有种类[18 -19 ] .青藏高原平均海拔超过4 500 m,环境极端恶劣,低温、严寒、干旱、低氧、高紫外线辐射等是制约高寒区生物生存的关键因素,同时也是促使该区域生物特有种分化形成的主要原因[20 -21 ] .地质时期构造活动和气候变化可以形成地理隔离障碍促使物种分化和多样性增加,也可以通过减少生物扩散障碍使得物种分布区扩大,不同区域间的生物交流增加[22 -25 ] .青藏高原高寒区物种的形成和演化过程受到青藏高原构造抬升及其相应气候变化的影响[25 -26 ] .伴随着高原海拔的增长,物种就地演化或低海拔物种向高海拔迁移并适应高寒生境,或寒冷地区物种通过长距离扩散进入正在生长的高原[26 -27 ] ,导致该区域高寒生物多样性增加.高寒区生物多样性快速增长的时间通常同步于或略晚于高山或高原形成的时间[22 ,25 ,27 ] ,因此,高寒区生物类群的多样性演化过程和分化时间对我们认知构造活动期次[28 -29 ] ,推断高原隆升历史及其气候效应[1 ,30 -31 ] 等具有启示意义,甚至能够揭示以前所没有注意到的地质事件[22 ] .近年来,青藏高寒区生物地理学研究取得长足发展,为我们提供了一个独特的视角来探讨区域生物多样性演化、地质活动和气候变化之间的关系[25 ] . ...
... [25 -26 ].伴随着高原海拔的增长,物种就地演化或低海拔物种向高海拔迁移并适应高寒生境,或寒冷地区物种通过长距离扩散进入正在生长的高原[26 -27 ] ,导致该区域高寒生物多样性增加.高寒区生物多样性快速增长的时间通常同步于或略晚于高山或高原形成的时间[22 ,25 ,27 ] ,因此,高寒区生物类群的多样性演化过程和分化时间对我们认知构造活动期次[28 -29 ] ,推断高原隆升历史及其气候效应[1 ,30 -31 ] 等具有启示意义,甚至能够揭示以前所没有注意到的地质事件[22 ] .近年来,青藏高寒区生物地理学研究取得长足发展,为我们提供了一个独特的视角来探讨区域生物多样性演化、地质活动和气候变化之间的关系[25 ] . ...
... ,25 ,27 ],因此,高寒区生物类群的多样性演化过程和分化时间对我们认知构造活动期次[28 -29 ] ,推断高原隆升历史及其气候效应[1 ,30 -31 ] 等具有启示意义,甚至能够揭示以前所没有注意到的地质事件[22 ] .近年来,青藏高寒区生物地理学研究取得长足发展,为我们提供了一个独特的视角来探讨区域生物多样性演化、地质活动和气候变化之间的关系[25 ] . ...
... [25 ]. ...
... 最近,青藏高原古地貌重建工作取得了丰硕的成果(图3 ),但是各种指标重建结果往往存在矛盾(图4 )[25 ,79 -83 ] .同位素重建结果表明高原南部在古近纪早期就存在安第斯型的巨大山系[84 -85 ] ;古生物研究发现高原腹地从中始新世到晚渐新世都存在东西向的大峡谷,该峡谷底部海拔在2 000 m左右,两侧为高大的冈底斯山脉和羌塘山脉[82 ,86 -88 ] ,动植物如巨犀(Paraceratherium )[89 ] 、攀鲈(Eoanabas )[90 -91 ] 、臭椿(Ailanthus )[88 ] 、椿榆(Cedrelospermum )[92 ] 、兔耳果(Lagokarpos )[93 ] 等在古近纪都可以通过青藏地区峡谷与周边动植物区系交流,新近纪之前的青藏地区虽然地形复杂,但是尚未隆升成今天的高原;横断山区某些区域则在晚始新世-早渐新世之交就已经获得现今的海拔高度[94 -95 ] ;高原北部和喜马拉雅山脉则在新近纪形成现在的地貌格局[25 ,96 -97 ] .古生物研究也发现青藏高原在晚上新世就已经存在高寒生境和冰缘环境,成为第四纪冰期动物忍耐寒冷环境的“训练营”,第四纪冰期来临之后这些适应寒冷环境的生物才扩散至北半球高纬度地区[34 ,98 -99 ] .因此,高原及周边地区高寒生物类群的起源时间表明在渐新世青藏高原部分区域就已经出现高寒生态系统,晚中新世以来青藏高原地貌与气候继续发生剧烈变化,促使这些物种的祖先类群迁入或就地演化积累适应高寒环境的基因和形态,并最终演化成适应高寒生境的新物种[25 ] .高寒区物种在晚上新世的加速分化可能与高原巨大山系和水系、冰川活动等造成的地理隔离有关[26 ] .随着第四纪冰期的到来,青藏高原及周边地区的高寒生物区也开始与北半球高纬度地区存在密切联系[34 ,98 -99 ] .同时,演化生物学研究也发现在第四纪冰期期间并未形成覆盖整个青藏高原的大冰盖,高原面上仍存在很多生物避难所[26 ] . ...
... [25 ,96 -97 ].古生物研究也发现青藏高原在晚上新世就已经存在高寒生境和冰缘环境,成为第四纪冰期动物忍耐寒冷环境的“训练营”,第四纪冰期来临之后这些适应寒冷环境的生物才扩散至北半球高纬度地区[34 ,98 -99 ] .因此,高原及周边地区高寒生物类群的起源时间表明在渐新世青藏高原部分区域就已经出现高寒生态系统,晚中新世以来青藏高原地貌与气候继续发生剧烈变化,促使这些物种的祖先类群迁入或就地演化积累适应高寒环境的基因和形态,并最终演化成适应高寒生境的新物种[25 ] .高寒区物种在晚上新世的加速分化可能与高原巨大山系和水系、冰川活动等造成的地理隔离有关[26 ] .随着第四纪冰期的到来,青藏高原及周边地区的高寒生物区也开始与北半球高纬度地区存在密切联系[34 ,98 -99 ] .同时,演化生物学研究也发现在第四纪冰期期间并未形成覆盖整个青藏高原的大冰盖,高原面上仍存在很多生物避难所[26 ] . ...
... [25 ].高寒区物种在晚上新世的加速分化可能与高原巨大山系和水系、冰川活动等造成的地理隔离有关[26 ] .随着第四纪冰期的到来,青藏高原及周边地区的高寒生物区也开始与北半球高纬度地区存在密切联系[34 ,98 -99 ] .同时,演化生物学研究也发现在第四纪冰期期间并未形成覆盖整个青藏高原的大冰盖,高原面上仍存在很多生物避难所[26 ] . ...
... 青藏高原新生代古高程定量重建研究点(根据文献[
25 ]修改)
Quantitative reconstruction paleoelevation sites on Tibetan Plateau (Revised from Reference [<xref ref-type="bibr" rid="R25">25</xref>]) Fig.3 ![]()
图4 青藏高原南北剖面古高程变化示意图(根据文献[<xref ref-type="bibr" rid="R25">25</xref>]、[<xref ref-type="bibr" rid="R34">34</xref>]、[<xref ref-type="bibr" rid="R82">82</xref>]、[<xref ref-type="bibr" rid="R87">87</xref>]、[<xref ref-type="bibr" rid="R100">100</xref>]修改) North-south paleoelevation profiles on the Tibetan Plateau (Revised from References [25],[34],[82],[87],[100]) Fig.4 ![]()
随着DNA测序技术和比较系统学方法的发展,大量涌现的青藏高寒区生物地理学研究为我们理解高原隆升过程及其气候效应提供了生物演化方面的证据[25 ] .但是,演化生物学家通常仅对单一特定类群的系统发育过程开展研究,很少进行系统的总结工作[25 ] ,对某一类群遗传数据的解释也往往与最新的青藏高原隆升和气候变化研究进展脱节,不能很好地将最新的关于高原隆升和气候变化的研究与生物类群演化联系在一起[101 ] ,同时也没有认识到青藏高原的隆升过程是阶段性的,没有对青藏高原的不同组成部分进行区分[26 ,53 ,81 ] .地质学研究大多认为高原地貌格局形成于中新世之前,但是综合近年来的生物地理学研究结果可以发现,青藏高寒生物类群的分化时间多集中在晚中新世,种内分化加速和多样性增加主要发生在晚上新世到第四纪期间(图5 ),这些不同类群在分化时间上的巧合可能反映青藏高寒区生物演化受到晚中新世以来构造和气候事件的影响,但是并未引起地质学家足够的重视[25 ] . ...
... Quantitative reconstruction paleoelevation sites on Tibetan Plateau (Revised from Reference [
25 ])
Fig.3 ![]()
图4 青藏高原南北剖面古高程变化示意图(根据文献[<xref ref-type="bibr" rid="R25">25</xref>]、[<xref ref-type="bibr" rid="R34">34</xref>]、[<xref ref-type="bibr" rid="R82">82</xref>]、[<xref ref-type="bibr" rid="R87">87</xref>]、[<xref ref-type="bibr" rid="R100">100</xref>]修改) North-south paleoelevation profiles on the Tibetan Plateau (Revised from References [25],[34],[82],[87],[100]) Fig.4 ![]()
随着DNA测序技术和比较系统学方法的发展,大量涌现的青藏高寒区生物地理学研究为我们理解高原隆升过程及其气候效应提供了生物演化方面的证据[25 ] .但是,演化生物学家通常仅对单一特定类群的系统发育过程开展研究,很少进行系统的总结工作[25 ] ,对某一类群遗传数据的解释也往往与最新的青藏高原隆升和气候变化研究进展脱节,不能很好地将最新的关于高原隆升和气候变化的研究与生物类群演化联系在一起[101 ] ,同时也没有认识到青藏高原的隆升过程是阶段性的,没有对青藏高原的不同组成部分进行区分[26 ,53 ,81 ] .地质学研究大多认为高原地貌格局形成于中新世之前,但是综合近年来的生物地理学研究结果可以发现,青藏高寒生物类群的分化时间多集中在晚中新世,种内分化加速和多样性增加主要发生在晚上新世到第四纪期间(图5 ),这些不同类群在分化时间上的巧合可能反映青藏高寒区生物演化受到晚中新世以来构造和气候事件的影响,但是并未引起地质学家足够的重视[25 ] . ...
... 青藏高原南北剖面古高程变化示意图(根据文献[
25 ]、[
34 ]、[
82 ]、[
87 ]、[
100 ]修改)
North-south paleoelevation profiles on the Tibetan Plateau (Revised from References [25],[34],[82],[87],[100]) Fig.4 ![]()
随着DNA测序技术和比较系统学方法的发展,大量涌现的青藏高寒区生物地理学研究为我们理解高原隆升过程及其气候效应提供了生物演化方面的证据[25 ] .但是,演化生物学家通常仅对单一特定类群的系统发育过程开展研究,很少进行系统的总结工作[25 ] ,对某一类群遗传数据的解释也往往与最新的青藏高原隆升和气候变化研究进展脱节,不能很好地将最新的关于高原隆升和气候变化的研究与生物类群演化联系在一起[101 ] ,同时也没有认识到青藏高原的隆升过程是阶段性的,没有对青藏高原的不同组成部分进行区分[26 ,53 ,81 ] .地质学研究大多认为高原地貌格局形成于中新世之前,但是综合近年来的生物地理学研究结果可以发现,青藏高寒生物类群的分化时间多集中在晚中新世,种内分化加速和多样性增加主要发生在晚上新世到第四纪期间(图5 ),这些不同类群在分化时间上的巧合可能反映青藏高寒区生物演化受到晚中新世以来构造和气候事件的影响,但是并未引起地质学家足够的重视[25 ] . ...
... 随着DNA测序技术和比较系统学方法的发展,大量涌现的青藏高寒区生物地理学研究为我们理解高原隆升过程及其气候效应提供了生物演化方面的证据[25 ] .但是,演化生物学家通常仅对单一特定类群的系统发育过程开展研究,很少进行系统的总结工作[25 ] ,对某一类群遗传数据的解释也往往与最新的青藏高原隆升和气候变化研究进展脱节,不能很好地将最新的关于高原隆升和气候变化的研究与生物类群演化联系在一起[101 ] ,同时也没有认识到青藏高原的隆升过程是阶段性的,没有对青藏高原的不同组成部分进行区分[26 ,53 ,81 ] .地质学研究大多认为高原地貌格局形成于中新世之前,但是综合近年来的生物地理学研究结果可以发现,青藏高寒生物类群的分化时间多集中在晚中新世,种内分化加速和多样性增加主要发生在晚上新世到第四纪期间(图5 ),这些不同类群在分化时间上的巧合可能反映青藏高寒区生物演化受到晚中新世以来构造和气候事件的影响,但是并未引起地质学家足够的重视[25 ] . ...
... [25 ],对某一类群遗传数据的解释也往往与最新的青藏高原隆升和气候变化研究进展脱节,不能很好地将最新的关于高原隆升和气候变化的研究与生物类群演化联系在一起[101 ] ,同时也没有认识到青藏高原的隆升过程是阶段性的,没有对青藏高原的不同组成部分进行区分[26 ,53 ,81 ] .地质学研究大多认为高原地貌格局形成于中新世之前,但是综合近年来的生物地理学研究结果可以发现,青藏高寒生物类群的分化时间多集中在晚中新世,种内分化加速和多样性增加主要发生在晚上新世到第四纪期间(图5 ),这些不同类群在分化时间上的巧合可能反映青藏高寒区生物演化受到晚中新世以来构造和气候事件的影响,但是并未引起地质学家足够的重视[25 ] . ...
... [25 ]. ...
Ancient orogenic and monsoon-driven assembly of the world’s richest temperate alpine flora
15
2020
... 青藏高原及周边地区物种丰富,是全球生物多样性热点地区之一(图1 )[10 -11 ] .超过1 500属和12 000种植物生活在青藏高原地区,其中3 673种是该区域的特有种子植物[12 -14 ] .该地区植物区系独特,是全球海拔最高、高寒生物多样性最丰富的地区,被称为植物区系的“第三极”[15 ] .此外,青藏高原地区还生活着148种特有哺乳动物[16 ] ,167种特有鸟类[17 ] 以及六足动物亚门的许多特有种类[18 -19 ] .青藏高原平均海拔超过4 500 m,环境极端恶劣,低温、严寒、干旱、低氧、高紫外线辐射等是制约高寒区生物生存的关键因素,同时也是促使该区域生物特有种分化形成的主要原因[20 -21 ] .地质时期构造活动和气候变化可以形成地理隔离障碍促使物种分化和多样性增加,也可以通过减少生物扩散障碍使得物种分布区扩大,不同区域间的生物交流增加[22 -25 ] .青藏高原高寒区物种的形成和演化过程受到青藏高原构造抬升及其相应气候变化的影响[25 -26 ] .伴随着高原海拔的增长,物种就地演化或低海拔物种向高海拔迁移并适应高寒生境,或寒冷地区物种通过长距离扩散进入正在生长的高原[26 -27 ] ,导致该区域高寒生物多样性增加.高寒区生物多样性快速增长的时间通常同步于或略晚于高山或高原形成的时间[22 ,25 ,27 ] ,因此,高寒区生物类群的多样性演化过程和分化时间对我们认知构造活动期次[28 -29 ] ,推断高原隆升历史及其气候效应[1 ,30 -31 ] 等具有启示意义,甚至能够揭示以前所没有注意到的地质事件[22 ] .近年来,青藏高寒区生物地理学研究取得长足发展,为我们提供了一个独特的视角来探讨区域生物多样性演化、地质活动和气候变化之间的关系[25 ] . ...
... [26 -27 ],导致该区域高寒生物多样性增加.高寒区生物多样性快速增长的时间通常同步于或略晚于高山或高原形成的时间[22 ,25 ,27 ] ,因此,高寒区生物类群的多样性演化过程和分化时间对我们认知构造活动期次[28 -29 ] ,推断高原隆升历史及其气候效应[1 ,30 -31 ] 等具有启示意义,甚至能够揭示以前所没有注意到的地质事件[22 ] .近年来,青藏高寒区生物地理学研究取得长足发展,为我们提供了一个独特的视角来探讨区域生物多样性演化、地质活动和气候变化之间的关系[25 ] . ...
... 高寒生物区是分布于树线和雪线之间的独特生态系统,遍布全球各大洲高山和高原之上[26 ,32 ] .高寒生物区自然环境极端恶劣,物种为适应环境进化出独特的抗寒、抗旱、耐低温、抗紫外线辐射、抗缺氧等性状,并在这些极端环境中经过悠久的演化历史最终存活下来[21 ,26 ] ,是获取抗性基因的重要宝库.高寒区生物是适应极端环境的产物,对环境变化非常敏感.近年来全球变暖加剧了高寒区生物灭绝风险,使得该区域成为全球生物多样性保护研究中重点关注的区域.同时,高寒区生物是山脉或高原隆升到一定海拔后的产物[33 ] ,研究高寒区生物多样性的起源和演化过程,不仅可以认识高寒区生物多样性的形成过程,还可以预测未来气候变化对高寒区生物多样性的影响,同时也能够推断高山、高原地貌和气候的演化历史[20 ,23 -24 ,34 ] . ...
... ,26 ],是获取抗性基因的重要宝库.高寒区生物是适应极端环境的产物,对环境变化非常敏感.近年来全球变暖加剧了高寒区生物灭绝风险,使得该区域成为全球生物多样性保护研究中重点关注的区域.同时,高寒区生物是山脉或高原隆升到一定海拔后的产物[33 ] ,研究高寒区生物多样性的起源和演化过程,不仅可以认识高寒区生物多样性的形成过程,还可以预测未来气候变化对高寒区生物多样性的影响,同时也能够推断高山、高原地貌和气候的演化历史[20 ,23 -24 ,34 ] . ...
... 生物地理学主要研究生物的时空分布格局以及性状演化过程及其形成原因和机制.从1852年开始,生物地理学家就开始寻找和总结动植物类群的分布模式,推断其历史分布区演化,并试图将这些分布模式与影响其演化的地质过程联系起来[35 ] .随着DNA测序技术的发展,研究者对标本进行DNA测序,基于宽松分子钟模型或使用年代和鉴定准确的化石记录来校正分子钟,使用最大似然法和贝叶斯法构建系统树,估算物种演化的分支时间和演化历史[18 ] ,通过现生类群的分布格局来推断祖先类群的地理分布.扩散-灭绝-进化分支(dispersal-extinction-cladogenesis)模型是目前最常使用的模型,使用最大似然法和贝叶斯法推断物种的扩散和灭绝历史,用隔离分化或支序发生来解释不同区域间类群分布格局[26 ,36 ] .同时,利用比较系统发育学方法还可以估算物种多样化速率,推断物种分布格局和多样性演化历史,探讨构造事件和气候变化对多样性形成和演化过程的影响[26 ] . ...
... [26 ]. ...
... 青藏高原平均海拔超过4 500 m,是全球中低纬冰川分布最为集中的区域[37 ] ,被誉为世界“第三极”[38 -39 ] ,高寒生物区分布广泛,对气候变化反映敏感[40 ] .近年来,全球变暖加剧,青藏高原及周边地区升温尤其明显[41 -42 ] ,高寒区生物生存压力加大.对青藏高寒区生物开展生物地理学研究有助于探讨青藏高寒区生物多样性起源演化,并预测未来多样性演化对全球变化的响应.此外,最近的研究表明青藏高原东南缘高寒植物多样性在早渐新世就开始积累,是现存高寒生物区中起源最早的[26 ] ,远比安第斯山脉(约3.5 Ma)[43 ] 、阿尔卑斯山脉(<1.75 Ma)[44 ] 、新西兰南阿尔卑斯山脉(<5 Ma)[45 ] 、东非高山(8~7 Ma)[46 ] 高寒植物群起源地早.青藏高寒区生物多样性演化、物种扩散、分化与灭绝受到高原阶段性隆升及其气候效应的影响,对其开展研究能够进一步理解高原隆升过程及其如何对生物多样性演化产生影响. ...
... 最近,青藏高原古地貌重建工作取得了丰硕的成果(图3 ),但是各种指标重建结果往往存在矛盾(图4 )[25 ,79 -83 ] .同位素重建结果表明高原南部在古近纪早期就存在安第斯型的巨大山系[84 -85 ] ;古生物研究发现高原腹地从中始新世到晚渐新世都存在东西向的大峡谷,该峡谷底部海拔在2 000 m左右,两侧为高大的冈底斯山脉和羌塘山脉[82 ,86 -88 ] ,动植物如巨犀(Paraceratherium )[89 ] 、攀鲈(Eoanabas )[90 -91 ] 、臭椿(Ailanthus )[88 ] 、椿榆(Cedrelospermum )[92 ] 、兔耳果(Lagokarpos )[93 ] 等在古近纪都可以通过青藏地区峡谷与周边动植物区系交流,新近纪之前的青藏地区虽然地形复杂,但是尚未隆升成今天的高原;横断山区某些区域则在晚始新世-早渐新世之交就已经获得现今的海拔高度[94 -95 ] ;高原北部和喜马拉雅山脉则在新近纪形成现在的地貌格局[25 ,96 -97 ] .古生物研究也发现青藏高原在晚上新世就已经存在高寒生境和冰缘环境,成为第四纪冰期动物忍耐寒冷环境的“训练营”,第四纪冰期来临之后这些适应寒冷环境的生物才扩散至北半球高纬度地区[34 ,98 -99 ] .因此,高原及周边地区高寒生物类群的起源时间表明在渐新世青藏高原部分区域就已经出现高寒生态系统,晚中新世以来青藏高原地貌与气候继续发生剧烈变化,促使这些物种的祖先类群迁入或就地演化积累适应高寒环境的基因和形态,并最终演化成适应高寒生境的新物种[25 ] .高寒区物种在晚上新世的加速分化可能与高原巨大山系和水系、冰川活动等造成的地理隔离有关[26 ] .随着第四纪冰期的到来,青藏高原及周边地区的高寒生物区也开始与北半球高纬度地区存在密切联系[34 ,98 -99 ] .同时,演化生物学研究也发现在第四纪冰期期间并未形成覆盖整个青藏高原的大冰盖,高原面上仍存在很多生物避难所[26 ] . ...
... [26 ]. ...
... 随着DNA测序技术和比较系统学方法的发展,大量涌现的青藏高寒区生物地理学研究为我们理解高原隆升过程及其气候效应提供了生物演化方面的证据[25 ] .但是,演化生物学家通常仅对单一特定类群的系统发育过程开展研究,很少进行系统的总结工作[25 ] ,对某一类群遗传数据的解释也往往与最新的青藏高原隆升和气候变化研究进展脱节,不能很好地将最新的关于高原隆升和气候变化的研究与生物类群演化联系在一起[101 ] ,同时也没有认识到青藏高原的隆升过程是阶段性的,没有对青藏高原的不同组成部分进行区分[26 ,53 ,81 ] .地质学研究大多认为高原地貌格局形成于中新世之前,但是综合近年来的生物地理学研究结果可以发现,青藏高寒生物类群的分化时间多集中在晚中新世,种内分化加速和多样性增加主要发生在晚上新世到第四纪期间(图5 ),这些不同类群在分化时间上的巧合可能反映青藏高寒区生物演化受到晚中新世以来构造和气候事件的影响,但是并未引起地质学家足够的重视[25 ] . ...
... 近期,演化生物学家与地质学家对青藏高原及周边高寒生物区18个被子植物类群共3 798个植物物种的遗传数据进行整合研究,发现横断山区是世界上起源最早的高寒生物区,该区域高寒植物多样性在早渐新世就已经开始积累,并分别在晚渐新世和中中新世就地演化速率快速增加(图4 )[26 ] .高寒植物在早渐新世就出现在横断山区与该区域在晚始新世-早渐新世之交就已经出现与现今海拔相似的高山环境有关[94 -95 ] .中新世就地演化速率快速增加与季风气候增强,降水增加,河流切割加剧形成生物地理隔离有关[26 ,104 ] .横断山区高寒生物演化进程与喜马拉雅山脉和青藏高原腹地的对比研究发现,横断山区是青藏高原高寒生物区物种起源和分化的摇篮,也是高寒生物的重要避难所,为青藏高原腹地和喜马拉雅山脉高寒生物多样性提供主要来源[26 ] .第四纪以来,冰期-间冰期气候变化剧烈,横断山区近南北向的高山狭谷地貌格局为遭受极端气候的北方类群和高寒生物类群的迁徙提供廊道,此外,深切狭谷与高山也可以保障山地动植物类群短时间、短距离地垂直迁徙;与此同时,横断山区高山狭谷地貌导致的地理和生境的巨大差异,提供了物种隔离障碍,成为许多动植物类群的新种分化地[26 ] .冰期时,全球变冷,冰川活动加剧,高寒生物区面积扩张,临近山地的高寒生物区之间的交流也大大增加[26 ] .该研究为高原隆升、气候变化和生物演化提供一个多学科交叉研究的范例. ...
... [26 ,104 ].横断山区高寒生物演化进程与喜马拉雅山脉和青藏高原腹地的对比研究发现,横断山区是青藏高原高寒生物区物种起源和分化的摇篮,也是高寒生物的重要避难所,为青藏高原腹地和喜马拉雅山脉高寒生物多样性提供主要来源[26 ] .第四纪以来,冰期-间冰期气候变化剧烈,横断山区近南北向的高山狭谷地貌格局为遭受极端气候的北方类群和高寒生物类群的迁徙提供廊道,此外,深切狭谷与高山也可以保障山地动植物类群短时间、短距离地垂直迁徙;与此同时,横断山区高山狭谷地貌导致的地理和生境的巨大差异,提供了物种隔离障碍,成为许多动植物类群的新种分化地[26 ] .冰期时,全球变冷,冰川活动加剧,高寒生物区面积扩张,临近山地的高寒生物区之间的交流也大大增加[26 ] .该研究为高原隆升、气候变化和生物演化提供一个多学科交叉研究的范例. ...
... [26 ].第四纪以来,冰期-间冰期气候变化剧烈,横断山区近南北向的高山狭谷地貌格局为遭受极端气候的北方类群和高寒生物类群的迁徙提供廊道,此外,深切狭谷与高山也可以保障山地动植物类群短时间、短距离地垂直迁徙;与此同时,横断山区高山狭谷地貌导致的地理和生境的巨大差异,提供了物种隔离障碍,成为许多动植物类群的新种分化地[26 ] .冰期时,全球变冷,冰川活动加剧,高寒生物区面积扩张,临近山地的高寒生物区之间的交流也大大增加[26 ] .该研究为高原隆升、气候变化和生物演化提供一个多学科交叉研究的范例. ...
... [26 ].冰期时,全球变冷,冰川活动加剧,高寒生物区面积扩张,临近山地的高寒生物区之间的交流也大大增加[26 ] .该研究为高原隆升、气候变化和生物演化提供一个多学科交叉研究的范例. ...
... [26 ].该研究为高原隆升、气候变化和生物演化提供一个多学科交叉研究的范例. ...
Evolution of endemism on a young tropical mountain
2
2015
... 青藏高原及周边地区物种丰富,是全球生物多样性热点地区之一(图1 )[10 -11 ] .超过1 500属和12 000种植物生活在青藏高原地区,其中3 673种是该区域的特有种子植物[12 -14 ] .该地区植物区系独特,是全球海拔最高、高寒生物多样性最丰富的地区,被称为植物区系的“第三极”[15 ] .此外,青藏高原地区还生活着148种特有哺乳动物[16 ] ,167种特有鸟类[17 ] 以及六足动物亚门的许多特有种类[18 -19 ] .青藏高原平均海拔超过4 500 m,环境极端恶劣,低温、严寒、干旱、低氧、高紫外线辐射等是制约高寒区生物生存的关键因素,同时也是促使该区域生物特有种分化形成的主要原因[20 -21 ] .地质时期构造活动和气候变化可以形成地理隔离障碍促使物种分化和多样性增加,也可以通过减少生物扩散障碍使得物种分布区扩大,不同区域间的生物交流增加[22 -25 ] .青藏高原高寒区物种的形成和演化过程受到青藏高原构造抬升及其相应气候变化的影响[25 -26 ] .伴随着高原海拔的增长,物种就地演化或低海拔物种向高海拔迁移并适应高寒生境,或寒冷地区物种通过长距离扩散进入正在生长的高原[26 -27 ] ,导致该区域高寒生物多样性增加.高寒区生物多样性快速增长的时间通常同步于或略晚于高山或高原形成的时间[22 ,25 ,27 ] ,因此,高寒区生物类群的多样性演化过程和分化时间对我们认知构造活动期次[28 -29 ] ,推断高原隆升历史及其气候效应[1 ,30 -31 ] 等具有启示意义,甚至能够揭示以前所没有注意到的地质事件[22 ] .近年来,青藏高寒区生物地理学研究取得长足发展,为我们提供了一个独特的视角来探讨区域生物多样性演化、地质活动和气候变化之间的关系[25 ] . ...
... ,27 ],因此,高寒区生物类群的多样性演化过程和分化时间对我们认知构造活动期次[28 -29 ] ,推断高原隆升历史及其气候效应[1 ,30 -31 ] 等具有启示意义,甚至能够揭示以前所没有注意到的地质事件[22 ] .近年来,青藏高寒区生物地理学研究取得长足发展,为我们提供了一个独特的视角来探讨区域生物多样性演化、地质活动和气候变化之间的关系[25 ] . ...
Evolutionary history and underlying adaptation of alpine plants on the Qinghai-Tibet Plateau
1
2014
... 青藏高原及周边地区物种丰富,是全球生物多样性热点地区之一(图1 )[10 -11 ] .超过1 500属和12 000种植物生活在青藏高原地区,其中3 673种是该区域的特有种子植物[12 -14 ] .该地区植物区系独特,是全球海拔最高、高寒生物多样性最丰富的地区,被称为植物区系的“第三极”[15 ] .此外,青藏高原地区还生活着148种特有哺乳动物[16 ] ,167种特有鸟类[17 ] 以及六足动物亚门的许多特有种类[18 -19 ] .青藏高原平均海拔超过4 500 m,环境极端恶劣,低温、严寒、干旱、低氧、高紫外线辐射等是制约高寒区生物生存的关键因素,同时也是促使该区域生物特有种分化形成的主要原因[20 -21 ] .地质时期构造活动和气候变化可以形成地理隔离障碍促使物种分化和多样性增加,也可以通过减少生物扩散障碍使得物种分布区扩大,不同区域间的生物交流增加[22 -25 ] .青藏高原高寒区物种的形成和演化过程受到青藏高原构造抬升及其相应气候变化的影响[25 -26 ] .伴随着高原海拔的增长,物种就地演化或低海拔物种向高海拔迁移并适应高寒生境,或寒冷地区物种通过长距离扩散进入正在生长的高原[26 -27 ] ,导致该区域高寒生物多样性增加.高寒区生物多样性快速增长的时间通常同步于或略晚于高山或高原形成的时间[22 ,25 ,27 ] ,因此,高寒区生物类群的多样性演化过程和分化时间对我们认知构造活动期次[28 -29 ] ,推断高原隆升历史及其气候效应[1 ,30 -31 ] 等具有启示意义,甚至能够揭示以前所没有注意到的地质事件[22 ] .近年来,青藏高寒区生物地理学研究取得长足发展,为我们提供了一个独特的视角来探讨区域生物多样性演化、地质活动和气候变化之间的关系[25 ] . ...
Origin, evolution, and systematics of Himalaya endemic genera
1
2007
... 青藏高原及周边地区物种丰富,是全球生物多样性热点地区之一(图1 )[10 -11 ] .超过1 500属和12 000种植物生活在青藏高原地区,其中3 673种是该区域的特有种子植物[12 -14 ] .该地区植物区系独特,是全球海拔最高、高寒生物多样性最丰富的地区,被称为植物区系的“第三极”[15 ] .此外,青藏高原地区还生活着148种特有哺乳动物[16 ] ,167种特有鸟类[17 ] 以及六足动物亚门的许多特有种类[18 -19 ] .青藏高原平均海拔超过4 500 m,环境极端恶劣,低温、严寒、干旱、低氧、高紫外线辐射等是制约高寒区生物生存的关键因素,同时也是促使该区域生物特有种分化形成的主要原因[20 -21 ] .地质时期构造活动和气候变化可以形成地理隔离障碍促使物种分化和多样性增加,也可以通过减少生物扩散障碍使得物种分布区扩大,不同区域间的生物交流增加[22 -25 ] .青藏高原高寒区物种的形成和演化过程受到青藏高原构造抬升及其相应气候变化的影响[25 -26 ] .伴随着高原海拔的增长,物种就地演化或低海拔物种向高海拔迁移并适应高寒生境,或寒冷地区物种通过长距离扩散进入正在生长的高原[26 -27 ] ,导致该区域高寒生物多样性增加.高寒区生物多样性快速增长的时间通常同步于或略晚于高山或高原形成的时间[22 ,25 ,27 ] ,因此,高寒区生物类群的多样性演化过程和分化时间对我们认知构造活动期次[28 -29 ] ,推断高原隆升历史及其气候效应[1 ,30 -31 ] 等具有启示意义,甚至能够揭示以前所没有注意到的地质事件[22 ] .近年来,青藏高寒区生物地理学研究取得长足发展,为我们提供了一个独特的视角来探讨区域生物多样性演化、地质活动和气候变化之间的关系[25 ] . ...
Partitioned Bayesian analyses, dispersal-vicariance analysis, and the biogeography of Chinese toad-headed lizards (Agamidae: Phrynocephalus ): A re-evaluation
1
2007
... 青藏高原及周边地区物种丰富,是全球生物多样性热点地区之一(图1 )[10 -11 ] .超过1 500属和12 000种植物生活在青藏高原地区,其中3 673种是该区域的特有种子植物[12 -14 ] .该地区植物区系独特,是全球海拔最高、高寒生物多样性最丰富的地区,被称为植物区系的“第三极”[15 ] .此外,青藏高原地区还生活着148种特有哺乳动物[16 ] ,167种特有鸟类[17 ] 以及六足动物亚门的许多特有种类[18 -19 ] .青藏高原平均海拔超过4 500 m,环境极端恶劣,低温、严寒、干旱、低氧、高紫外线辐射等是制约高寒区生物生存的关键因素,同时也是促使该区域生物特有种分化形成的主要原因[20 -21 ] .地质时期构造活动和气候变化可以形成地理隔离障碍促使物种分化和多样性增加,也可以通过减少生物扩散障碍使得物种分布区扩大,不同区域间的生物交流增加[22 -25 ] .青藏高原高寒区物种的形成和演化过程受到青藏高原构造抬升及其相应气候变化的影响[25 -26 ] .伴随着高原海拔的增长,物种就地演化或低海拔物种向高海拔迁移并适应高寒生境,或寒冷地区物种通过长距离扩散进入正在生长的高原[26 -27 ] ,导致该区域高寒生物多样性增加.高寒区生物多样性快速增长的时间通常同步于或略晚于高山或高原形成的时间[22 ,25 ,27 ] ,因此,高寒区生物类群的多样性演化过程和分化时间对我们认知构造活动期次[28 -29 ] ,推断高原隆升历史及其气候效应[1 ,30 -31 ] 等具有启示意义,甚至能够揭示以前所没有注意到的地质事件[22 ] .近年来,青藏高寒区生物地理学研究取得长足发展,为我们提供了一个独特的视角来探讨区域生物多样性演化、地质活动和气候变化之间的关系[25 ] . ...
Uplift of the Tibetan Plateau: evidence from divergence times of glyptosternoid catfishes
3
2006
... 青藏高原及周边地区物种丰富,是全球生物多样性热点地区之一(图1 )[10 -11 ] .超过1 500属和12 000种植物生活在青藏高原地区,其中3 673种是该区域的特有种子植物[12 -14 ] .该地区植物区系独特,是全球海拔最高、高寒生物多样性最丰富的地区,被称为植物区系的“第三极”[15 ] .此外,青藏高原地区还生活着148种特有哺乳动物[16 ] ,167种特有鸟类[17 ] 以及六足动物亚门的许多特有种类[18 -19 ] .青藏高原平均海拔超过4 500 m,环境极端恶劣,低温、严寒、干旱、低氧、高紫外线辐射等是制约高寒区生物生存的关键因素,同时也是促使该区域生物特有种分化形成的主要原因[20 -21 ] .地质时期构造活动和气候变化可以形成地理隔离障碍促使物种分化和多样性增加,也可以通过减少生物扩散障碍使得物种分布区扩大,不同区域间的生物交流增加[22 -25 ] .青藏高原高寒区物种的形成和演化过程受到青藏高原构造抬升及其相应气候变化的影响[25 -26 ] .伴随着高原海拔的增长,物种就地演化或低海拔物种向高海拔迁移并适应高寒生境,或寒冷地区物种通过长距离扩散进入正在生长的高原[26 -27 ] ,导致该区域高寒生物多样性增加.高寒区生物多样性快速增长的时间通常同步于或略晚于高山或高原形成的时间[22 ,25 ,27 ] ,因此,高寒区生物类群的多样性演化过程和分化时间对我们认知构造活动期次[28 -29 ] ,推断高原隆升历史及其气候效应[1 ,30 -31 ] 等具有启示意义,甚至能够揭示以前所没有注意到的地质事件[22 ] .近年来,青藏高寒区生物地理学研究取得长足发展,为我们提供了一个独特的视角来探讨区域生物多样性演化、地质活动和气候变化之间的关系[25 ] . ...
... 鲤形目裂腹鱼亚科(Schizothoracinae)鱼类是青藏高原及其周边地区的特有鱼类[74 ] ,地貌和水系格局的改变对裂腹鱼亚科的系统演化和分布产生深刻影响,而地质活动和气候事件制约着地貌和水系格局的形成和演化,因此可以通过裂腹鱼亚科系统演化、历史地理分布和化石解剖形态来推断地质活动和气候事件发生的时间,为青藏高原的隆升研究提供证据[74 -75 ] .裂腹鱼亚科系统发育研究表明在晚中新世晚期或上新世裂腹鱼亚科鱼类就已经出现在青藏高原及周边水系中,在晚更新世期间种内分化加速,不同海拔之间的类群产生等级分化,种间多样性增加,第四纪以来青藏高原隆升导致的水系改组是推动裂腹鱼亚科鱼类演化的原动力[31 ,76 -78 ] . ...
... Biological evolution in the alpine region of Tibetan Plateau
Table 1 种属 分布区域 适应高海拔类群分化时间或多样性增加时间 文献来源 Sinopodophyllum hexandrum 青藏高原东部 在约(6.52±1.89) Ma期间出现在青藏高原东部,但是直到3~2 Ma以来才演化出适应高山环境的形态 [47 ] Nannoglottis 青藏高原 在23~32 Ma期间出现在青藏高原,随后多样性增高,其高山灌丛和针叶林生活类型分支时间约为3.4 Ma [48 ] Reaumuria soongarica 西北干旱区、青藏高原北部 高原内部分布类群演化时间2.0~6.3 Ma,高原东北缘与高原西北类群分化时间1.1~3.5 Ma [49 ] Orchidinae 青藏高原及喜马拉雅山脉 青藏高原特有种大多是在过去6 Ma里进化而来 [50 ] Oxyria sinensis 青藏高原东南部 高山类群出现时间7.54~24.36 Ma,种内多样化增加时间1.74~0.86 Ma [51 ] Juniperus microsperma 西藏波密 早中新世21.6~14 Ma期间与其他种群分化 [52 ] Rhododendron 北半球 横断山区概属多样性在晚中新世(约8 Ma)快速增加 [53 ] Roscoea, Cautleya 喜马拉雅-横断山区 在早渐新世(~32 Ma)两属出现,Roscoea在横断山区与喜马拉雅山脉的两个分支大概在渐新世-中新世之交分化 [54 ] Nanorana parkeri 青藏高原南部 高原东部和西部类群分化时间1.4~3.7 Ma [55 ] Paini 南亚、东南亚、青藏高原 在渐新世早期棘蛙族高原-喜马拉雅类群与中国第二阶梯上生活的类群分化,随后在在26.1~12.5 Ma高原生活类群与喜马拉雅山脉生活类群分化,高原内部生活类群在晚中新世到上新世期间分化速率加快 [22 ] Bufo tibetanus 青藏高原、四川、甘肃、陕西等 高原种类形成于上新世期间 [56 ] Phrynocephalus 亚洲干旱区 在7.2~13.0 Ma期间演化出胎生类群,3.2~7.3 Ma期间种间多样性增加 [57 ] Phrynocephalus erythrurus 羌塘盆地 在3.67 Ma左右北部类群与南部类群发生分化,约2.76 Ma北部类群内部进一步分化 [58 ] Gnaptorina 青藏高原及周边高海拔地区 始新世起源于青藏高原东南缘 [18 -19 ] Coelotinae 北半球温带、亚热带 晚始新世高原东南缘隆升阻碍南方隙蛛向北扩散,渐新世高原中部和北部隆起导致北方隙蛛早期多样化进程中的灭绝事件 [59 ] Montifringilla, Onychostruthus, Pyrgilauda 青藏高原 雪雀的共同祖先在约14 Ma就应经生活在青藏高原,并进化出适应高海拔极端环境的性状 [21 ] Parus major 亚洲、欧洲 蒙古分布类群与其他种类分化时间为2.8~0.7 Ma,喜马拉雅山脉东部与中国中东部类群分化时间为1.9~0.4 Ma [60 ] Pseudopodoces humilis 青藏高原、柴达木盆地周边高山 在9.9~7.7 Ma期间地山雀就与与其他山雀分化开来,并出现适应高海拔性状 [61 ] Phylloscopus, Seicercus 亚洲 喜马拉雅山脉分布类群在7 Ma加速进化 [62 -63 ] Perdix 西藏、甘肃、青海、四川、新疆 高原类群出现于3.63 (4.52~2.75) Ma [64 ] Pantholops hodgsonii 西藏、新疆、青海 上新世-更新世就已经出现在青藏高原 [65 ] Procapra picticaudata 西藏、四川、甘肃、青海、新疆 种内分化时间约2.2~4.4 Ma [66 ] Rhinopithecus 四川、甘肃、陕西、贵州、云南、缅甸、越南 北方类群川金丝猴、黔金丝猴与喜马拉雅类群滇金丝猴、缅甸金丝猴分化时间约1.6 Ma [67 ] Ochotona 北半球 分布于青藏高原的高山类群和灌丛草原类群与低海拔泛北极分布类群分化时间大约为2.4 Ma [68 ] Apodemus draco 四川、云南、甘肃、青海、陕西、湖北、湖南、福建、台湾 2.2 Ma (3.8~0.87 Ma)分化形成,后来由于高原的阶段性隆升以及冰川作用种系及遗传结构更加复杂 [69 -70 ] Eothenomys 东亚 高原种类与低海拔种类分裂时间约2.7 Ma [71 ] Schizothoracinae 青藏高原及周边地区 在晚中新世晚期或上新世裂腹鱼亚科鱼类就已经出现在青藏高原及周边水系中,在晚更新世期间种内分化加速,不同海拔之间的类群产生等级分化,种间多样性增加,第四纪以来青藏高原隆升导致的水系改组是推动裂腹鱼亚科鱼类演化的原动力 [31 ,76 -78 ]
3 讨论 最近,青藏高原古地貌重建工作取得了丰硕的成果(图3 ),但是各种指标重建结果往往存在矛盾(图4 )[25 ,79 -83 ] .同位素重建结果表明高原南部在古近纪早期就存在安第斯型的巨大山系[84 -85 ] ;古生物研究发现高原腹地从中始新世到晚渐新世都存在东西向的大峡谷,该峡谷底部海拔在2 000 m左右,两侧为高大的冈底斯山脉和羌塘山脉[82 ,86 -88 ] ,动植物如巨犀(Paraceratherium )[89 ] 、攀鲈(Eoanabas )[90 -91 ] 、臭椿(Ailanthus )[88 ] 、椿榆(Cedrelospermum )[92 ] 、兔耳果(Lagokarpos )[93 ] 等在古近纪都可以通过青藏地区峡谷与周边动植物区系交流,新近纪之前的青藏地区虽然地形复杂,但是尚未隆升成今天的高原;横断山区某些区域则在晚始新世-早渐新世之交就已经获得现今的海拔高度[94 -95 ] ;高原北部和喜马拉雅山脉则在新近纪形成现在的地貌格局[25 ,96 -97 ] .古生物研究也发现青藏高原在晚上新世就已经存在高寒生境和冰缘环境,成为第四纪冰期动物忍耐寒冷环境的“训练营”,第四纪冰期来临之后这些适应寒冷环境的生物才扩散至北半球高纬度地区[34 ,98 -99 ] .因此,高原及周边地区高寒生物类群的起源时间表明在渐新世青藏高原部分区域就已经出现高寒生态系统,晚中新世以来青藏高原地貌与气候继续发生剧烈变化,促使这些物种的祖先类群迁入或就地演化积累适应高寒环境的基因和形态,并最终演化成适应高寒生境的新物种[25 ] .高寒区物种在晚上新世的加速分化可能与高原巨大山系和水系、冰川活动等造成的地理隔离有关[26 ] .随着第四纪冰期的到来,青藏高原及周边地区的高寒生物区也开始与北半球高纬度地区存在密切联系[34 ,98 -99 ] .同时,演化生物学研究也发现在第四纪冰期期间并未形成覆盖整个青藏高原的大冰盖,高原面上仍存在很多生物避难所[26 ] . ...
Evolutionary assembly of flowering plants into sky islands
1
2021
... 高寒生物区是分布于树线和雪线之间的独特生态系统,遍布全球各大洲高山和高原之上[26 ,32 ] .高寒生物区自然环境极端恶劣,物种为适应环境进化出独特的抗寒、抗旱、耐低温、抗紫外线辐射、抗缺氧等性状,并在这些极端环境中经过悠久的演化历史最终存活下来[21 ,26 ] ,是获取抗性基因的重要宝库.高寒区生物是适应极端环境的产物,对环境变化非常敏感.近年来全球变暖加剧了高寒区生物灭绝风险,使得该区域成为全球生物多样性保护研究中重点关注的区域.同时,高寒区生物是山脉或高原隆升到一定海拔后的产物[33 ] ,研究高寒区生物多样性的起源和演化过程,不仅可以认识高寒区生物多样性的形成过程,还可以预测未来气候变化对高寒区生物多样性的影响,同时也能够推断高山、高原地貌和气候的演化历史[20 ,23 -24 ,34 ] . ...
Biodiversity from mountain building
1
2013
... 高寒生物区是分布于树线和雪线之间的独特生态系统,遍布全球各大洲高山和高原之上[26 ,32 ] .高寒生物区自然环境极端恶劣,物种为适应环境进化出独特的抗寒、抗旱、耐低温、抗紫外线辐射、抗缺氧等性状,并在这些极端环境中经过悠久的演化历史最终存活下来[21 ,26 ] ,是获取抗性基因的重要宝库.高寒区生物是适应极端环境的产物,对环境变化非常敏感.近年来全球变暖加剧了高寒区生物灭绝风险,使得该区域成为全球生物多样性保护研究中重点关注的区域.同时,高寒区生物是山脉或高原隆升到一定海拔后的产物[33 ] ,研究高寒区生物多样性的起源和演化过程,不仅可以认识高寒区生物多样性的形成过程,还可以预测未来气候变化对高寒区生物多样性的影响,同时也能够推断高山、高原地貌和气候的演化历史[20 ,23 -24 ,34 ] . ...
Cenozoic vertebrate evolution and paleoenvironment in Tibetan Plateau: Progress and prospects
4
2015
... 高寒生物区是分布于树线和雪线之间的独特生态系统,遍布全球各大洲高山和高原之上[26 ,32 ] .高寒生物区自然环境极端恶劣,物种为适应环境进化出独特的抗寒、抗旱、耐低温、抗紫外线辐射、抗缺氧等性状,并在这些极端环境中经过悠久的演化历史最终存活下来[21 ,26 ] ,是获取抗性基因的重要宝库.高寒区生物是适应极端环境的产物,对环境变化非常敏感.近年来全球变暖加剧了高寒区生物灭绝风险,使得该区域成为全球生物多样性保护研究中重点关注的区域.同时,高寒区生物是山脉或高原隆升到一定海拔后的产物[33 ] ,研究高寒区生物多样性的起源和演化过程,不仅可以认识高寒区生物多样性的形成过程,还可以预测未来气候变化对高寒区生物多样性的影响,同时也能够推断高山、高原地貌和气候的演化历史[20 ,23 -24 ,34 ] . ...
... 最近,青藏高原古地貌重建工作取得了丰硕的成果(图3 ),但是各种指标重建结果往往存在矛盾(图4 )[25 ,79 -83 ] .同位素重建结果表明高原南部在古近纪早期就存在安第斯型的巨大山系[84 -85 ] ;古生物研究发现高原腹地从中始新世到晚渐新世都存在东西向的大峡谷,该峡谷底部海拔在2 000 m左右,两侧为高大的冈底斯山脉和羌塘山脉[82 ,86 -88 ] ,动植物如巨犀(Paraceratherium )[89 ] 、攀鲈(Eoanabas )[90 -91 ] 、臭椿(Ailanthus )[88 ] 、椿榆(Cedrelospermum )[92 ] 、兔耳果(Lagokarpos )[93 ] 等在古近纪都可以通过青藏地区峡谷与周边动植物区系交流,新近纪之前的青藏地区虽然地形复杂,但是尚未隆升成今天的高原;横断山区某些区域则在晚始新世-早渐新世之交就已经获得现今的海拔高度[94 -95 ] ;高原北部和喜马拉雅山脉则在新近纪形成现在的地貌格局[25 ,96 -97 ] .古生物研究也发现青藏高原在晚上新世就已经存在高寒生境和冰缘环境,成为第四纪冰期动物忍耐寒冷环境的“训练营”,第四纪冰期来临之后这些适应寒冷环境的生物才扩散至北半球高纬度地区[34 ,98 -99 ] .因此,高原及周边地区高寒生物类群的起源时间表明在渐新世青藏高原部分区域就已经出现高寒生态系统,晚中新世以来青藏高原地貌与气候继续发生剧烈变化,促使这些物种的祖先类群迁入或就地演化积累适应高寒环境的基因和形态,并最终演化成适应高寒生境的新物种[25 ] .高寒区物种在晚上新世的加速分化可能与高原巨大山系和水系、冰川活动等造成的地理隔离有关[26 ] .随着第四纪冰期的到来,青藏高原及周边地区的高寒生物区也开始与北半球高纬度地区存在密切联系[34 ,98 -99 ] .同时,演化生物学研究也发现在第四纪冰期期间并未形成覆盖整个青藏高原的大冰盖,高原面上仍存在很多生物避难所[26 ] . ...
... [34 ,98 -99 ].同时,演化生物学研究也发现在第四纪冰期期间并未形成覆盖整个青藏高原的大冰盖,高原面上仍存在很多生物避难所[26 ] . ...
... ]、[
34 ]、[
82 ]、[
87 ]、[
100 ]修改)
North-south paleoelevation profiles on the Tibetan Plateau (Revised from References [25],[34],[82],[87],[100]) Fig.4 ![]()
随着DNA测序技术和比较系统学方法的发展,大量涌现的青藏高寒区生物地理学研究为我们理解高原隆升过程及其气候效应提供了生物演化方面的证据[25 ] .但是,演化生物学家通常仅对单一特定类群的系统发育过程开展研究,很少进行系统的总结工作[25 ] ,对某一类群遗传数据的解释也往往与最新的青藏高原隆升和气候变化研究进展脱节,不能很好地将最新的关于高原隆升和气候变化的研究与生物类群演化联系在一起[101 ] ,同时也没有认识到青藏高原的隆升过程是阶段性的,没有对青藏高原的不同组成部分进行区分[26 ,53 ,81 ] .地质学研究大多认为高原地貌格局形成于中新世之前,但是综合近年来的生物地理学研究结果可以发现,青藏高寒生物类群的分化时间多集中在晚中新世,种内分化加速和多样性增加主要发生在晚上新世到第四纪期间(图5 ),这些不同类群在分化时间上的巧合可能反映青藏高寒区生物演化受到晚中新世以来构造和气候事件的影响,但是并未引起地质学家足够的重视[25 ] . ...
On the monkeys of the Amazon
1
1854
... 生物地理学主要研究生物的时空分布格局以及性状演化过程及其形成原因和机制.从1852年开始,生物地理学家就开始寻找和总结动植物类群的分布模式,推断其历史分布区演化,并试图将这些分布模式与影响其演化的地质过程联系起来[35 ] .随着DNA测序技术的发展,研究者对标本进行DNA测序,基于宽松分子钟模型或使用年代和鉴定准确的化石记录来校正分子钟,使用最大似然法和贝叶斯法构建系统树,估算物种演化的分支时间和演化历史[18 ] ,通过现生类群的分布格局来推断祖先类群的地理分布.扩散-灭绝-进化分支(dispersal-extinction-cladogenesis)模型是目前最常使用的模型,使用最大似然法和贝叶斯法推断物种的扩散和灭绝历史,用隔离分化或支序发生来解释不同区域间类群分布格局[26 ,36 ] .同时,利用比较系统发育学方法还可以估算物种多样化速率,推断物种分布格局和多样性演化历史,探讨构造事件和气候变化对多样性形成和演化过程的影响[26 ] . ...
A likelihood framework for inferring the evolution of geographic range on phylogenetic trees
1
2005
... 生物地理学主要研究生物的时空分布格局以及性状演化过程及其形成原因和机制.从1852年开始,生物地理学家就开始寻找和总结动植物类群的分布模式,推断其历史分布区演化,并试图将这些分布模式与影响其演化的地质过程联系起来[35 ] .随着DNA测序技术的发展,研究者对标本进行DNA测序,基于宽松分子钟模型或使用年代和鉴定准确的化石记录来校正分子钟,使用最大似然法和贝叶斯法构建系统树,估算物种演化的分支时间和演化历史[18 ] ,通过现生类群的分布格局来推断祖先类群的地理分布.扩散-灭绝-进化分支(dispersal-extinction-cladogenesis)模型是目前最常使用的模型,使用最大似然法和贝叶斯法推断物种的扩散和灭绝历史,用隔离分化或支序发生来解释不同区域间类群分布格局[26 ,36 ] .同时,利用比较系统发育学方法还可以估算物种多样化速率,推断物种分布格局和多样性演化历史,探讨构造事件和气候变化对多样性形成和演化过程的影响[26 ] . ...
青藏高原西部阿汝冰芯记录的近100 a气温变化研究
1
2021
... 青藏高原平均海拔超过4 500 m,是全球中低纬冰川分布最为集中的区域[37 ] ,被誉为世界“第三极”[38 -39 ] ,高寒生物区分布广泛,对气候变化反映敏感[40 ] .近年来,全球变暖加剧,青藏高原及周边地区升温尤其明显[41 -42 ] ,高寒区生物生存压力加大.对青藏高寒区生物开展生物地理学研究有助于探讨青藏高寒区生物多样性起源演化,并预测未来多样性演化对全球变化的响应.此外,最近的研究表明青藏高原东南缘高寒植物多样性在早渐新世就开始积累,是现存高寒生物区中起源最早的[26 ] ,远比安第斯山脉(约3.5 Ma)[43 ] 、阿尔卑斯山脉(<1.75 Ma)[44 ] 、新西兰南阿尔卑斯山脉(<5 Ma)[45 ] 、东非高山(8~7 Ma)[46 ] 高寒植物群起源地早.青藏高寒区生物多样性演化、物种扩散、分化与灭绝受到高原阶段性隆升及其气候效应的影响,对其开展研究能够进一步理解高原隆升过程及其如何对生物多样性演化产生影响. ...
青藏高原西部阿汝冰芯记录的近100 a气温变化研究
1
2021
... 青藏高原平均海拔超过4 500 m,是全球中低纬冰川分布最为集中的区域[37 ] ,被誉为世界“第三极”[38 -39 ] ,高寒生物区分布广泛,对气候变化反映敏感[40 ] .近年来,全球变暖加剧,青藏高原及周边地区升温尤其明显[41 -42 ] ,高寒区生物生存压力加大.对青藏高寒区生物开展生物地理学研究有助于探讨青藏高寒区生物多样性起源演化,并预测未来多样性演化对全球变化的响应.此外,最近的研究表明青藏高原东南缘高寒植物多样性在早渐新世就开始积累,是现存高寒生物区中起源最早的[26 ] ,远比安第斯山脉(约3.5 Ma)[43 ] 、阿尔卑斯山脉(<1.75 Ma)[44 ] 、新西兰南阿尔卑斯山脉(<5 Ma)[45 ] 、东非高山(8~7 Ma)[46 ] 高寒植物群起源地早.青藏高寒区生物多样性演化、物种扩散、分化与灭绝受到高原阶段性隆升及其气候效应的影响,对其开展研究能够进一步理解高原隆升过程及其如何对生物多样性演化产生影响. ...
1
1955
... 青藏高原平均海拔超过4 500 m,是全球中低纬冰川分布最为集中的区域[37 ] ,被誉为世界“第三极”[38 -39 ] ,高寒生物区分布广泛,对气候变化反映敏感[40 ] .近年来,全球变暖加剧,青藏高原及周边地区升温尤其明显[41 -42 ] ,高寒区生物生存压力加大.对青藏高寒区生物开展生物地理学研究有助于探讨青藏高寒区生物多样性起源演化,并预测未来多样性演化对全球变化的响应.此外,最近的研究表明青藏高原东南缘高寒植物多样性在早渐新世就开始积累,是现存高寒生物区中起源最早的[26 ] ,远比安第斯山脉(约3.5 Ma)[43 ] 、阿尔卑斯山脉(<1.75 Ma)[44 ] 、新西兰南阿尔卑斯山脉(<5 Ma)[45 ] 、东非高山(8~7 Ma)[46 ] 高寒植物群起源地早.青藏高寒区生物多样性演化、物种扩散、分化与灭绝受到高原阶段性隆升及其气候效应的影响,对其开展研究能够进一步理解高原隆升过程及其如何对生物多样性演化产生影响. ...
China: the third pole
1
2008
... 青藏高原平均海拔超过4 500 m,是全球中低纬冰川分布最为集中的区域[37 ] ,被誉为世界“第三极”[38 -39 ] ,高寒生物区分布广泛,对气候变化反映敏感[40 ] .近年来,全球变暖加剧,青藏高原及周边地区升温尤其明显[41 -42 ] ,高寒区生物生存压力加大.对青藏高寒区生物开展生物地理学研究有助于探讨青藏高寒区生物多样性起源演化,并预测未来多样性演化对全球变化的响应.此外,最近的研究表明青藏高原东南缘高寒植物多样性在早渐新世就开始积累,是现存高寒生物区中起源最早的[26 ] ,远比安第斯山脉(约3.5 Ma)[43 ] 、阿尔卑斯山脉(<1.75 Ma)[44 ] 、新西兰南阿尔卑斯山脉(<5 Ma)[45 ] 、东非高山(8~7 Ma)[46 ] 高寒植物群起源地早.青藏高寒区生物多样性演化、物种扩散、分化与灭绝受到高原阶段性隆升及其气候效应的影响,对其开展研究能够进一步理解高原隆升过程及其如何对生物多样性演化产生影响. ...
青藏高原海拔要素对温度、降水和气候型分布格局的影响
1
2019
... 青藏高原平均海拔超过4 500 m,是全球中低纬冰川分布最为集中的区域[37 ] ,被誉为世界“第三极”[38 -39 ] ,高寒生物区分布广泛,对气候变化反映敏感[40 ] .近年来,全球变暖加剧,青藏高原及周边地区升温尤其明显[41 -42 ] ,高寒区生物生存压力加大.对青藏高寒区生物开展生物地理学研究有助于探讨青藏高寒区生物多样性起源演化,并预测未来多样性演化对全球变化的响应.此外,最近的研究表明青藏高原东南缘高寒植物多样性在早渐新世就开始积累,是现存高寒生物区中起源最早的[26 ] ,远比安第斯山脉(约3.5 Ma)[43 ] 、阿尔卑斯山脉(<1.75 Ma)[44 ] 、新西兰南阿尔卑斯山脉(<5 Ma)[45 ] 、东非高山(8~7 Ma)[46 ] 高寒植物群起源地早.青藏高寒区生物多样性演化、物种扩散、分化与灭绝受到高原阶段性隆升及其气候效应的影响,对其开展研究能够进一步理解高原隆升过程及其如何对生物多样性演化产生影响. ...
青藏高原海拔要素对温度、降水和气候型分布格局的影响
1
2019
... 青藏高原平均海拔超过4 500 m,是全球中低纬冰川分布最为集中的区域[37 ] ,被誉为世界“第三极”[38 -39 ] ,高寒生物区分布广泛,对气候变化反映敏感[40 ] .近年来,全球变暖加剧,青藏高原及周边地区升温尤其明显[41 -42 ] ,高寒区生物生存压力加大.对青藏高寒区生物开展生物地理学研究有助于探讨青藏高寒区生物多样性起源演化,并预测未来多样性演化对全球变化的响应.此外,最近的研究表明青藏高原东南缘高寒植物多样性在早渐新世就开始积累,是现存高寒生物区中起源最早的[26 ] ,远比安第斯山脉(约3.5 Ma)[43 ] 、阿尔卑斯山脉(<1.75 Ma)[44 ] 、新西兰南阿尔卑斯山脉(<5 Ma)[45 ] 、东非高山(8~7 Ma)[46 ] 高寒植物群起源地早.青藏高寒区生物多样性演化、物种扩散、分化与灭绝受到高原阶段性隆升及其气候效应的影响,对其开展研究能够进一步理解高原隆升过程及其如何对生物多样性演化产生影响. ...
Species interactions slow warming-induced upward shifts of treelines on the Tibetan Plateau
1
2016
... 青藏高原平均海拔超过4 500 m,是全球中低纬冰川分布最为集中的区域[37 ] ,被誉为世界“第三极”[38 -39 ] ,高寒生物区分布广泛,对气候变化反映敏感[40 ] .近年来,全球变暖加剧,青藏高原及周边地区升温尤其明显[41 -42 ] ,高寒区生物生存压力加大.对青藏高寒区生物开展生物地理学研究有助于探讨青藏高寒区生物多样性起源演化,并预测未来多样性演化对全球变化的响应.此外,最近的研究表明青藏高原东南缘高寒植物多样性在早渐新世就开始积累,是现存高寒生物区中起源最早的[26 ] ,远比安第斯山脉(约3.5 Ma)[43 ] 、阿尔卑斯山脉(<1.75 Ma)[44 ] 、新西兰南阿尔卑斯山脉(<5 Ma)[45 ] 、东非高山(8~7 Ma)[46 ] 高寒植物群起源地早.青藏高寒区生物多样性演化、物种扩散、分化与灭绝受到高原阶段性隆升及其气候效应的影响,对其开展研究能够进一步理解高原隆升过程及其如何对生物多样性演化产生影响. ...
Multispherical interactions and their effects on the Tibetan Plateau’s earth system: a review of the recent researches
1
2015
... 青藏高原平均海拔超过4 500 m,是全球中低纬冰川分布最为集中的区域[37 ] ,被誉为世界“第三极”[38 -39 ] ,高寒生物区分布广泛,对气候变化反映敏感[40 ] .近年来,全球变暖加剧,青藏高原及周边地区升温尤其明显[41 -42 ] ,高寒区生物生存压力加大.对青藏高寒区生物开展生物地理学研究有助于探讨青藏高寒区生物多样性起源演化,并预测未来多样性演化对全球变化的响应.此外,最近的研究表明青藏高原东南缘高寒植物多样性在早渐新世就开始积累,是现存高寒生物区中起源最早的[26 ] ,远比安第斯山脉(约3.5 Ma)[43 ] 、阿尔卑斯山脉(<1.75 Ma)[44 ] 、新西兰南阿尔卑斯山脉(<5 Ma)[45 ] 、东非高山(8~7 Ma)[46 ] 高寒植物群起源地早.青藏高寒区生物多样性演化、物种扩散、分化与灭绝受到高原阶段性隆升及其气候效应的影响,对其开展研究能够进一步理解高原隆升过程及其如何对生物多样性演化产生影响. ...
Páramo is the world's fastest evolving and coolest biodiversity hotspot
1
2013
... 青藏高原平均海拔超过4 500 m,是全球中低纬冰川分布最为集中的区域[37 ] ,被誉为世界“第三极”[38 -39 ] ,高寒生物区分布广泛,对气候变化反映敏感[40 ] .近年来,全球变暖加剧,青藏高原及周边地区升温尤其明显[41 -42 ] ,高寒区生物生存压力加大.对青藏高寒区生物开展生物地理学研究有助于探讨青藏高寒区生物多样性起源演化,并预测未来多样性演化对全球变化的响应.此外,最近的研究表明青藏高原东南缘高寒植物多样性在早渐新世就开始积累,是现存高寒生物区中起源最早的[26 ] ,远比安第斯山脉(约3.5 Ma)[43 ] 、阿尔卑斯山脉(<1.75 Ma)[44 ] 、新西兰南阿尔卑斯山脉(<5 Ma)[45 ] 、东非高山(8~7 Ma)[46 ] 高寒植物群起源地早.青藏高寒区生物多样性演化、物种扩散、分化与灭绝受到高原阶段性隆升及其气候效应的影响,对其开展研究能够进一步理解高原隆升过程及其如何对生物多样性演化产生影响. ...
Molecular evidence for multiple diversification patterns of alpine plants in Mediterranean Europe
1
2003
... 青藏高原平均海拔超过4 500 m,是全球中低纬冰川分布最为集中的区域[37 ] ,被誉为世界“第三极”[38 -39 ] ,高寒生物区分布广泛,对气候变化反映敏感[40 ] .近年来,全球变暖加剧,青藏高原及周边地区升温尤其明显[41 -42 ] ,高寒区生物生存压力加大.对青藏高寒区生物开展生物地理学研究有助于探讨青藏高寒区生物多样性起源演化,并预测未来多样性演化对全球变化的响应.此外,最近的研究表明青藏高原东南缘高寒植物多样性在早渐新世就开始积累,是现存高寒生物区中起源最早的[26 ] ,远比安第斯山脉(约3.5 Ma)[43 ] 、阿尔卑斯山脉(<1.75 Ma)[44 ] 、新西兰南阿尔卑斯山脉(<5 Ma)[45 ] 、东非高山(8~7 Ma)[46 ] 高寒植物群起源地早.青藏高寒区生物多样性演化、物种扩散、分化与灭绝受到高原阶段性隆升及其气候效应的影响,对其开展研究能够进一步理解高原隆升过程及其如何对生物多样性演化产生影响. ...
Evolution of New Zealand alpine and open-habitat plant species during the late Cenozoic
1
2013
... 青藏高原平均海拔超过4 500 m,是全球中低纬冰川分布最为集中的区域[37 ] ,被誉为世界“第三极”[38 -39 ] ,高寒生物区分布广泛,对气候变化反映敏感[40 ] .近年来,全球变暖加剧,青藏高原及周边地区升温尤其明显[41 -42 ] ,高寒区生物生存压力加大.对青藏高寒区生物开展生物地理学研究有助于探讨青藏高寒区生物多样性起源演化,并预测未来多样性演化对全球变化的响应.此外,最近的研究表明青藏高原东南缘高寒植物多样性在早渐新世就开始积累,是现存高寒生物区中起源最早的[26 ] ,远比安第斯山脉(约3.5 Ma)[43 ] 、阿尔卑斯山脉(<1.75 Ma)[44 ] 、新西兰南阿尔卑斯山脉(<5 Ma)[45 ] 、东非高山(8~7 Ma)[46 ] 高寒植物群起源地早.青藏高寒区生物多样性演化、物种扩散、分化与灭绝受到高原阶段性隆升及其气候效应的影响,对其开展研究能够进一步理解高原隆升过程及其如何对生物多样性演化产生影响. ...
The evolution of African plant diversity
1
2014
... 青藏高原平均海拔超过4 500 m,是全球中低纬冰川分布最为集中的区域[37 ] ,被誉为世界“第三极”[38 -39 ] ,高寒生物区分布广泛,对气候变化反映敏感[40 ] .近年来,全球变暖加剧,青藏高原及周边地区升温尤其明显[41 -42 ] ,高寒区生物生存压力加大.对青藏高寒区生物开展生物地理学研究有助于探讨青藏高寒区生物多样性起源演化,并预测未来多样性演化对全球变化的响应.此外,最近的研究表明青藏高原东南缘高寒植物多样性在早渐新世就开始积累,是现存高寒生物区中起源最早的[26 ] ,远比安第斯山脉(约3.5 Ma)[43 ] 、阿尔卑斯山脉(<1.75 Ma)[44 ] 、新西兰南阿尔卑斯山脉(<5 Ma)[45 ] 、东非高山(8~7 Ma)[46 ] 高寒植物群起源地早.青藏高寒区生物多样性演化、物种扩散、分化与灭绝受到高原阶段性隆升及其气候效应的影响,对其开展研究能够进一步理解高原隆升过程及其如何对生物多样性演化产生影响. ...
Molecular evidence for the sister relationship of the eastern Asia-North American intercontinental species pair in the Podophyllum group (Berberidaceae)
2
2002
... 植物中的桃儿七(Sinopodophyllum hexandrum )分布于青藏高原东部,系统发育关系推断其出现在青藏高原的时间约为(6.52±1.89) Ma,但是适应高寒性状出现的时间为晚上新世[47 ] .菊科毛冠菊属(Nannoglottis )植物中生活于高山灌丛和亚高山针叶林的类群形态明显不同,两者的分化时间约为3.4 Ma[48 ] .分布于中国西北干旱区和青藏高原北部及东北部的红砂(Reaumuria soongarica ),在6.3~2.0 Ma期间演化出适应高寒生境的类群,高原北部分布类群也在3.5~1.1 Ma期间响应构造活动造成的地理隔离而发生分化[49 ] .红门兰亚族(Orchidinae)六大分支在多样化的时空序列上表现出明显的差异,其生活在青藏高原地区的特有种大多数是在过去6 Ma里进化而来的[50 ] .中华山蓼(Oxyria sinensis )分布于青藏高原东南部海拔1 600~3 800 m的高山上,约13.8 Ma演化出高山类群,第四纪期间种内多样化增加[51 ] .小子圆柏(Juniperus microsperma )是青藏高原特有种,现在仅生长于西藏波密地区,高原南部在中新世早期构造隆升造成的物种隔离使其与其他刺柏种群发生分化[52 ] .横断山区杜鹃花属(Rhododendron )多样性在晚中新世(约8 Ma)开始快速增加,对青藏高原不同区域杜鹃花属的演化历史进行定量化比较研究,发现横断山区在晚中新世的快速隆升促进生物多样性的形成[53 ] .对喜马拉雅-横断山分布的高山姜类的研究发现,在早渐新世(约32 Ma)象牙参属(Roscoea )和距药姜属(Cautleya )就与其姐妹类群分化开来,而象牙参属在横断山区与喜马拉雅山脉的两个分支大概在渐新世-中新世之交分化[54 ] . ...
... Biological evolution in the alpine region of Tibetan Plateau
Table 1 种属 分布区域 适应高海拔类群分化时间或多样性增加时间 文献来源 Sinopodophyllum hexandrum 青藏高原东部 在约(6.52±1.89) Ma期间出现在青藏高原东部,但是直到3~2 Ma以来才演化出适应高山环境的形态 [47 ] Nannoglottis 青藏高原 在23~32 Ma期间出现在青藏高原,随后多样性增高,其高山灌丛和针叶林生活类型分支时间约为3.4 Ma [48 ] Reaumuria soongarica 西北干旱区、青藏高原北部 高原内部分布类群演化时间2.0~6.3 Ma,高原东北缘与高原西北类群分化时间1.1~3.5 Ma [49 ] Orchidinae 青藏高原及喜马拉雅山脉 青藏高原特有种大多是在过去6 Ma里进化而来 [50 ] Oxyria sinensis 青藏高原东南部 高山类群出现时间7.54~24.36 Ma,种内多样化增加时间1.74~0.86 Ma [51 ] Juniperus microsperma 西藏波密 早中新世21.6~14 Ma期间与其他种群分化 [52 ] Rhododendron 北半球 横断山区概属多样性在晚中新世(约8 Ma)快速增加 [53 ] Roscoea, Cautleya 喜马拉雅-横断山区 在早渐新世(~32 Ma)两属出现,Roscoea在横断山区与喜马拉雅山脉的两个分支大概在渐新世-中新世之交分化 [54 ] Nanorana parkeri 青藏高原南部 高原东部和西部类群分化时间1.4~3.7 Ma [55 ] Paini 南亚、东南亚、青藏高原 在渐新世早期棘蛙族高原-喜马拉雅类群与中国第二阶梯上生活的类群分化,随后在在26.1~12.5 Ma高原生活类群与喜马拉雅山脉生活类群分化,高原内部生活类群在晚中新世到上新世期间分化速率加快 [22 ] Bufo tibetanus 青藏高原、四川、甘肃、陕西等 高原种类形成于上新世期间 [56 ] Phrynocephalus 亚洲干旱区 在7.2~13.0 Ma期间演化出胎生类群,3.2~7.3 Ma期间种间多样性增加 [57 ] Phrynocephalus erythrurus 羌塘盆地 在3.67 Ma左右北部类群与南部类群发生分化,约2.76 Ma北部类群内部进一步分化 [58 ] Gnaptorina 青藏高原及周边高海拔地区 始新世起源于青藏高原东南缘 [18 -19 ] Coelotinae 北半球温带、亚热带 晚始新世高原东南缘隆升阻碍南方隙蛛向北扩散,渐新世高原中部和北部隆起导致北方隙蛛早期多样化进程中的灭绝事件 [59 ] Montifringilla, Onychostruthus, Pyrgilauda 青藏高原 雪雀的共同祖先在约14 Ma就应经生活在青藏高原,并进化出适应高海拔极端环境的性状 [21 ] Parus major 亚洲、欧洲 蒙古分布类群与其他种类分化时间为2.8~0.7 Ma,喜马拉雅山脉东部与中国中东部类群分化时间为1.9~0.4 Ma [60 ] Pseudopodoces humilis 青藏高原、柴达木盆地周边高山 在9.9~7.7 Ma期间地山雀就与与其他山雀分化开来,并出现适应高海拔性状 [61 ] Phylloscopus, Seicercus 亚洲 喜马拉雅山脉分布类群在7 Ma加速进化 [62 -63 ] Perdix 西藏、甘肃、青海、四川、新疆 高原类群出现于3.63 (4.52~2.75) Ma [64 ] Pantholops hodgsonii 西藏、新疆、青海 上新世-更新世就已经出现在青藏高原 [65 ] Procapra picticaudata 西藏、四川、甘肃、青海、新疆 种内分化时间约2.2~4.4 Ma [66 ] Rhinopithecus 四川、甘肃、陕西、贵州、云南、缅甸、越南 北方类群川金丝猴、黔金丝猴与喜马拉雅类群滇金丝猴、缅甸金丝猴分化时间约1.6 Ma [67 ] Ochotona 北半球 分布于青藏高原的高山类群和灌丛草原类群与低海拔泛北极分布类群分化时间大约为2.4 Ma [68 ] Apodemus draco 四川、云南、甘肃、青海、陕西、湖北、湖南、福建、台湾 2.2 Ma (3.8~0.87 Ma)分化形成,后来由于高原的阶段性隆升以及冰川作用种系及遗传结构更加复杂 [69 -70 ] Eothenomys 东亚 高原种类与低海拔种类分裂时间约2.7 Ma [71 ] Schizothoracinae 青藏高原及周边地区 在晚中新世晚期或上新世裂腹鱼亚科鱼类就已经出现在青藏高原及周边水系中,在晚更新世期间种内分化加速,不同海拔之间的类群产生等级分化,种间多样性增加,第四纪以来青藏高原隆升导致的水系改组是推动裂腹鱼亚科鱼类演化的原动力 [31 ,76 -78 ]
3 讨论 最近,青藏高原古地貌重建工作取得了丰硕的成果(图3 ),但是各种指标重建结果往往存在矛盾(图4 )[25 ,79 -83 ] .同位素重建结果表明高原南部在古近纪早期就存在安第斯型的巨大山系[84 -85 ] ;古生物研究发现高原腹地从中始新世到晚渐新世都存在东西向的大峡谷,该峡谷底部海拔在2 000 m左右,两侧为高大的冈底斯山脉和羌塘山脉[82 ,86 -88 ] ,动植物如巨犀(Paraceratherium )[89 ] 、攀鲈(Eoanabas )[90 -91 ] 、臭椿(Ailanthus )[88 ] 、椿榆(Cedrelospermum )[92 ] 、兔耳果(Lagokarpos )[93 ] 等在古近纪都可以通过青藏地区峡谷与周边动植物区系交流,新近纪之前的青藏地区虽然地形复杂,但是尚未隆升成今天的高原;横断山区某些区域则在晚始新世-早渐新世之交就已经获得现今的海拔高度[94 -95 ] ;高原北部和喜马拉雅山脉则在新近纪形成现在的地貌格局[25 ,96 -97 ] .古生物研究也发现青藏高原在晚上新世就已经存在高寒生境和冰缘环境,成为第四纪冰期动物忍耐寒冷环境的“训练营”,第四纪冰期来临之后这些适应寒冷环境的生物才扩散至北半球高纬度地区[34 ,98 -99 ] .因此,高原及周边地区高寒生物类群的起源时间表明在渐新世青藏高原部分区域就已经出现高寒生态系统,晚中新世以来青藏高原地貌与气候继续发生剧烈变化,促使这些物种的祖先类群迁入或就地演化积累适应高寒环境的基因和形态,并最终演化成适应高寒生境的新物种[25 ] .高寒区物种在晚上新世的加速分化可能与高原巨大山系和水系、冰川活动等造成的地理隔离有关[26 ] .随着第四纪冰期的到来,青藏高原及周边地区的高寒生物区也开始与北半球高纬度地区存在密切联系[34 ,98 -99 ] .同时,演化生物学研究也发现在第四纪冰期期间并未形成覆盖整个青藏高原的大冰盖,高原面上仍存在很多生物避难所[26 ] . ...
Molecular phylogeny and biogeography of the Qinghai-Tibet Plateau endemic Nannoglottis (Asteraceae)
2
2002
... 植物中的桃儿七(Sinopodophyllum hexandrum )分布于青藏高原东部,系统发育关系推断其出现在青藏高原的时间约为(6.52±1.89) Ma,但是适应高寒性状出现的时间为晚上新世[47 ] .菊科毛冠菊属(Nannoglottis )植物中生活于高山灌丛和亚高山针叶林的类群形态明显不同,两者的分化时间约为3.4 Ma[48 ] .分布于中国西北干旱区和青藏高原北部及东北部的红砂(Reaumuria soongarica ),在6.3~2.0 Ma期间演化出适应高寒生境的类群,高原北部分布类群也在3.5~1.1 Ma期间响应构造活动造成的地理隔离而发生分化[49 ] .红门兰亚族(Orchidinae)六大分支在多样化的时空序列上表现出明显的差异,其生活在青藏高原地区的特有种大多数是在过去6 Ma里进化而来的[50 ] .中华山蓼(Oxyria sinensis )分布于青藏高原东南部海拔1 600~3 800 m的高山上,约13.8 Ma演化出高山类群,第四纪期间种内多样化增加[51 ] .小子圆柏(Juniperus microsperma )是青藏高原特有种,现在仅生长于西藏波密地区,高原南部在中新世早期构造隆升造成的物种隔离使其与其他刺柏种群发生分化[52 ] .横断山区杜鹃花属(Rhododendron )多样性在晚中新世(约8 Ma)开始快速增加,对青藏高原不同区域杜鹃花属的演化历史进行定量化比较研究,发现横断山区在晚中新世的快速隆升促进生物多样性的形成[53 ] .对喜马拉雅-横断山分布的高山姜类的研究发现,在早渐新世(约32 Ma)象牙参属(Roscoea )和距药姜属(Cautleya )就与其姐妹类群分化开来,而象牙参属在横断山区与喜马拉雅山脉的两个分支大概在渐新世-中新世之交分化[54 ] . ...
... Biological evolution in the alpine region of Tibetan Plateau
Table 1 种属 分布区域 适应高海拔类群分化时间或多样性增加时间 文献来源 Sinopodophyllum hexandrum 青藏高原东部 在约(6.52±1.89) Ma期间出现在青藏高原东部,但是直到3~2 Ma以来才演化出适应高山环境的形态 [47 ] Nannoglottis 青藏高原 在23~32 Ma期间出现在青藏高原,随后多样性增高,其高山灌丛和针叶林生活类型分支时间约为3.4 Ma [48 ] Reaumuria soongarica 西北干旱区、青藏高原北部 高原内部分布类群演化时间2.0~6.3 Ma,高原东北缘与高原西北类群分化时间1.1~3.5 Ma [49 ] Orchidinae 青藏高原及喜马拉雅山脉 青藏高原特有种大多是在过去6 Ma里进化而来 [50 ] Oxyria sinensis 青藏高原东南部 高山类群出现时间7.54~24.36 Ma,种内多样化增加时间1.74~0.86 Ma [51 ] Juniperus microsperma 西藏波密 早中新世21.6~14 Ma期间与其他种群分化 [52 ] Rhododendron 北半球 横断山区概属多样性在晚中新世(约8 Ma)快速增加 [53 ] Roscoea, Cautleya 喜马拉雅-横断山区 在早渐新世(~32 Ma)两属出现,Roscoea在横断山区与喜马拉雅山脉的两个分支大概在渐新世-中新世之交分化 [54 ] Nanorana parkeri 青藏高原南部 高原东部和西部类群分化时间1.4~3.7 Ma [55 ] Paini 南亚、东南亚、青藏高原 在渐新世早期棘蛙族高原-喜马拉雅类群与中国第二阶梯上生活的类群分化,随后在在26.1~12.5 Ma高原生活类群与喜马拉雅山脉生活类群分化,高原内部生活类群在晚中新世到上新世期间分化速率加快 [22 ] Bufo tibetanus 青藏高原、四川、甘肃、陕西等 高原种类形成于上新世期间 [56 ] Phrynocephalus 亚洲干旱区 在7.2~13.0 Ma期间演化出胎生类群,3.2~7.3 Ma期间种间多样性增加 [57 ] Phrynocephalus erythrurus 羌塘盆地 在3.67 Ma左右北部类群与南部类群发生分化,约2.76 Ma北部类群内部进一步分化 [58 ] Gnaptorina 青藏高原及周边高海拔地区 始新世起源于青藏高原东南缘 [18 -19 ] Coelotinae 北半球温带、亚热带 晚始新世高原东南缘隆升阻碍南方隙蛛向北扩散,渐新世高原中部和北部隆起导致北方隙蛛早期多样化进程中的灭绝事件 [59 ] Montifringilla, Onychostruthus, Pyrgilauda 青藏高原 雪雀的共同祖先在约14 Ma就应经生活在青藏高原,并进化出适应高海拔极端环境的性状 [21 ] Parus major 亚洲、欧洲 蒙古分布类群与其他种类分化时间为2.8~0.7 Ma,喜马拉雅山脉东部与中国中东部类群分化时间为1.9~0.4 Ma [60 ] Pseudopodoces humilis 青藏高原、柴达木盆地周边高山 在9.9~7.7 Ma期间地山雀就与与其他山雀分化开来,并出现适应高海拔性状 [61 ] Phylloscopus, Seicercus 亚洲 喜马拉雅山脉分布类群在7 Ma加速进化 [62 -63 ] Perdix 西藏、甘肃、青海、四川、新疆 高原类群出现于3.63 (4.52~2.75) Ma [64 ] Pantholops hodgsonii 西藏、新疆、青海 上新世-更新世就已经出现在青藏高原 [65 ] Procapra picticaudata 西藏、四川、甘肃、青海、新疆 种内分化时间约2.2~4.4 Ma [66 ] Rhinopithecus 四川、甘肃、陕西、贵州、云南、缅甸、越南 北方类群川金丝猴、黔金丝猴与喜马拉雅类群滇金丝猴、缅甸金丝猴分化时间约1.6 Ma [67 ] Ochotona 北半球 分布于青藏高原的高山类群和灌丛草原类群与低海拔泛北极分布类群分化时间大约为2.4 Ma [68 ] Apodemus draco 四川、云南、甘肃、青海、陕西、湖北、湖南、福建、台湾 2.2 Ma (3.8~0.87 Ma)分化形成,后来由于高原的阶段性隆升以及冰川作用种系及遗传结构更加复杂 [69 -70 ] Eothenomys 东亚 高原种类与低海拔种类分裂时间约2.7 Ma [71 ] Schizothoracinae 青藏高原及周边地区 在晚中新世晚期或上新世裂腹鱼亚科鱼类就已经出现在青藏高原及周边水系中,在晚更新世期间种内分化加速,不同海拔之间的类群产生等级分化,种间多样性增加,第四纪以来青藏高原隆升导致的水系改组是推动裂腹鱼亚科鱼类演化的原动力 [31 ,76 -78 ]
3 讨论 最近,青藏高原古地貌重建工作取得了丰硕的成果(图3 ),但是各种指标重建结果往往存在矛盾(图4 )[25 ,79 -83 ] .同位素重建结果表明高原南部在古近纪早期就存在安第斯型的巨大山系[84 -85 ] ;古生物研究发现高原腹地从中始新世到晚渐新世都存在东西向的大峡谷,该峡谷底部海拔在2 000 m左右,两侧为高大的冈底斯山脉和羌塘山脉[82 ,86 -88 ] ,动植物如巨犀(Paraceratherium )[89 ] 、攀鲈(Eoanabas )[90 -91 ] 、臭椿(Ailanthus )[88 ] 、椿榆(Cedrelospermum )[92 ] 、兔耳果(Lagokarpos )[93 ] 等在古近纪都可以通过青藏地区峡谷与周边动植物区系交流,新近纪之前的青藏地区虽然地形复杂,但是尚未隆升成今天的高原;横断山区某些区域则在晚始新世-早渐新世之交就已经获得现今的海拔高度[94 -95 ] ;高原北部和喜马拉雅山脉则在新近纪形成现在的地貌格局[25 ,96 -97 ] .古生物研究也发现青藏高原在晚上新世就已经存在高寒生境和冰缘环境,成为第四纪冰期动物忍耐寒冷环境的“训练营”,第四纪冰期来临之后这些适应寒冷环境的生物才扩散至北半球高纬度地区[34 ,98 -99 ] .因此,高原及周边地区高寒生物类群的起源时间表明在渐新世青藏高原部分区域就已经出现高寒生态系统,晚中新世以来青藏高原地貌与气候继续发生剧烈变化,促使这些物种的祖先类群迁入或就地演化积累适应高寒环境的基因和形态,并最终演化成适应高寒生境的新物种[25 ] .高寒区物种在晚上新世的加速分化可能与高原巨大山系和水系、冰川活动等造成的地理隔离有关[26 ] .随着第四纪冰期的到来,青藏高原及周边地区的高寒生物区也开始与北半球高纬度地区存在密切联系[34 ,98 -99 ] .同时,演化生物学研究也发现在第四纪冰期期间并未形成覆盖整个青藏高原的大冰盖,高原面上仍存在很多生物避难所[26 ] . ...
Response of a desert shrub to past geological and climatic change: a phylogeographic study of Reaumuria soongarica (Tamaricaceae) in western China
2
2012
... 植物中的桃儿七(Sinopodophyllum hexandrum )分布于青藏高原东部,系统发育关系推断其出现在青藏高原的时间约为(6.52±1.89) Ma,但是适应高寒性状出现的时间为晚上新世[47 ] .菊科毛冠菊属(Nannoglottis )植物中生活于高山灌丛和亚高山针叶林的类群形态明显不同,两者的分化时间约为3.4 Ma[48 ] .分布于中国西北干旱区和青藏高原北部及东北部的红砂(Reaumuria soongarica ),在6.3~2.0 Ma期间演化出适应高寒生境的类群,高原北部分布类群也在3.5~1.1 Ma期间响应构造活动造成的地理隔离而发生分化[49 ] .红门兰亚族(Orchidinae)六大分支在多样化的时空序列上表现出明显的差异,其生活在青藏高原地区的特有种大多数是在过去6 Ma里进化而来的[50 ] .中华山蓼(Oxyria sinensis )分布于青藏高原东南部海拔1 600~3 800 m的高山上,约13.8 Ma演化出高山类群,第四纪期间种内多样化增加[51 ] .小子圆柏(Juniperus microsperma )是青藏高原特有种,现在仅生长于西藏波密地区,高原南部在中新世早期构造隆升造成的物种隔离使其与其他刺柏种群发生分化[52 ] .横断山区杜鹃花属(Rhododendron )多样性在晚中新世(约8 Ma)开始快速增加,对青藏高原不同区域杜鹃花属的演化历史进行定量化比较研究,发现横断山区在晚中新世的快速隆升促进生物多样性的形成[53 ] .对喜马拉雅-横断山分布的高山姜类的研究发现,在早渐新世(约32 Ma)象牙参属(Roscoea )和距药姜属(Cautleya )就与其姐妹类群分化开来,而象牙参属在横断山区与喜马拉雅山脉的两个分支大概在渐新世-中新世之交分化[54 ] . ...
... Biological evolution in the alpine region of Tibetan Plateau
Table 1 种属 分布区域 适应高海拔类群分化时间或多样性增加时间 文献来源 Sinopodophyllum hexandrum 青藏高原东部 在约(6.52±1.89) Ma期间出现在青藏高原东部,但是直到3~2 Ma以来才演化出适应高山环境的形态 [47 ] Nannoglottis 青藏高原 在23~32 Ma期间出现在青藏高原,随后多样性增高,其高山灌丛和针叶林生活类型分支时间约为3.4 Ma [48 ] Reaumuria soongarica 西北干旱区、青藏高原北部 高原内部分布类群演化时间2.0~6.3 Ma,高原东北缘与高原西北类群分化时间1.1~3.5 Ma [49 ] Orchidinae 青藏高原及喜马拉雅山脉 青藏高原特有种大多是在过去6 Ma里进化而来 [50 ] Oxyria sinensis 青藏高原东南部 高山类群出现时间7.54~24.36 Ma,种内多样化增加时间1.74~0.86 Ma [51 ] Juniperus microsperma 西藏波密 早中新世21.6~14 Ma期间与其他种群分化 [52 ] Rhododendron 北半球 横断山区概属多样性在晚中新世(约8 Ma)快速增加 [53 ] Roscoea, Cautleya 喜马拉雅-横断山区 在早渐新世(~32 Ma)两属出现,Roscoea在横断山区与喜马拉雅山脉的两个分支大概在渐新世-中新世之交分化 [54 ] Nanorana parkeri 青藏高原南部 高原东部和西部类群分化时间1.4~3.7 Ma [55 ] Paini 南亚、东南亚、青藏高原 在渐新世早期棘蛙族高原-喜马拉雅类群与中国第二阶梯上生活的类群分化,随后在在26.1~12.5 Ma高原生活类群与喜马拉雅山脉生活类群分化,高原内部生活类群在晚中新世到上新世期间分化速率加快 [22 ] Bufo tibetanus 青藏高原、四川、甘肃、陕西等 高原种类形成于上新世期间 [56 ] Phrynocephalus 亚洲干旱区 在7.2~13.0 Ma期间演化出胎生类群,3.2~7.3 Ma期间种间多样性增加 [57 ] Phrynocephalus erythrurus 羌塘盆地 在3.67 Ma左右北部类群与南部类群发生分化,约2.76 Ma北部类群内部进一步分化 [58 ] Gnaptorina 青藏高原及周边高海拔地区 始新世起源于青藏高原东南缘 [18 -19 ] Coelotinae 北半球温带、亚热带 晚始新世高原东南缘隆升阻碍南方隙蛛向北扩散,渐新世高原中部和北部隆起导致北方隙蛛早期多样化进程中的灭绝事件 [59 ] Montifringilla, Onychostruthus, Pyrgilauda 青藏高原 雪雀的共同祖先在约14 Ma就应经生活在青藏高原,并进化出适应高海拔极端环境的性状 [21 ] Parus major 亚洲、欧洲 蒙古分布类群与其他种类分化时间为2.8~0.7 Ma,喜马拉雅山脉东部与中国中东部类群分化时间为1.9~0.4 Ma [60 ] Pseudopodoces humilis 青藏高原、柴达木盆地周边高山 在9.9~7.7 Ma期间地山雀就与与其他山雀分化开来,并出现适应高海拔性状 [61 ] Phylloscopus, Seicercus 亚洲 喜马拉雅山脉分布类群在7 Ma加速进化 [62 -63 ] Perdix 西藏、甘肃、青海、四川、新疆 高原类群出现于3.63 (4.52~2.75) Ma [64 ] Pantholops hodgsonii 西藏、新疆、青海 上新世-更新世就已经出现在青藏高原 [65 ] Procapra picticaudata 西藏、四川、甘肃、青海、新疆 种内分化时间约2.2~4.4 Ma [66 ] Rhinopithecus 四川、甘肃、陕西、贵州、云南、缅甸、越南 北方类群川金丝猴、黔金丝猴与喜马拉雅类群滇金丝猴、缅甸金丝猴分化时间约1.6 Ma [67 ] Ochotona 北半球 分布于青藏高原的高山类群和灌丛草原类群与低海拔泛北极分布类群分化时间大约为2.4 Ma [68 ] Apodemus draco 四川、云南、甘肃、青海、陕西、湖北、湖南、福建、台湾 2.2 Ma (3.8~0.87 Ma)分化形成,后来由于高原的阶段性隆升以及冰川作用种系及遗传结构更加复杂 [69 -70 ] Eothenomys 东亚 高原种类与低海拔种类分裂时间约2.7 Ma [71 ] Schizothoracinae 青藏高原及周边地区 在晚中新世晚期或上新世裂腹鱼亚科鱼类就已经出现在青藏高原及周边水系中,在晚更新世期间种内分化加速,不同海拔之间的类群产生等级分化,种间多样性增加,第四纪以来青藏高原隆升导致的水系改组是推动裂腹鱼亚科鱼类演化的原动力 [31 ,76 -78 ]
3 讨论 最近,青藏高原古地貌重建工作取得了丰硕的成果(图3 ),但是各种指标重建结果往往存在矛盾(图4 )[25 ,79 -83 ] .同位素重建结果表明高原南部在古近纪早期就存在安第斯型的巨大山系[84 -85 ] ;古生物研究发现高原腹地从中始新世到晚渐新世都存在东西向的大峡谷,该峡谷底部海拔在2 000 m左右,两侧为高大的冈底斯山脉和羌塘山脉[82 ,86 -88 ] ,动植物如巨犀(Paraceratherium )[89 ] 、攀鲈(Eoanabas )[90 -91 ] 、臭椿(Ailanthus )[88 ] 、椿榆(Cedrelospermum )[92 ] 、兔耳果(Lagokarpos )[93 ] 等在古近纪都可以通过青藏地区峡谷与周边动植物区系交流,新近纪之前的青藏地区虽然地形复杂,但是尚未隆升成今天的高原;横断山区某些区域则在晚始新世-早渐新世之交就已经获得现今的海拔高度[94 -95 ] ;高原北部和喜马拉雅山脉则在新近纪形成现在的地貌格局[25 ,96 -97 ] .古生物研究也发现青藏高原在晚上新世就已经存在高寒生境和冰缘环境,成为第四纪冰期动物忍耐寒冷环境的“训练营”,第四纪冰期来临之后这些适应寒冷环境的生物才扩散至北半球高纬度地区[34 ,98 -99 ] .因此,高原及周边地区高寒生物类群的起源时间表明在渐新世青藏高原部分区域就已经出现高寒生态系统,晚中新世以来青藏高原地貌与气候继续发生剧烈变化,促使这些物种的祖先类群迁入或就地演化积累适应高寒环境的基因和形态,并最终演化成适应高寒生境的新物种[25 ] .高寒区物种在晚上新世的加速分化可能与高原巨大山系和水系、冰川活动等造成的地理隔离有关[26 ] .随着第四纪冰期的到来,青藏高原及周边地区的高寒生物区也开始与北半球高纬度地区存在密切联系[34 ,98 -99 ] .同时,演化生物学研究也发现在第四纪冰期期间并未形成覆盖整个青藏高原的大冰盖,高原面上仍存在很多生物避难所[26 ] . ...
Diversification in Qinghai-Tibet Plateau: Orchidinae (Orchidaceae) clades exhibiting pre-adaptations play critical role
2
2021
... 植物中的桃儿七(Sinopodophyllum hexandrum )分布于青藏高原东部,系统发育关系推断其出现在青藏高原的时间约为(6.52±1.89) Ma,但是适应高寒性状出现的时间为晚上新世[47 ] .菊科毛冠菊属(Nannoglottis )植物中生活于高山灌丛和亚高山针叶林的类群形态明显不同,两者的分化时间约为3.4 Ma[48 ] .分布于中国西北干旱区和青藏高原北部及东北部的红砂(Reaumuria soongarica ),在6.3~2.0 Ma期间演化出适应高寒生境的类群,高原北部分布类群也在3.5~1.1 Ma期间响应构造活动造成的地理隔离而发生分化[49 ] .红门兰亚族(Orchidinae)六大分支在多样化的时空序列上表现出明显的差异,其生活在青藏高原地区的特有种大多数是在过去6 Ma里进化而来的[50 ] .中华山蓼(Oxyria sinensis )分布于青藏高原东南部海拔1 600~3 800 m的高山上,约13.8 Ma演化出高山类群,第四纪期间种内多样化增加[51 ] .小子圆柏(Juniperus microsperma )是青藏高原特有种,现在仅生长于西藏波密地区,高原南部在中新世早期构造隆升造成的物种隔离使其与其他刺柏种群发生分化[52 ] .横断山区杜鹃花属(Rhododendron )多样性在晚中新世(约8 Ma)开始快速增加,对青藏高原不同区域杜鹃花属的演化历史进行定量化比较研究,发现横断山区在晚中新世的快速隆升促进生物多样性的形成[53 ] .对喜马拉雅-横断山分布的高山姜类的研究发现,在早渐新世(约32 Ma)象牙参属(Roscoea )和距药姜属(Cautleya )就与其姐妹类群分化开来,而象牙参属在横断山区与喜马拉雅山脉的两个分支大概在渐新世-中新世之交分化[54 ] . ...
... Biological evolution in the alpine region of Tibetan Plateau
Table 1 种属 分布区域 适应高海拔类群分化时间或多样性增加时间 文献来源 Sinopodophyllum hexandrum 青藏高原东部 在约(6.52±1.89) Ma期间出现在青藏高原东部,但是直到3~2 Ma以来才演化出适应高山环境的形态 [47 ] Nannoglottis 青藏高原 在23~32 Ma期间出现在青藏高原,随后多样性增高,其高山灌丛和针叶林生活类型分支时间约为3.4 Ma [48 ] Reaumuria soongarica 西北干旱区、青藏高原北部 高原内部分布类群演化时间2.0~6.3 Ma,高原东北缘与高原西北类群分化时间1.1~3.5 Ma [49 ] Orchidinae 青藏高原及喜马拉雅山脉 青藏高原特有种大多是在过去6 Ma里进化而来 [50 ] Oxyria sinensis 青藏高原东南部 高山类群出现时间7.54~24.36 Ma,种内多样化增加时间1.74~0.86 Ma [51 ] Juniperus microsperma 西藏波密 早中新世21.6~14 Ma期间与其他种群分化 [52 ] Rhododendron 北半球 横断山区概属多样性在晚中新世(约8 Ma)快速增加 [53 ] Roscoea, Cautleya 喜马拉雅-横断山区 在早渐新世(~32 Ma)两属出现,Roscoea在横断山区与喜马拉雅山脉的两个分支大概在渐新世-中新世之交分化 [54 ] Nanorana parkeri 青藏高原南部 高原东部和西部类群分化时间1.4~3.7 Ma [55 ] Paini 南亚、东南亚、青藏高原 在渐新世早期棘蛙族高原-喜马拉雅类群与中国第二阶梯上生活的类群分化,随后在在26.1~12.5 Ma高原生活类群与喜马拉雅山脉生活类群分化,高原内部生活类群在晚中新世到上新世期间分化速率加快 [22 ] Bufo tibetanus 青藏高原、四川、甘肃、陕西等 高原种类形成于上新世期间 [56 ] Phrynocephalus 亚洲干旱区 在7.2~13.0 Ma期间演化出胎生类群,3.2~7.3 Ma期间种间多样性增加 [57 ] Phrynocephalus erythrurus 羌塘盆地 在3.67 Ma左右北部类群与南部类群发生分化,约2.76 Ma北部类群内部进一步分化 [58 ] Gnaptorina 青藏高原及周边高海拔地区 始新世起源于青藏高原东南缘 [18 -19 ] Coelotinae 北半球温带、亚热带 晚始新世高原东南缘隆升阻碍南方隙蛛向北扩散,渐新世高原中部和北部隆起导致北方隙蛛早期多样化进程中的灭绝事件 [59 ] Montifringilla, Onychostruthus, Pyrgilauda 青藏高原 雪雀的共同祖先在约14 Ma就应经生活在青藏高原,并进化出适应高海拔极端环境的性状 [21 ] Parus major 亚洲、欧洲 蒙古分布类群与其他种类分化时间为2.8~0.7 Ma,喜马拉雅山脉东部与中国中东部类群分化时间为1.9~0.4 Ma [60 ] Pseudopodoces humilis 青藏高原、柴达木盆地周边高山 在9.9~7.7 Ma期间地山雀就与与其他山雀分化开来,并出现适应高海拔性状 [61 ] Phylloscopus, Seicercus 亚洲 喜马拉雅山脉分布类群在7 Ma加速进化 [62 -63 ] Perdix 西藏、甘肃、青海、四川、新疆 高原类群出现于3.63 (4.52~2.75) Ma [64 ] Pantholops hodgsonii 西藏、新疆、青海 上新世-更新世就已经出现在青藏高原 [65 ] Procapra picticaudata 西藏、四川、甘肃、青海、新疆 种内分化时间约2.2~4.4 Ma [66 ] Rhinopithecus 四川、甘肃、陕西、贵州、云南、缅甸、越南 北方类群川金丝猴、黔金丝猴与喜马拉雅类群滇金丝猴、缅甸金丝猴分化时间约1.6 Ma [67 ] Ochotona 北半球 分布于青藏高原的高山类群和灌丛草原类群与低海拔泛北极分布类群分化时间大约为2.4 Ma [68 ] Apodemus draco 四川、云南、甘肃、青海、陕西、湖北、湖南、福建、台湾 2.2 Ma (3.8~0.87 Ma)分化形成,后来由于高原的阶段性隆升以及冰川作用种系及遗传结构更加复杂 [69 -70 ] Eothenomys 东亚 高原种类与低海拔种类分裂时间约2.7 Ma [71 ] Schizothoracinae 青藏高原及周边地区 在晚中新世晚期或上新世裂腹鱼亚科鱼类就已经出现在青藏高原及周边水系中,在晚更新世期间种内分化加速,不同海拔之间的类群产生等级分化,种间多样性增加,第四纪以来青藏高原隆升导致的水系改组是推动裂腹鱼亚科鱼类演化的原动力 [31 ,76 -78 ]
3 讨论 最近,青藏高原古地貌重建工作取得了丰硕的成果(图3 ),但是各种指标重建结果往往存在矛盾(图4 )[25 ,79 -83 ] .同位素重建结果表明高原南部在古近纪早期就存在安第斯型的巨大山系[84 -85 ] ;古生物研究发现高原腹地从中始新世到晚渐新世都存在东西向的大峡谷,该峡谷底部海拔在2 000 m左右,两侧为高大的冈底斯山脉和羌塘山脉[82 ,86 -88 ] ,动植物如巨犀(Paraceratherium )[89 ] 、攀鲈(Eoanabas )[90 -91 ] 、臭椿(Ailanthus )[88 ] 、椿榆(Cedrelospermum )[92 ] 、兔耳果(Lagokarpos )[93 ] 等在古近纪都可以通过青藏地区峡谷与周边动植物区系交流,新近纪之前的青藏地区虽然地形复杂,但是尚未隆升成今天的高原;横断山区某些区域则在晚始新世-早渐新世之交就已经获得现今的海拔高度[94 -95 ] ;高原北部和喜马拉雅山脉则在新近纪形成现在的地貌格局[25 ,96 -97 ] .古生物研究也发现青藏高原在晚上新世就已经存在高寒生境和冰缘环境,成为第四纪冰期动物忍耐寒冷环境的“训练营”,第四纪冰期来临之后这些适应寒冷环境的生物才扩散至北半球高纬度地区[34 ,98 -99 ] .因此,高原及周边地区高寒生物类群的起源时间表明在渐新世青藏高原部分区域就已经出现高寒生态系统,晚中新世以来青藏高原地貌与气候继续发生剧烈变化,促使这些物种的祖先类群迁入或就地演化积累适应高寒环境的基因和形态,并最终演化成适应高寒生境的新物种[25 ] .高寒区物种在晚上新世的加速分化可能与高原巨大山系和水系、冰川活动等造成的地理隔离有关[26 ] .随着第四纪冰期的到来,青藏高原及周边地区的高寒生物区也开始与北半球高纬度地区存在密切联系[34 ,98 -99 ] .同时,演化生物学研究也发现在第四纪冰期期间并未形成覆盖整个青藏高原的大冰盖,高原面上仍存在很多生物避难所[26 ] . ...
Refugial isolation and range expansions drive the genetic structure of Oxyria sinensis (Polygonaceae) in the Himalaya-Hengduan Mountains
2
2015
... 植物中的桃儿七(Sinopodophyllum hexandrum )分布于青藏高原东部,系统发育关系推断其出现在青藏高原的时间约为(6.52±1.89) Ma,但是适应高寒性状出现的时间为晚上新世[47 ] .菊科毛冠菊属(Nannoglottis )植物中生活于高山灌丛和亚高山针叶林的类群形态明显不同,两者的分化时间约为3.4 Ma[48 ] .分布于中国西北干旱区和青藏高原北部及东北部的红砂(Reaumuria soongarica ),在6.3~2.0 Ma期间演化出适应高寒生境的类群,高原北部分布类群也在3.5~1.1 Ma期间响应构造活动造成的地理隔离而发生分化[49 ] .红门兰亚族(Orchidinae)六大分支在多样化的时空序列上表现出明显的差异,其生活在青藏高原地区的特有种大多数是在过去6 Ma里进化而来的[50 ] .中华山蓼(Oxyria sinensis )分布于青藏高原东南部海拔1 600~3 800 m的高山上,约13.8 Ma演化出高山类群,第四纪期间种内多样化增加[51 ] .小子圆柏(Juniperus microsperma )是青藏高原特有种,现在仅生长于西藏波密地区,高原南部在中新世早期构造隆升造成的物种隔离使其与其他刺柏种群发生分化[52 ] .横断山区杜鹃花属(Rhododendron )多样性在晚中新世(约8 Ma)开始快速增加,对青藏高原不同区域杜鹃花属的演化历史进行定量化比较研究,发现横断山区在晚中新世的快速隆升促进生物多样性的形成[53 ] .对喜马拉雅-横断山分布的高山姜类的研究发现,在早渐新世(约32 Ma)象牙参属(Roscoea )和距药姜属(Cautleya )就与其姐妹类群分化开来,而象牙参属在横断山区与喜马拉雅山脉的两个分支大概在渐新世-中新世之交分化[54 ] . ...
... Biological evolution in the alpine region of Tibetan Plateau
Table 1 种属 分布区域 适应高海拔类群分化时间或多样性增加时间 文献来源 Sinopodophyllum hexandrum 青藏高原东部 在约(6.52±1.89) Ma期间出现在青藏高原东部,但是直到3~2 Ma以来才演化出适应高山环境的形态 [47 ] Nannoglottis 青藏高原 在23~32 Ma期间出现在青藏高原,随后多样性增高,其高山灌丛和针叶林生活类型分支时间约为3.4 Ma [48 ] Reaumuria soongarica 西北干旱区、青藏高原北部 高原内部分布类群演化时间2.0~6.3 Ma,高原东北缘与高原西北类群分化时间1.1~3.5 Ma [49 ] Orchidinae 青藏高原及喜马拉雅山脉 青藏高原特有种大多是在过去6 Ma里进化而来 [50 ] Oxyria sinensis 青藏高原东南部 高山类群出现时间7.54~24.36 Ma,种内多样化增加时间1.74~0.86 Ma [51 ] Juniperus microsperma 西藏波密 早中新世21.6~14 Ma期间与其他种群分化 [52 ] Rhododendron 北半球 横断山区概属多样性在晚中新世(约8 Ma)快速增加 [53 ] Roscoea, Cautleya 喜马拉雅-横断山区 在早渐新世(~32 Ma)两属出现,Roscoea在横断山区与喜马拉雅山脉的两个分支大概在渐新世-中新世之交分化 [54 ] Nanorana parkeri 青藏高原南部 高原东部和西部类群分化时间1.4~3.7 Ma [55 ] Paini 南亚、东南亚、青藏高原 在渐新世早期棘蛙族高原-喜马拉雅类群与中国第二阶梯上生活的类群分化,随后在在26.1~12.5 Ma高原生活类群与喜马拉雅山脉生活类群分化,高原内部生活类群在晚中新世到上新世期间分化速率加快 [22 ] Bufo tibetanus 青藏高原、四川、甘肃、陕西等 高原种类形成于上新世期间 [56 ] Phrynocephalus 亚洲干旱区 在7.2~13.0 Ma期间演化出胎生类群,3.2~7.3 Ma期间种间多样性增加 [57 ] Phrynocephalus erythrurus 羌塘盆地 在3.67 Ma左右北部类群与南部类群发生分化,约2.76 Ma北部类群内部进一步分化 [58 ] Gnaptorina 青藏高原及周边高海拔地区 始新世起源于青藏高原东南缘 [18 -19 ] Coelotinae 北半球温带、亚热带 晚始新世高原东南缘隆升阻碍南方隙蛛向北扩散,渐新世高原中部和北部隆起导致北方隙蛛早期多样化进程中的灭绝事件 [59 ] Montifringilla, Onychostruthus, Pyrgilauda 青藏高原 雪雀的共同祖先在约14 Ma就应经生活在青藏高原,并进化出适应高海拔极端环境的性状 [21 ] Parus major 亚洲、欧洲 蒙古分布类群与其他种类分化时间为2.8~0.7 Ma,喜马拉雅山脉东部与中国中东部类群分化时间为1.9~0.4 Ma [60 ] Pseudopodoces humilis 青藏高原、柴达木盆地周边高山 在9.9~7.7 Ma期间地山雀就与与其他山雀分化开来,并出现适应高海拔性状 [61 ] Phylloscopus, Seicercus 亚洲 喜马拉雅山脉分布类群在7 Ma加速进化 [62 -63 ] Perdix 西藏、甘肃、青海、四川、新疆 高原类群出现于3.63 (4.52~2.75) Ma [64 ] Pantholops hodgsonii 西藏、新疆、青海 上新世-更新世就已经出现在青藏高原 [65 ] Procapra picticaudata 西藏、四川、甘肃、青海、新疆 种内分化时间约2.2~4.4 Ma [66 ] Rhinopithecus 四川、甘肃、陕西、贵州、云南、缅甸、越南 北方类群川金丝猴、黔金丝猴与喜马拉雅类群滇金丝猴、缅甸金丝猴分化时间约1.6 Ma [67 ] Ochotona 北半球 分布于青藏高原的高山类群和灌丛草原类群与低海拔泛北极分布类群分化时间大约为2.4 Ma [68 ] Apodemus draco 四川、云南、甘肃、青海、陕西、湖北、湖南、福建、台湾 2.2 Ma (3.8~0.87 Ma)分化形成,后来由于高原的阶段性隆升以及冰川作用种系及遗传结构更加复杂 [69 -70 ] Eothenomys 东亚 高原种类与低海拔种类分裂时间约2.7 Ma [71 ] Schizothoracinae 青藏高原及周边地区 在晚中新世晚期或上新世裂腹鱼亚科鱼类就已经出现在青藏高原及周边水系中,在晚更新世期间种内分化加速,不同海拔之间的类群产生等级分化,种间多样性增加,第四纪以来青藏高原隆升导致的水系改组是推动裂腹鱼亚科鱼类演化的原动力 [31 ,76 -78 ]
3 讨论 最近,青藏高原古地貌重建工作取得了丰硕的成果(图3 ),但是各种指标重建结果往往存在矛盾(图4 )[25 ,79 -83 ] .同位素重建结果表明高原南部在古近纪早期就存在安第斯型的巨大山系[84 -85 ] ;古生物研究发现高原腹地从中始新世到晚渐新世都存在东西向的大峡谷,该峡谷底部海拔在2 000 m左右,两侧为高大的冈底斯山脉和羌塘山脉[82 ,86 -88 ] ,动植物如巨犀(Paraceratherium )[89 ] 、攀鲈(Eoanabas )[90 -91 ] 、臭椿(Ailanthus )[88 ] 、椿榆(Cedrelospermum )[92 ] 、兔耳果(Lagokarpos )[93 ] 等在古近纪都可以通过青藏地区峡谷与周边动植物区系交流,新近纪之前的青藏地区虽然地形复杂,但是尚未隆升成今天的高原;横断山区某些区域则在晚始新世-早渐新世之交就已经获得现今的海拔高度[94 -95 ] ;高原北部和喜马拉雅山脉则在新近纪形成现在的地貌格局[25 ,96 -97 ] .古生物研究也发现青藏高原在晚上新世就已经存在高寒生境和冰缘环境,成为第四纪冰期动物忍耐寒冷环境的“训练营”,第四纪冰期来临之后这些适应寒冷环境的生物才扩散至北半球高纬度地区[34 ,98 -99 ] .因此,高原及周边地区高寒生物类群的起源时间表明在渐新世青藏高原部分区域就已经出现高寒生态系统,晚中新世以来青藏高原地貌与气候继续发生剧烈变化,促使这些物种的祖先类群迁入或就地演化积累适应高寒环境的基因和形态,并最终演化成适应高寒生境的新物种[25 ] .高寒区物种在晚上新世的加速分化可能与高原巨大山系和水系、冰川活动等造成的地理隔离有关[26 ] .随着第四纪冰期的到来,青藏高原及周边地区的高寒生物区也开始与北半球高纬度地区存在密切联系[34 ,98 -99 ] .同时,演化生物学研究也发现在第四纪冰期期间并未形成覆盖整个青藏高原的大冰盖,高原面上仍存在很多生物避难所[26 ] . ...
Evolutionary origin and demographic history of an ancient conifer (Juniperus microsperma ) in the Qinghai-Tibetan Plateau
2
2015
... 植物中的桃儿七(Sinopodophyllum hexandrum )分布于青藏高原东部,系统发育关系推断其出现在青藏高原的时间约为(6.52±1.89) Ma,但是适应高寒性状出现的时间为晚上新世[47 ] .菊科毛冠菊属(Nannoglottis )植物中生活于高山灌丛和亚高山针叶林的类群形态明显不同,两者的分化时间约为3.4 Ma[48 ] .分布于中国西北干旱区和青藏高原北部及东北部的红砂(Reaumuria soongarica ),在6.3~2.0 Ma期间演化出适应高寒生境的类群,高原北部分布类群也在3.5~1.1 Ma期间响应构造活动造成的地理隔离而发生分化[49 ] .红门兰亚族(Orchidinae)六大分支在多样化的时空序列上表现出明显的差异,其生活在青藏高原地区的特有种大多数是在过去6 Ma里进化而来的[50 ] .中华山蓼(Oxyria sinensis )分布于青藏高原东南部海拔1 600~3 800 m的高山上,约13.8 Ma演化出高山类群,第四纪期间种内多样化增加[51 ] .小子圆柏(Juniperus microsperma )是青藏高原特有种,现在仅生长于西藏波密地区,高原南部在中新世早期构造隆升造成的物种隔离使其与其他刺柏种群发生分化[52 ] .横断山区杜鹃花属(Rhododendron )多样性在晚中新世(约8 Ma)开始快速增加,对青藏高原不同区域杜鹃花属的演化历史进行定量化比较研究,发现横断山区在晚中新世的快速隆升促进生物多样性的形成[53 ] .对喜马拉雅-横断山分布的高山姜类的研究发现,在早渐新世(约32 Ma)象牙参属(Roscoea )和距药姜属(Cautleya )就与其姐妹类群分化开来,而象牙参属在横断山区与喜马拉雅山脉的两个分支大概在渐新世-中新世之交分化[54 ] . ...
... Biological evolution in the alpine region of Tibetan Plateau
Table 1 种属 分布区域 适应高海拔类群分化时间或多样性增加时间 文献来源 Sinopodophyllum hexandrum 青藏高原东部 在约(6.52±1.89) Ma期间出现在青藏高原东部,但是直到3~2 Ma以来才演化出适应高山环境的形态 [47 ] Nannoglottis 青藏高原 在23~32 Ma期间出现在青藏高原,随后多样性增高,其高山灌丛和针叶林生活类型分支时间约为3.4 Ma [48 ] Reaumuria soongarica 西北干旱区、青藏高原北部 高原内部分布类群演化时间2.0~6.3 Ma,高原东北缘与高原西北类群分化时间1.1~3.5 Ma [49 ] Orchidinae 青藏高原及喜马拉雅山脉 青藏高原特有种大多是在过去6 Ma里进化而来 [50 ] Oxyria sinensis 青藏高原东南部 高山类群出现时间7.54~24.36 Ma,种内多样化增加时间1.74~0.86 Ma [51 ] Juniperus microsperma 西藏波密 早中新世21.6~14 Ma期间与其他种群分化 [52 ] Rhododendron 北半球 横断山区概属多样性在晚中新世(约8 Ma)快速增加 [53 ] Roscoea, Cautleya 喜马拉雅-横断山区 在早渐新世(~32 Ma)两属出现,Roscoea在横断山区与喜马拉雅山脉的两个分支大概在渐新世-中新世之交分化 [54 ] Nanorana parkeri 青藏高原南部 高原东部和西部类群分化时间1.4~3.7 Ma [55 ] Paini 南亚、东南亚、青藏高原 在渐新世早期棘蛙族高原-喜马拉雅类群与中国第二阶梯上生活的类群分化,随后在在26.1~12.5 Ma高原生活类群与喜马拉雅山脉生活类群分化,高原内部生活类群在晚中新世到上新世期间分化速率加快 [22 ] Bufo tibetanus 青藏高原、四川、甘肃、陕西等 高原种类形成于上新世期间 [56 ] Phrynocephalus 亚洲干旱区 在7.2~13.0 Ma期间演化出胎生类群,3.2~7.3 Ma期间种间多样性增加 [57 ] Phrynocephalus erythrurus 羌塘盆地 在3.67 Ma左右北部类群与南部类群发生分化,约2.76 Ma北部类群内部进一步分化 [58 ] Gnaptorina 青藏高原及周边高海拔地区 始新世起源于青藏高原东南缘 [18 -19 ] Coelotinae 北半球温带、亚热带 晚始新世高原东南缘隆升阻碍南方隙蛛向北扩散,渐新世高原中部和北部隆起导致北方隙蛛早期多样化进程中的灭绝事件 [59 ] Montifringilla, Onychostruthus, Pyrgilauda 青藏高原 雪雀的共同祖先在约14 Ma就应经生活在青藏高原,并进化出适应高海拔极端环境的性状 [21 ] Parus major 亚洲、欧洲 蒙古分布类群与其他种类分化时间为2.8~0.7 Ma,喜马拉雅山脉东部与中国中东部类群分化时间为1.9~0.4 Ma [60 ] Pseudopodoces humilis 青藏高原、柴达木盆地周边高山 在9.9~7.7 Ma期间地山雀就与与其他山雀分化开来,并出现适应高海拔性状 [61 ] Phylloscopus, Seicercus 亚洲 喜马拉雅山脉分布类群在7 Ma加速进化 [62 -63 ] Perdix 西藏、甘肃、青海、四川、新疆 高原类群出现于3.63 (4.52~2.75) Ma [64 ] Pantholops hodgsonii 西藏、新疆、青海 上新世-更新世就已经出现在青藏高原 [65 ] Procapra picticaudata 西藏、四川、甘肃、青海、新疆 种内分化时间约2.2~4.4 Ma [66 ] Rhinopithecus 四川、甘肃、陕西、贵州、云南、缅甸、越南 北方类群川金丝猴、黔金丝猴与喜马拉雅类群滇金丝猴、缅甸金丝猴分化时间约1.6 Ma [67 ] Ochotona 北半球 分布于青藏高原的高山类群和灌丛草原类群与低海拔泛北极分布类群分化时间大约为2.4 Ma [68 ] Apodemus draco 四川、云南、甘肃、青海、陕西、湖北、湖南、福建、台湾 2.2 Ma (3.8~0.87 Ma)分化形成,后来由于高原的阶段性隆升以及冰川作用种系及遗传结构更加复杂 [69 -70 ] Eothenomys 东亚 高原种类与低海拔种类分裂时间约2.7 Ma [71 ] Schizothoracinae 青藏高原及周边地区 在晚中新世晚期或上新世裂腹鱼亚科鱼类就已经出现在青藏高原及周边水系中,在晚更新世期间种内分化加速,不同海拔之间的类群产生等级分化,种间多样性增加,第四纪以来青藏高原隆升导致的水系改组是推动裂腹鱼亚科鱼类演化的原动力 [31 ,76 -78 ]
3 讨论 最近,青藏高原古地貌重建工作取得了丰硕的成果(图3 ),但是各种指标重建结果往往存在矛盾(图4 )[25 ,79 -83 ] .同位素重建结果表明高原南部在古近纪早期就存在安第斯型的巨大山系[84 -85 ] ;古生物研究发现高原腹地从中始新世到晚渐新世都存在东西向的大峡谷,该峡谷底部海拔在2 000 m左右,两侧为高大的冈底斯山脉和羌塘山脉[82 ,86 -88 ] ,动植物如巨犀(Paraceratherium )[89 ] 、攀鲈(Eoanabas )[90 -91 ] 、臭椿(Ailanthus )[88 ] 、椿榆(Cedrelospermum )[92 ] 、兔耳果(Lagokarpos )[93 ] 等在古近纪都可以通过青藏地区峡谷与周边动植物区系交流,新近纪之前的青藏地区虽然地形复杂,但是尚未隆升成今天的高原;横断山区某些区域则在晚始新世-早渐新世之交就已经获得现今的海拔高度[94 -95 ] ;高原北部和喜马拉雅山脉则在新近纪形成现在的地貌格局[25 ,96 -97 ] .古生物研究也发现青藏高原在晚上新世就已经存在高寒生境和冰缘环境,成为第四纪冰期动物忍耐寒冷环境的“训练营”,第四纪冰期来临之后这些适应寒冷环境的生物才扩散至北半球高纬度地区[34 ,98 -99 ] .因此,高原及周边地区高寒生物类群的起源时间表明在渐新世青藏高原部分区域就已经出现高寒生态系统,晚中新世以来青藏高原地貌与气候继续发生剧烈变化,促使这些物种的祖先类群迁入或就地演化积累适应高寒环境的基因和形态,并最终演化成适应高寒生境的新物种[25 ] .高寒区物种在晚上新世的加速分化可能与高原巨大山系和水系、冰川活动等造成的地理隔离有关[26 ] .随着第四纪冰期的到来,青藏高原及周边地区的高寒生物区也开始与北半球高纬度地区存在密切联系[34 ,98 -99 ] .同时,演化生物学研究也发现在第四纪冰期期间并未形成覆盖整个青藏高原的大冰盖,高原面上仍存在很多生物避难所[26 ] . ...
Uplift-driven diversification in the Hengduan Mountains, a temperate biodiversity hotspot
3
2017
... 植物中的桃儿七(Sinopodophyllum hexandrum )分布于青藏高原东部,系统发育关系推断其出现在青藏高原的时间约为(6.52±1.89) Ma,但是适应高寒性状出现的时间为晚上新世[47 ] .菊科毛冠菊属(Nannoglottis )植物中生活于高山灌丛和亚高山针叶林的类群形态明显不同,两者的分化时间约为3.4 Ma[48 ] .分布于中国西北干旱区和青藏高原北部及东北部的红砂(Reaumuria soongarica ),在6.3~2.0 Ma期间演化出适应高寒生境的类群,高原北部分布类群也在3.5~1.1 Ma期间响应构造活动造成的地理隔离而发生分化[49 ] .红门兰亚族(Orchidinae)六大分支在多样化的时空序列上表现出明显的差异,其生活在青藏高原地区的特有种大多数是在过去6 Ma里进化而来的[50 ] .中华山蓼(Oxyria sinensis )分布于青藏高原东南部海拔1 600~3 800 m的高山上,约13.8 Ma演化出高山类群,第四纪期间种内多样化增加[51 ] .小子圆柏(Juniperus microsperma )是青藏高原特有种,现在仅生长于西藏波密地区,高原南部在中新世早期构造隆升造成的物种隔离使其与其他刺柏种群发生分化[52 ] .横断山区杜鹃花属(Rhododendron )多样性在晚中新世(约8 Ma)开始快速增加,对青藏高原不同区域杜鹃花属的演化历史进行定量化比较研究,发现横断山区在晚中新世的快速隆升促进生物多样性的形成[53 ] .对喜马拉雅-横断山分布的高山姜类的研究发现,在早渐新世(约32 Ma)象牙参属(Roscoea )和距药姜属(Cautleya )就与其姐妹类群分化开来,而象牙参属在横断山区与喜马拉雅山脉的两个分支大概在渐新世-中新世之交分化[54 ] . ...
... Biological evolution in the alpine region of Tibetan Plateau
Table 1 种属 分布区域 适应高海拔类群分化时间或多样性增加时间 文献来源 Sinopodophyllum hexandrum 青藏高原东部 在约(6.52±1.89) Ma期间出现在青藏高原东部,但是直到3~2 Ma以来才演化出适应高山环境的形态 [47 ] Nannoglottis 青藏高原 在23~32 Ma期间出现在青藏高原,随后多样性增高,其高山灌丛和针叶林生活类型分支时间约为3.4 Ma [48 ] Reaumuria soongarica 西北干旱区、青藏高原北部 高原内部分布类群演化时间2.0~6.3 Ma,高原东北缘与高原西北类群分化时间1.1~3.5 Ma [49 ] Orchidinae 青藏高原及喜马拉雅山脉 青藏高原特有种大多是在过去6 Ma里进化而来 [50 ] Oxyria sinensis 青藏高原东南部 高山类群出现时间7.54~24.36 Ma,种内多样化增加时间1.74~0.86 Ma [51 ] Juniperus microsperma 西藏波密 早中新世21.6~14 Ma期间与其他种群分化 [52 ] Rhododendron 北半球 横断山区概属多样性在晚中新世(约8 Ma)快速增加 [53 ] Roscoea, Cautleya 喜马拉雅-横断山区 在早渐新世(~32 Ma)两属出现,Roscoea在横断山区与喜马拉雅山脉的两个分支大概在渐新世-中新世之交分化 [54 ] Nanorana parkeri 青藏高原南部 高原东部和西部类群分化时间1.4~3.7 Ma [55 ] Paini 南亚、东南亚、青藏高原 在渐新世早期棘蛙族高原-喜马拉雅类群与中国第二阶梯上生活的类群分化,随后在在26.1~12.5 Ma高原生活类群与喜马拉雅山脉生活类群分化,高原内部生活类群在晚中新世到上新世期间分化速率加快 [22 ] Bufo tibetanus 青藏高原、四川、甘肃、陕西等 高原种类形成于上新世期间 [56 ] Phrynocephalus 亚洲干旱区 在7.2~13.0 Ma期间演化出胎生类群,3.2~7.3 Ma期间种间多样性增加 [57 ] Phrynocephalus erythrurus 羌塘盆地 在3.67 Ma左右北部类群与南部类群发生分化,约2.76 Ma北部类群内部进一步分化 [58 ] Gnaptorina 青藏高原及周边高海拔地区 始新世起源于青藏高原东南缘 [18 -19 ] Coelotinae 北半球温带、亚热带 晚始新世高原东南缘隆升阻碍南方隙蛛向北扩散,渐新世高原中部和北部隆起导致北方隙蛛早期多样化进程中的灭绝事件 [59 ] Montifringilla, Onychostruthus, Pyrgilauda 青藏高原 雪雀的共同祖先在约14 Ma就应经生活在青藏高原,并进化出适应高海拔极端环境的性状 [21 ] Parus major 亚洲、欧洲 蒙古分布类群与其他种类分化时间为2.8~0.7 Ma,喜马拉雅山脉东部与中国中东部类群分化时间为1.9~0.4 Ma [60 ] Pseudopodoces humilis 青藏高原、柴达木盆地周边高山 在9.9~7.7 Ma期间地山雀就与与其他山雀分化开来,并出现适应高海拔性状 [61 ] Phylloscopus, Seicercus 亚洲 喜马拉雅山脉分布类群在7 Ma加速进化 [62 -63 ] Perdix 西藏、甘肃、青海、四川、新疆 高原类群出现于3.63 (4.52~2.75) Ma [64 ] Pantholops hodgsonii 西藏、新疆、青海 上新世-更新世就已经出现在青藏高原 [65 ] Procapra picticaudata 西藏、四川、甘肃、青海、新疆 种内分化时间约2.2~4.4 Ma [66 ] Rhinopithecus 四川、甘肃、陕西、贵州、云南、缅甸、越南 北方类群川金丝猴、黔金丝猴与喜马拉雅类群滇金丝猴、缅甸金丝猴分化时间约1.6 Ma [67 ] Ochotona 北半球 分布于青藏高原的高山类群和灌丛草原类群与低海拔泛北极分布类群分化时间大约为2.4 Ma [68 ] Apodemus draco 四川、云南、甘肃、青海、陕西、湖北、湖南、福建、台湾 2.2 Ma (3.8~0.87 Ma)分化形成,后来由于高原的阶段性隆升以及冰川作用种系及遗传结构更加复杂 [69 -70 ] Eothenomys 东亚 高原种类与低海拔种类分裂时间约2.7 Ma [71 ] Schizothoracinae 青藏高原及周边地区 在晚中新世晚期或上新世裂腹鱼亚科鱼类就已经出现在青藏高原及周边水系中,在晚更新世期间种内分化加速,不同海拔之间的类群产生等级分化,种间多样性增加,第四纪以来青藏高原隆升导致的水系改组是推动裂腹鱼亚科鱼类演化的原动力 [31 ,76 -78 ]
3 讨论 最近,青藏高原古地貌重建工作取得了丰硕的成果(图3 ),但是各种指标重建结果往往存在矛盾(图4 )[25 ,79 -83 ] .同位素重建结果表明高原南部在古近纪早期就存在安第斯型的巨大山系[84 -85 ] ;古生物研究发现高原腹地从中始新世到晚渐新世都存在东西向的大峡谷,该峡谷底部海拔在2 000 m左右,两侧为高大的冈底斯山脉和羌塘山脉[82 ,86 -88 ] ,动植物如巨犀(Paraceratherium )[89 ] 、攀鲈(Eoanabas )[90 -91 ] 、臭椿(Ailanthus )[88 ] 、椿榆(Cedrelospermum )[92 ] 、兔耳果(Lagokarpos )[93 ] 等在古近纪都可以通过青藏地区峡谷与周边动植物区系交流,新近纪之前的青藏地区虽然地形复杂,但是尚未隆升成今天的高原;横断山区某些区域则在晚始新世-早渐新世之交就已经获得现今的海拔高度[94 -95 ] ;高原北部和喜马拉雅山脉则在新近纪形成现在的地貌格局[25 ,96 -97 ] .古生物研究也发现青藏高原在晚上新世就已经存在高寒生境和冰缘环境,成为第四纪冰期动物忍耐寒冷环境的“训练营”,第四纪冰期来临之后这些适应寒冷环境的生物才扩散至北半球高纬度地区[34 ,98 -99 ] .因此,高原及周边地区高寒生物类群的起源时间表明在渐新世青藏高原部分区域就已经出现高寒生态系统,晚中新世以来青藏高原地貌与气候继续发生剧烈变化,促使这些物种的祖先类群迁入或就地演化积累适应高寒环境的基因和形态,并最终演化成适应高寒生境的新物种[25 ] .高寒区物种在晚上新世的加速分化可能与高原巨大山系和水系、冰川活动等造成的地理隔离有关[26 ] .随着第四纪冰期的到来,青藏高原及周边地区的高寒生物区也开始与北半球高纬度地区存在密切联系[34 ,98 -99 ] .同时,演化生物学研究也发现在第四纪冰期期间并未形成覆盖整个青藏高原的大冰盖,高原面上仍存在很多生物避难所[26 ] . ...
... 随着DNA测序技术和比较系统学方法的发展,大量涌现的青藏高寒区生物地理学研究为我们理解高原隆升过程及其气候效应提供了生物演化方面的证据[25 ] .但是,演化生物学家通常仅对单一特定类群的系统发育过程开展研究,很少进行系统的总结工作[25 ] ,对某一类群遗传数据的解释也往往与最新的青藏高原隆升和气候变化研究进展脱节,不能很好地将最新的关于高原隆升和气候变化的研究与生物类群演化联系在一起[101 ] ,同时也没有认识到青藏高原的隆升过程是阶段性的,没有对青藏高原的不同组成部分进行区分[26 ,53 ,81 ] .地质学研究大多认为高原地貌格局形成于中新世之前,但是综合近年来的生物地理学研究结果可以发现,青藏高寒生物类群的分化时间多集中在晚中新世,种内分化加速和多样性增加主要发生在晚上新世到第四纪期间(图5 ),这些不同类群在分化时间上的巧合可能反映青藏高寒区生物演化受到晚中新世以来构造和气候事件的影响,但是并未引起地质学家足够的重视[25 ] . ...
Evolutionary diversification of alpine ginger reflects the early uplift of the Himalayan-Tibetan Plateau and rapid extrusion of Indochina
2
2016
... 植物中的桃儿七(Sinopodophyllum hexandrum )分布于青藏高原东部,系统发育关系推断其出现在青藏高原的时间约为(6.52±1.89) Ma,但是适应高寒性状出现的时间为晚上新世[47 ] .菊科毛冠菊属(Nannoglottis )植物中生活于高山灌丛和亚高山针叶林的类群形态明显不同,两者的分化时间约为3.4 Ma[48 ] .分布于中国西北干旱区和青藏高原北部及东北部的红砂(Reaumuria soongarica ),在6.3~2.0 Ma期间演化出适应高寒生境的类群,高原北部分布类群也在3.5~1.1 Ma期间响应构造活动造成的地理隔离而发生分化[49 ] .红门兰亚族(Orchidinae)六大分支在多样化的时空序列上表现出明显的差异,其生活在青藏高原地区的特有种大多数是在过去6 Ma里进化而来的[50 ] .中华山蓼(Oxyria sinensis )分布于青藏高原东南部海拔1 600~3 800 m的高山上,约13.8 Ma演化出高山类群,第四纪期间种内多样化增加[51 ] .小子圆柏(Juniperus microsperma )是青藏高原特有种,现在仅生长于西藏波密地区,高原南部在中新世早期构造隆升造成的物种隔离使其与其他刺柏种群发生分化[52 ] .横断山区杜鹃花属(Rhododendron )多样性在晚中新世(约8 Ma)开始快速增加,对青藏高原不同区域杜鹃花属的演化历史进行定量化比较研究,发现横断山区在晚中新世的快速隆升促进生物多样性的形成[53 ] .对喜马拉雅-横断山分布的高山姜类的研究发现,在早渐新世(约32 Ma)象牙参属(Roscoea )和距药姜属(Cautleya )就与其姐妹类群分化开来,而象牙参属在横断山区与喜马拉雅山脉的两个分支大概在渐新世-中新世之交分化[54 ] . ...
... Biological evolution in the alpine region of Tibetan Plateau
Table 1 种属 分布区域 适应高海拔类群分化时间或多样性增加时间 文献来源 Sinopodophyllum hexandrum 青藏高原东部 在约(6.52±1.89) Ma期间出现在青藏高原东部,但是直到3~2 Ma以来才演化出适应高山环境的形态 [47 ] Nannoglottis 青藏高原 在23~32 Ma期间出现在青藏高原,随后多样性增高,其高山灌丛和针叶林生活类型分支时间约为3.4 Ma [48 ] Reaumuria soongarica 西北干旱区、青藏高原北部 高原内部分布类群演化时间2.0~6.3 Ma,高原东北缘与高原西北类群分化时间1.1~3.5 Ma [49 ] Orchidinae 青藏高原及喜马拉雅山脉 青藏高原特有种大多是在过去6 Ma里进化而来 [50 ] Oxyria sinensis 青藏高原东南部 高山类群出现时间7.54~24.36 Ma,种内多样化增加时间1.74~0.86 Ma [51 ] Juniperus microsperma 西藏波密 早中新世21.6~14 Ma期间与其他种群分化 [52 ] Rhododendron 北半球 横断山区概属多样性在晚中新世(约8 Ma)快速增加 [53 ] Roscoea, Cautleya 喜马拉雅-横断山区 在早渐新世(~32 Ma)两属出现,Roscoea在横断山区与喜马拉雅山脉的两个分支大概在渐新世-中新世之交分化 [54 ] Nanorana parkeri 青藏高原南部 高原东部和西部类群分化时间1.4~3.7 Ma [55 ] Paini 南亚、东南亚、青藏高原 在渐新世早期棘蛙族高原-喜马拉雅类群与中国第二阶梯上生活的类群分化,随后在在26.1~12.5 Ma高原生活类群与喜马拉雅山脉生活类群分化,高原内部生活类群在晚中新世到上新世期间分化速率加快 [22 ] Bufo tibetanus 青藏高原、四川、甘肃、陕西等 高原种类形成于上新世期间 [56 ] Phrynocephalus 亚洲干旱区 在7.2~13.0 Ma期间演化出胎生类群,3.2~7.3 Ma期间种间多样性增加 [57 ] Phrynocephalus erythrurus 羌塘盆地 在3.67 Ma左右北部类群与南部类群发生分化,约2.76 Ma北部类群内部进一步分化 [58 ] Gnaptorina 青藏高原及周边高海拔地区 始新世起源于青藏高原东南缘 [18 -19 ] Coelotinae 北半球温带、亚热带 晚始新世高原东南缘隆升阻碍南方隙蛛向北扩散,渐新世高原中部和北部隆起导致北方隙蛛早期多样化进程中的灭绝事件 [59 ] Montifringilla, Onychostruthus, Pyrgilauda 青藏高原 雪雀的共同祖先在约14 Ma就应经生活在青藏高原,并进化出适应高海拔极端环境的性状 [21 ] Parus major 亚洲、欧洲 蒙古分布类群与其他种类分化时间为2.8~0.7 Ma,喜马拉雅山脉东部与中国中东部类群分化时间为1.9~0.4 Ma [60 ] Pseudopodoces humilis 青藏高原、柴达木盆地周边高山 在9.9~7.7 Ma期间地山雀就与与其他山雀分化开来,并出现适应高海拔性状 [61 ] Phylloscopus, Seicercus 亚洲 喜马拉雅山脉分布类群在7 Ma加速进化 [62 -63 ] Perdix 西藏、甘肃、青海、四川、新疆 高原类群出现于3.63 (4.52~2.75) Ma [64 ] Pantholops hodgsonii 西藏、新疆、青海 上新世-更新世就已经出现在青藏高原 [65 ] Procapra picticaudata 西藏、四川、甘肃、青海、新疆 种内分化时间约2.2~4.4 Ma [66 ] Rhinopithecus 四川、甘肃、陕西、贵州、云南、缅甸、越南 北方类群川金丝猴、黔金丝猴与喜马拉雅类群滇金丝猴、缅甸金丝猴分化时间约1.6 Ma [67 ] Ochotona 北半球 分布于青藏高原的高山类群和灌丛草原类群与低海拔泛北极分布类群分化时间大约为2.4 Ma [68 ] Apodemus draco 四川、云南、甘肃、青海、陕西、湖北、湖南、福建、台湾 2.2 Ma (3.8~0.87 Ma)分化形成,后来由于高原的阶段性隆升以及冰川作用种系及遗传结构更加复杂 [69 -70 ] Eothenomys 东亚 高原种类与低海拔种类分裂时间约2.7 Ma [71 ] Schizothoracinae 青藏高原及周边地区 在晚中新世晚期或上新世裂腹鱼亚科鱼类就已经出现在青藏高原及周边水系中,在晚更新世期间种内分化加速,不同海拔之间的类群产生等级分化,种间多样性增加,第四纪以来青藏高原隆升导致的水系改组是推动裂腹鱼亚科鱼类演化的原动力 [31 ,76 -78 ]
3 讨论 最近,青藏高原古地貌重建工作取得了丰硕的成果(图3 ),但是各种指标重建结果往往存在矛盾(图4 )[25 ,79 -83 ] .同位素重建结果表明高原南部在古近纪早期就存在安第斯型的巨大山系[84 -85 ] ;古生物研究发现高原腹地从中始新世到晚渐新世都存在东西向的大峡谷,该峡谷底部海拔在2 000 m左右,两侧为高大的冈底斯山脉和羌塘山脉[82 ,86 -88 ] ,动植物如巨犀(Paraceratherium )[89 ] 、攀鲈(Eoanabas )[90 -91 ] 、臭椿(Ailanthus )[88 ] 、椿榆(Cedrelospermum )[92 ] 、兔耳果(Lagokarpos )[93 ] 等在古近纪都可以通过青藏地区峡谷与周边动植物区系交流,新近纪之前的青藏地区虽然地形复杂,但是尚未隆升成今天的高原;横断山区某些区域则在晚始新世-早渐新世之交就已经获得现今的海拔高度[94 -95 ] ;高原北部和喜马拉雅山脉则在新近纪形成现在的地貌格局[25 ,96 -97 ] .古生物研究也发现青藏高原在晚上新世就已经存在高寒生境和冰缘环境,成为第四纪冰期动物忍耐寒冷环境的“训练营”,第四纪冰期来临之后这些适应寒冷环境的生物才扩散至北半球高纬度地区[34 ,98 -99 ] .因此,高原及周边地区高寒生物类群的起源时间表明在渐新世青藏高原部分区域就已经出现高寒生态系统,晚中新世以来青藏高原地貌与气候继续发生剧烈变化,促使这些物种的祖先类群迁入或就地演化积累适应高寒环境的基因和形态,并最终演化成适应高寒生境的新物种[25 ] .高寒区物种在晚上新世的加速分化可能与高原巨大山系和水系、冰川活动等造成的地理隔离有关[26 ] .随着第四纪冰期的到来,青藏高原及周边地区的高寒生物区也开始与北半球高纬度地区存在密切联系[34 ,98 -99 ] .同时,演化生物学研究也发现在第四纪冰期期间并未形成覆盖整个青藏高原的大冰盖,高原面上仍存在很多生物避难所[26 ] . ...
Phylogeography of Nanorana parkeri (Anura: Ranidae) and multiple refugia on the Tibetan Plateau revealed by mitochondrial and nuclear DNA
2
2015
... 两栖动物中的高山倭蛙(Nanorana parkeri )分布于青藏高原南部海拔2 850~4 700 m之间的高山地区,其高原东部和西部类群分化时间约为3.7~1.4 Ma[55 ] .棘蛙族(Paini)物种多分布于深山急流中,结合其分子进化历史和生物地理分析发现在渐新世早期棘蛙族高原-喜马拉雅类群与中国第二阶梯上生活的类群分化,随后在26.1~12.5 Ma高原生活类群与喜马拉雅山脉生活类群分化,高原内部生活类群在晚中新世到上新世期间分化速率加快[22 ] .生活于青藏高原南部海拔2 850~4 700 m区域的西藏蟾蜍(Bufo tibetanus ),系统发育学研究表明其在上新世就演化出适应高原生活的性状[56 ] .爬行类动物沙蜥属(Phrynocephalus )一般分布于海拔超过2 200 m地区,为适应高原恶劣环境在约13.0~7.2 Ma进化出胎生类群,在7.3~3.2 Ma期间沙蜥属种间多样性快速增加[57 ] .分布于羌塘高原海拔4 500~5 000 m区域的红尾沙蜥(P.erythrurus )在3.67 Ma左右北部类群与南部类群发生分化,约2.76 Ma北部类群内部进一步分化,可能与晚上新世以来唐古拉山脉、乌兰乌拉山脉以及北麓河构造活动形成地理隔离有关[58 ] . ...
... Biological evolution in the alpine region of Tibetan Plateau
Table 1 种属 分布区域 适应高海拔类群分化时间或多样性增加时间 文献来源 Sinopodophyllum hexandrum 青藏高原东部 在约(6.52±1.89) Ma期间出现在青藏高原东部,但是直到3~2 Ma以来才演化出适应高山环境的形态 [47 ] Nannoglottis 青藏高原 在23~32 Ma期间出现在青藏高原,随后多样性增高,其高山灌丛和针叶林生活类型分支时间约为3.4 Ma [48 ] Reaumuria soongarica 西北干旱区、青藏高原北部 高原内部分布类群演化时间2.0~6.3 Ma,高原东北缘与高原西北类群分化时间1.1~3.5 Ma [49 ] Orchidinae 青藏高原及喜马拉雅山脉 青藏高原特有种大多是在过去6 Ma里进化而来 [50 ] Oxyria sinensis 青藏高原东南部 高山类群出现时间7.54~24.36 Ma,种内多样化增加时间1.74~0.86 Ma [51 ] Juniperus microsperma 西藏波密 早中新世21.6~14 Ma期间与其他种群分化 [52 ] Rhododendron 北半球 横断山区概属多样性在晚中新世(约8 Ma)快速增加 [53 ] Roscoea, Cautleya 喜马拉雅-横断山区 在早渐新世(~32 Ma)两属出现,Roscoea在横断山区与喜马拉雅山脉的两个分支大概在渐新世-中新世之交分化 [54 ] Nanorana parkeri 青藏高原南部 高原东部和西部类群分化时间1.4~3.7 Ma [55 ] Paini 南亚、东南亚、青藏高原 在渐新世早期棘蛙族高原-喜马拉雅类群与中国第二阶梯上生活的类群分化,随后在在26.1~12.5 Ma高原生活类群与喜马拉雅山脉生活类群分化,高原内部生活类群在晚中新世到上新世期间分化速率加快 [22 ] Bufo tibetanus 青藏高原、四川、甘肃、陕西等 高原种类形成于上新世期间 [56 ] Phrynocephalus 亚洲干旱区 在7.2~13.0 Ma期间演化出胎生类群,3.2~7.3 Ma期间种间多样性增加 [57 ] Phrynocephalus erythrurus 羌塘盆地 在3.67 Ma左右北部类群与南部类群发生分化,约2.76 Ma北部类群内部进一步分化 [58 ] Gnaptorina 青藏高原及周边高海拔地区 始新世起源于青藏高原东南缘 [18 -19 ] Coelotinae 北半球温带、亚热带 晚始新世高原东南缘隆升阻碍南方隙蛛向北扩散,渐新世高原中部和北部隆起导致北方隙蛛早期多样化进程中的灭绝事件 [59 ] Montifringilla, Onychostruthus, Pyrgilauda 青藏高原 雪雀的共同祖先在约14 Ma就应经生活在青藏高原,并进化出适应高海拔极端环境的性状 [21 ] Parus major 亚洲、欧洲 蒙古分布类群与其他种类分化时间为2.8~0.7 Ma,喜马拉雅山脉东部与中国中东部类群分化时间为1.9~0.4 Ma [60 ] Pseudopodoces humilis 青藏高原、柴达木盆地周边高山 在9.9~7.7 Ma期间地山雀就与与其他山雀分化开来,并出现适应高海拔性状 [61 ] Phylloscopus, Seicercus 亚洲 喜马拉雅山脉分布类群在7 Ma加速进化 [62 -63 ] Perdix 西藏、甘肃、青海、四川、新疆 高原类群出现于3.63 (4.52~2.75) Ma [64 ] Pantholops hodgsonii 西藏、新疆、青海 上新世-更新世就已经出现在青藏高原 [65 ] Procapra picticaudata 西藏、四川、甘肃、青海、新疆 种内分化时间约2.2~4.4 Ma [66 ] Rhinopithecus 四川、甘肃、陕西、贵州、云南、缅甸、越南 北方类群川金丝猴、黔金丝猴与喜马拉雅类群滇金丝猴、缅甸金丝猴分化时间约1.6 Ma [67 ] Ochotona 北半球 分布于青藏高原的高山类群和灌丛草原类群与低海拔泛北极分布类群分化时间大约为2.4 Ma [68 ] Apodemus draco 四川、云南、甘肃、青海、陕西、湖北、湖南、福建、台湾 2.2 Ma (3.8~0.87 Ma)分化形成,后来由于高原的阶段性隆升以及冰川作用种系及遗传结构更加复杂 [69 -70 ] Eothenomys 东亚 高原种类与低海拔种类分裂时间约2.7 Ma [71 ] Schizothoracinae 青藏高原及周边地区 在晚中新世晚期或上新世裂腹鱼亚科鱼类就已经出现在青藏高原及周边水系中,在晚更新世期间种内分化加速,不同海拔之间的类群产生等级分化,种间多样性增加,第四纪以来青藏高原隆升导致的水系改组是推动裂腹鱼亚科鱼类演化的原动力 [31 ,76 -78 ]
3 讨论 最近,青藏高原古地貌重建工作取得了丰硕的成果(图3 ),但是各种指标重建结果往往存在矛盾(图4 )[25 ,79 -83 ] .同位素重建结果表明高原南部在古近纪早期就存在安第斯型的巨大山系[84 -85 ] ;古生物研究发现高原腹地从中始新世到晚渐新世都存在东西向的大峡谷,该峡谷底部海拔在2 000 m左右,两侧为高大的冈底斯山脉和羌塘山脉[82 ,86 -88 ] ,动植物如巨犀(Paraceratherium )[89 ] 、攀鲈(Eoanabas )[90 -91 ] 、臭椿(Ailanthus )[88 ] 、椿榆(Cedrelospermum )[92 ] 、兔耳果(Lagokarpos )[93 ] 等在古近纪都可以通过青藏地区峡谷与周边动植物区系交流,新近纪之前的青藏地区虽然地形复杂,但是尚未隆升成今天的高原;横断山区某些区域则在晚始新世-早渐新世之交就已经获得现今的海拔高度[94 -95 ] ;高原北部和喜马拉雅山脉则在新近纪形成现在的地貌格局[25 ,96 -97 ] .古生物研究也发现青藏高原在晚上新世就已经存在高寒生境和冰缘环境,成为第四纪冰期动物忍耐寒冷环境的“训练营”,第四纪冰期来临之后这些适应寒冷环境的生物才扩散至北半球高纬度地区[34 ,98 -99 ] .因此,高原及周边地区高寒生物类群的起源时间表明在渐新世青藏高原部分区域就已经出现高寒生态系统,晚中新世以来青藏高原地貌与气候继续发生剧烈变化,促使这些物种的祖先类群迁入或就地演化积累适应高寒环境的基因和形态,并最终演化成适应高寒生境的新物种[25 ] .高寒区物种在晚上新世的加速分化可能与高原巨大山系和水系、冰川活动等造成的地理隔离有关[26 ] .随着第四纪冰期的到来,青藏高原及周边地区的高寒生物区也开始与北半球高纬度地区存在密切联系[34 ,98 -99 ] .同时,演化生物学研究也发现在第四纪冰期期间并未形成覆盖整个青藏高原的大冰盖,高原面上仍存在很多生物避难所[26 ] . ...
Phylogenetic relationships of toads in the Bufo bufo species group from the eastern escarpment of the Tibetan Plateau: A case of vicariance and dispersal
2
1998
... 两栖动物中的高山倭蛙(Nanorana parkeri )分布于青藏高原南部海拔2 850~4 700 m之间的高山地区,其高原东部和西部类群分化时间约为3.7~1.4 Ma[55 ] .棘蛙族(Paini)物种多分布于深山急流中,结合其分子进化历史和生物地理分析发现在渐新世早期棘蛙族高原-喜马拉雅类群与中国第二阶梯上生活的类群分化,随后在26.1~12.5 Ma高原生活类群与喜马拉雅山脉生活类群分化,高原内部生活类群在晚中新世到上新世期间分化速率加快[22 ] .生活于青藏高原南部海拔2 850~4 700 m区域的西藏蟾蜍(Bufo tibetanus ),系统发育学研究表明其在上新世就演化出适应高原生活的性状[56 ] .爬行类动物沙蜥属(Phrynocephalus )一般分布于海拔超过2 200 m地区,为适应高原恶劣环境在约13.0~7.2 Ma进化出胎生类群,在7.3~3.2 Ma期间沙蜥属种间多样性快速增加[57 ] .分布于羌塘高原海拔4 500~5 000 m区域的红尾沙蜥(P.erythrurus )在3.67 Ma左右北部类群与南部类群发生分化,约2.76 Ma北部类群内部进一步分化,可能与晚上新世以来唐古拉山脉、乌兰乌拉山脉以及北麓河构造活动形成地理隔离有关[58 ] . ...
... Biological evolution in the alpine region of Tibetan Plateau
Table 1 种属 分布区域 适应高海拔类群分化时间或多样性增加时间 文献来源 Sinopodophyllum hexandrum 青藏高原东部 在约(6.52±1.89) Ma期间出现在青藏高原东部,但是直到3~2 Ma以来才演化出适应高山环境的形态 [47 ] Nannoglottis 青藏高原 在23~32 Ma期间出现在青藏高原,随后多样性增高,其高山灌丛和针叶林生活类型分支时间约为3.4 Ma [48 ] Reaumuria soongarica 西北干旱区、青藏高原北部 高原内部分布类群演化时间2.0~6.3 Ma,高原东北缘与高原西北类群分化时间1.1~3.5 Ma [49 ] Orchidinae 青藏高原及喜马拉雅山脉 青藏高原特有种大多是在过去6 Ma里进化而来 [50 ] Oxyria sinensis 青藏高原东南部 高山类群出现时间7.54~24.36 Ma,种内多样化增加时间1.74~0.86 Ma [51 ] Juniperus microsperma 西藏波密 早中新世21.6~14 Ma期间与其他种群分化 [52 ] Rhododendron 北半球 横断山区概属多样性在晚中新世(约8 Ma)快速增加 [53 ] Roscoea, Cautleya 喜马拉雅-横断山区 在早渐新世(~32 Ma)两属出现,Roscoea在横断山区与喜马拉雅山脉的两个分支大概在渐新世-中新世之交分化 [54 ] Nanorana parkeri 青藏高原南部 高原东部和西部类群分化时间1.4~3.7 Ma [55 ] Paini 南亚、东南亚、青藏高原 在渐新世早期棘蛙族高原-喜马拉雅类群与中国第二阶梯上生活的类群分化,随后在在26.1~12.5 Ma高原生活类群与喜马拉雅山脉生活类群分化,高原内部生活类群在晚中新世到上新世期间分化速率加快 [22 ] Bufo tibetanus 青藏高原、四川、甘肃、陕西等 高原种类形成于上新世期间 [56 ] Phrynocephalus 亚洲干旱区 在7.2~13.0 Ma期间演化出胎生类群,3.2~7.3 Ma期间种间多样性增加 [57 ] Phrynocephalus erythrurus 羌塘盆地 在3.67 Ma左右北部类群与南部类群发生分化,约2.76 Ma北部类群内部进一步分化 [58 ] Gnaptorina 青藏高原及周边高海拔地区 始新世起源于青藏高原东南缘 [18 -19 ] Coelotinae 北半球温带、亚热带 晚始新世高原东南缘隆升阻碍南方隙蛛向北扩散,渐新世高原中部和北部隆起导致北方隙蛛早期多样化进程中的灭绝事件 [59 ] Montifringilla, Onychostruthus, Pyrgilauda 青藏高原 雪雀的共同祖先在约14 Ma就应经生活在青藏高原,并进化出适应高海拔极端环境的性状 [21 ] Parus major 亚洲、欧洲 蒙古分布类群与其他种类分化时间为2.8~0.7 Ma,喜马拉雅山脉东部与中国中东部类群分化时间为1.9~0.4 Ma [60 ] Pseudopodoces humilis 青藏高原、柴达木盆地周边高山 在9.9~7.7 Ma期间地山雀就与与其他山雀分化开来,并出现适应高海拔性状 [61 ] Phylloscopus, Seicercus 亚洲 喜马拉雅山脉分布类群在7 Ma加速进化 [62 -63 ] Perdix 西藏、甘肃、青海、四川、新疆 高原类群出现于3.63 (4.52~2.75) Ma [64 ] Pantholops hodgsonii 西藏、新疆、青海 上新世-更新世就已经出现在青藏高原 [65 ] Procapra picticaudata 西藏、四川、甘肃、青海、新疆 种内分化时间约2.2~4.4 Ma [66 ] Rhinopithecus 四川、甘肃、陕西、贵州、云南、缅甸、越南 北方类群川金丝猴、黔金丝猴与喜马拉雅类群滇金丝猴、缅甸金丝猴分化时间约1.6 Ma [67 ] Ochotona 北半球 分布于青藏高原的高山类群和灌丛草原类群与低海拔泛北极分布类群分化时间大约为2.4 Ma [68 ] Apodemus draco 四川、云南、甘肃、青海、陕西、湖北、湖南、福建、台湾 2.2 Ma (3.8~0.87 Ma)分化形成,后来由于高原的阶段性隆升以及冰川作用种系及遗传结构更加复杂 [69 -70 ] Eothenomys 东亚 高原种类与低海拔种类分裂时间约2.7 Ma [71 ] Schizothoracinae 青藏高原及周边地区 在晚中新世晚期或上新世裂腹鱼亚科鱼类就已经出现在青藏高原及周边水系中,在晚更新世期间种内分化加速,不同海拔之间的类群产生等级分化,种间多样性增加,第四纪以来青藏高原隆升导致的水系改组是推动裂腹鱼亚科鱼类演化的原动力 [31 ,76 -78 ]
3 讨论 最近,青藏高原古地貌重建工作取得了丰硕的成果(图3 ),但是各种指标重建结果往往存在矛盾(图4 )[25 ,79 -83 ] .同位素重建结果表明高原南部在古近纪早期就存在安第斯型的巨大山系[84 -85 ] ;古生物研究发现高原腹地从中始新世到晚渐新世都存在东西向的大峡谷,该峡谷底部海拔在2 000 m左右,两侧为高大的冈底斯山脉和羌塘山脉[82 ,86 -88 ] ,动植物如巨犀(Paraceratherium )[89 ] 、攀鲈(Eoanabas )[90 -91 ] 、臭椿(Ailanthus )[88 ] 、椿榆(Cedrelospermum )[92 ] 、兔耳果(Lagokarpos )[93 ] 等在古近纪都可以通过青藏地区峡谷与周边动植物区系交流,新近纪之前的青藏地区虽然地形复杂,但是尚未隆升成今天的高原;横断山区某些区域则在晚始新世-早渐新世之交就已经获得现今的海拔高度[94 -95 ] ;高原北部和喜马拉雅山脉则在新近纪形成现在的地貌格局[25 ,96 -97 ] .古生物研究也发现青藏高原在晚上新世就已经存在高寒生境和冰缘环境,成为第四纪冰期动物忍耐寒冷环境的“训练营”,第四纪冰期来临之后这些适应寒冷环境的生物才扩散至北半球高纬度地区[34 ,98 -99 ] .因此,高原及周边地区高寒生物类群的起源时间表明在渐新世青藏高原部分区域就已经出现高寒生态系统,晚中新世以来青藏高原地貌与气候继续发生剧烈变化,促使这些物种的祖先类群迁入或就地演化积累适应高寒环境的基因和形态,并最终演化成适应高寒生境的新物种[25 ] .高寒区物种在晚上新世的加速分化可能与高原巨大山系和水系、冰川活动等造成的地理隔离有关[26 ] .随着第四纪冰期的到来,青藏高原及周边地区的高寒生物区也开始与北半球高纬度地区存在密切联系[34 ,98 -99 ] .同时,演化生物学研究也发现在第四纪冰期期间并未形成覆盖整个青藏高原的大冰盖,高原面上仍存在很多生物避难所[26 ] . ...
Species history and divergence times of viviparous and oviparous Chinese toad-headed sand lizards (Phrynocephalus ) on the Qinghai-Tibetan Plateau
2
2013
... 两栖动物中的高山倭蛙(Nanorana parkeri )分布于青藏高原南部海拔2 850~4 700 m之间的高山地区,其高原东部和西部类群分化时间约为3.7~1.4 Ma[55 ] .棘蛙族(Paini)物种多分布于深山急流中,结合其分子进化历史和生物地理分析发现在渐新世早期棘蛙族高原-喜马拉雅类群与中国第二阶梯上生活的类群分化,随后在26.1~12.5 Ma高原生活类群与喜马拉雅山脉生活类群分化,高原内部生活类群在晚中新世到上新世期间分化速率加快[22 ] .生活于青藏高原南部海拔2 850~4 700 m区域的西藏蟾蜍(Bufo tibetanus ),系统发育学研究表明其在上新世就演化出适应高原生活的性状[56 ] .爬行类动物沙蜥属(Phrynocephalus )一般分布于海拔超过2 200 m地区,为适应高原恶劣环境在约13.0~7.2 Ma进化出胎生类群,在7.3~3.2 Ma期间沙蜥属种间多样性快速增加[57 ] .分布于羌塘高原海拔4 500~5 000 m区域的红尾沙蜥(P.erythrurus )在3.67 Ma左右北部类群与南部类群发生分化,约2.76 Ma北部类群内部进一步分化,可能与晚上新世以来唐古拉山脉、乌兰乌拉山脉以及北麓河构造活动形成地理隔离有关[58 ] . ...
... Biological evolution in the alpine region of Tibetan Plateau
Table 1 种属 分布区域 适应高海拔类群分化时间或多样性增加时间 文献来源 Sinopodophyllum hexandrum 青藏高原东部 在约(6.52±1.89) Ma期间出现在青藏高原东部,但是直到3~2 Ma以来才演化出适应高山环境的形态 [47 ] Nannoglottis 青藏高原 在23~32 Ma期间出现在青藏高原,随后多样性增高,其高山灌丛和针叶林生活类型分支时间约为3.4 Ma [48 ] Reaumuria soongarica 西北干旱区、青藏高原北部 高原内部分布类群演化时间2.0~6.3 Ma,高原东北缘与高原西北类群分化时间1.1~3.5 Ma [49 ] Orchidinae 青藏高原及喜马拉雅山脉 青藏高原特有种大多是在过去6 Ma里进化而来 [50 ] Oxyria sinensis 青藏高原东南部 高山类群出现时间7.54~24.36 Ma,种内多样化增加时间1.74~0.86 Ma [51 ] Juniperus microsperma 西藏波密 早中新世21.6~14 Ma期间与其他种群分化 [52 ] Rhododendron 北半球 横断山区概属多样性在晚中新世(约8 Ma)快速增加 [53 ] Roscoea, Cautleya 喜马拉雅-横断山区 在早渐新世(~32 Ma)两属出现,Roscoea在横断山区与喜马拉雅山脉的两个分支大概在渐新世-中新世之交分化 [54 ] Nanorana parkeri 青藏高原南部 高原东部和西部类群分化时间1.4~3.7 Ma [55 ] Paini 南亚、东南亚、青藏高原 在渐新世早期棘蛙族高原-喜马拉雅类群与中国第二阶梯上生活的类群分化,随后在在26.1~12.5 Ma高原生活类群与喜马拉雅山脉生活类群分化,高原内部生活类群在晚中新世到上新世期间分化速率加快 [22 ] Bufo tibetanus 青藏高原、四川、甘肃、陕西等 高原种类形成于上新世期间 [56 ] Phrynocephalus 亚洲干旱区 在7.2~13.0 Ma期间演化出胎生类群,3.2~7.3 Ma期间种间多样性增加 [57 ] Phrynocephalus erythrurus 羌塘盆地 在3.67 Ma左右北部类群与南部类群发生分化,约2.76 Ma北部类群内部进一步分化 [58 ] Gnaptorina 青藏高原及周边高海拔地区 始新世起源于青藏高原东南缘 [18 -19 ] Coelotinae 北半球温带、亚热带 晚始新世高原东南缘隆升阻碍南方隙蛛向北扩散,渐新世高原中部和北部隆起导致北方隙蛛早期多样化进程中的灭绝事件 [59 ] Montifringilla, Onychostruthus, Pyrgilauda 青藏高原 雪雀的共同祖先在约14 Ma就应经生活在青藏高原,并进化出适应高海拔极端环境的性状 [21 ] Parus major 亚洲、欧洲 蒙古分布类群与其他种类分化时间为2.8~0.7 Ma,喜马拉雅山脉东部与中国中东部类群分化时间为1.9~0.4 Ma [60 ] Pseudopodoces humilis 青藏高原、柴达木盆地周边高山 在9.9~7.7 Ma期间地山雀就与与其他山雀分化开来,并出现适应高海拔性状 [61 ] Phylloscopus, Seicercus 亚洲 喜马拉雅山脉分布类群在7 Ma加速进化 [62 -63 ] Perdix 西藏、甘肃、青海、四川、新疆 高原类群出现于3.63 (4.52~2.75) Ma [64 ] Pantholops hodgsonii 西藏、新疆、青海 上新世-更新世就已经出现在青藏高原 [65 ] Procapra picticaudata 西藏、四川、甘肃、青海、新疆 种内分化时间约2.2~4.4 Ma [66 ] Rhinopithecus 四川、甘肃、陕西、贵州、云南、缅甸、越南 北方类群川金丝猴、黔金丝猴与喜马拉雅类群滇金丝猴、缅甸金丝猴分化时间约1.6 Ma [67 ] Ochotona 北半球 分布于青藏高原的高山类群和灌丛草原类群与低海拔泛北极分布类群分化时间大约为2.4 Ma [68 ] Apodemus draco 四川、云南、甘肃、青海、陕西、湖北、湖南、福建、台湾 2.2 Ma (3.8~0.87 Ma)分化形成,后来由于高原的阶段性隆升以及冰川作用种系及遗传结构更加复杂 [69 -70 ] Eothenomys 东亚 高原种类与低海拔种类分裂时间约2.7 Ma [71 ] Schizothoracinae 青藏高原及周边地区 在晚中新世晚期或上新世裂腹鱼亚科鱼类就已经出现在青藏高原及周边水系中,在晚更新世期间种内分化加速,不同海拔之间的类群产生等级分化,种间多样性增加,第四纪以来青藏高原隆升导致的水系改组是推动裂腹鱼亚科鱼类演化的原动力 [31 ,76 -78 ]
3 讨论 最近,青藏高原古地貌重建工作取得了丰硕的成果(图3 ),但是各种指标重建结果往往存在矛盾(图4 )[25 ,79 -83 ] .同位素重建结果表明高原南部在古近纪早期就存在安第斯型的巨大山系[84 -85 ] ;古生物研究发现高原腹地从中始新世到晚渐新世都存在东西向的大峡谷,该峡谷底部海拔在2 000 m左右,两侧为高大的冈底斯山脉和羌塘山脉[82 ,86 -88 ] ,动植物如巨犀(Paraceratherium )[89 ] 、攀鲈(Eoanabas )[90 -91 ] 、臭椿(Ailanthus )[88 ] 、椿榆(Cedrelospermum )[92 ] 、兔耳果(Lagokarpos )[93 ] 等在古近纪都可以通过青藏地区峡谷与周边动植物区系交流,新近纪之前的青藏地区虽然地形复杂,但是尚未隆升成今天的高原;横断山区某些区域则在晚始新世-早渐新世之交就已经获得现今的海拔高度[94 -95 ] ;高原北部和喜马拉雅山脉则在新近纪形成现在的地貌格局[25 ,96 -97 ] .古生物研究也发现青藏高原在晚上新世就已经存在高寒生境和冰缘环境,成为第四纪冰期动物忍耐寒冷环境的“训练营”,第四纪冰期来临之后这些适应寒冷环境的生物才扩散至北半球高纬度地区[34 ,98 -99 ] .因此,高原及周边地区高寒生物类群的起源时间表明在渐新世青藏高原部分区域就已经出现高寒生态系统,晚中新世以来青藏高原地貌与气候继续发生剧烈变化,促使这些物种的祖先类群迁入或就地演化积累适应高寒环境的基因和形态,并最终演化成适应高寒生境的新物种[25 ] .高寒区物种在晚上新世的加速分化可能与高原巨大山系和水系、冰川活动等造成的地理隔离有关[26 ] .随着第四纪冰期的到来,青藏高原及周边地区的高寒生物区也开始与北半球高纬度地区存在密切联系[34 ,98 -99 ] .同时,演化生物学研究也发现在第四纪冰期期间并未形成覆盖整个青藏高原的大冰盖,高原面上仍存在很多生物避难所[26 ] . ...
Phylogeography of Phrynocephalus erythrurus from the Qiangtang Plateau of the Tibetan Plateau
2
2010
... 两栖动物中的高山倭蛙(Nanorana parkeri )分布于青藏高原南部海拔2 850~4 700 m之间的高山地区,其高原东部和西部类群分化时间约为3.7~1.4 Ma[55 ] .棘蛙族(Paini)物种多分布于深山急流中,结合其分子进化历史和生物地理分析发现在渐新世早期棘蛙族高原-喜马拉雅类群与中国第二阶梯上生活的类群分化,随后在26.1~12.5 Ma高原生活类群与喜马拉雅山脉生活类群分化,高原内部生活类群在晚中新世到上新世期间分化速率加快[22 ] .生活于青藏高原南部海拔2 850~4 700 m区域的西藏蟾蜍(Bufo tibetanus ),系统发育学研究表明其在上新世就演化出适应高原生活的性状[56 ] .爬行类动物沙蜥属(Phrynocephalus )一般分布于海拔超过2 200 m地区,为适应高原恶劣环境在约13.0~7.2 Ma进化出胎生类群,在7.3~3.2 Ma期间沙蜥属种间多样性快速增加[57 ] .分布于羌塘高原海拔4 500~5 000 m区域的红尾沙蜥(P.erythrurus )在3.67 Ma左右北部类群与南部类群发生分化,约2.76 Ma北部类群内部进一步分化,可能与晚上新世以来唐古拉山脉、乌兰乌拉山脉以及北麓河构造活动形成地理隔离有关[58 ] . ...
... Biological evolution in the alpine region of Tibetan Plateau
Table 1 种属 分布区域 适应高海拔类群分化时间或多样性增加时间 文献来源 Sinopodophyllum hexandrum 青藏高原东部 在约(6.52±1.89) Ma期间出现在青藏高原东部,但是直到3~2 Ma以来才演化出适应高山环境的形态 [47 ] Nannoglottis 青藏高原 在23~32 Ma期间出现在青藏高原,随后多样性增高,其高山灌丛和针叶林生活类型分支时间约为3.4 Ma [48 ] Reaumuria soongarica 西北干旱区、青藏高原北部 高原内部分布类群演化时间2.0~6.3 Ma,高原东北缘与高原西北类群分化时间1.1~3.5 Ma [49 ] Orchidinae 青藏高原及喜马拉雅山脉 青藏高原特有种大多是在过去6 Ma里进化而来 [50 ] Oxyria sinensis 青藏高原东南部 高山类群出现时间7.54~24.36 Ma,种内多样化增加时间1.74~0.86 Ma [51 ] Juniperus microsperma 西藏波密 早中新世21.6~14 Ma期间与其他种群分化 [52 ] Rhododendron 北半球 横断山区概属多样性在晚中新世(约8 Ma)快速增加 [53 ] Roscoea, Cautleya 喜马拉雅-横断山区 在早渐新世(~32 Ma)两属出现,Roscoea在横断山区与喜马拉雅山脉的两个分支大概在渐新世-中新世之交分化 [54 ] Nanorana parkeri 青藏高原南部 高原东部和西部类群分化时间1.4~3.7 Ma [55 ] Paini 南亚、东南亚、青藏高原 在渐新世早期棘蛙族高原-喜马拉雅类群与中国第二阶梯上生活的类群分化,随后在在26.1~12.5 Ma高原生活类群与喜马拉雅山脉生活类群分化,高原内部生活类群在晚中新世到上新世期间分化速率加快 [22 ] Bufo tibetanus 青藏高原、四川、甘肃、陕西等 高原种类形成于上新世期间 [56 ] Phrynocephalus 亚洲干旱区 在7.2~13.0 Ma期间演化出胎生类群,3.2~7.3 Ma期间种间多样性增加 [57 ] Phrynocephalus erythrurus 羌塘盆地 在3.67 Ma左右北部类群与南部类群发生分化,约2.76 Ma北部类群内部进一步分化 [58 ] Gnaptorina 青藏高原及周边高海拔地区 始新世起源于青藏高原东南缘 [18 -19 ] Coelotinae 北半球温带、亚热带 晚始新世高原东南缘隆升阻碍南方隙蛛向北扩散,渐新世高原中部和北部隆起导致北方隙蛛早期多样化进程中的灭绝事件 [59 ] Montifringilla, Onychostruthus, Pyrgilauda 青藏高原 雪雀的共同祖先在约14 Ma就应经生活在青藏高原,并进化出适应高海拔极端环境的性状 [21 ] Parus major 亚洲、欧洲 蒙古分布类群与其他种类分化时间为2.8~0.7 Ma,喜马拉雅山脉东部与中国中东部类群分化时间为1.9~0.4 Ma [60 ] Pseudopodoces humilis 青藏高原、柴达木盆地周边高山 在9.9~7.7 Ma期间地山雀就与与其他山雀分化开来,并出现适应高海拔性状 [61 ] Phylloscopus, Seicercus 亚洲 喜马拉雅山脉分布类群在7 Ma加速进化 [62 -63 ] Perdix 西藏、甘肃、青海、四川、新疆 高原类群出现于3.63 (4.52~2.75) Ma [64 ] Pantholops hodgsonii 西藏、新疆、青海 上新世-更新世就已经出现在青藏高原 [65 ] Procapra picticaudata 西藏、四川、甘肃、青海、新疆 种内分化时间约2.2~4.4 Ma [66 ] Rhinopithecus 四川、甘肃、陕西、贵州、云南、缅甸、越南 北方类群川金丝猴、黔金丝猴与喜马拉雅类群滇金丝猴、缅甸金丝猴分化时间约1.6 Ma [67 ] Ochotona 北半球 分布于青藏高原的高山类群和灌丛草原类群与低海拔泛北极分布类群分化时间大约为2.4 Ma [68 ] Apodemus draco 四川、云南、甘肃、青海、陕西、湖北、湖南、福建、台湾 2.2 Ma (3.8~0.87 Ma)分化形成,后来由于高原的阶段性隆升以及冰川作用种系及遗传结构更加复杂 [69 -70 ] Eothenomys 东亚 高原种类与低海拔种类分裂时间约2.7 Ma [71 ] Schizothoracinae 青藏高原及周边地区 在晚中新世晚期或上新世裂腹鱼亚科鱼类就已经出现在青藏高原及周边水系中,在晚更新世期间种内分化加速,不同海拔之间的类群产生等级分化,种间多样性增加,第四纪以来青藏高原隆升导致的水系改组是推动裂腹鱼亚科鱼类演化的原动力 [31 ,76 -78 ]
3 讨论 最近,青藏高原古地貌重建工作取得了丰硕的成果(图3 ),但是各种指标重建结果往往存在矛盾(图4 )[25 ,79 -83 ] .同位素重建结果表明高原南部在古近纪早期就存在安第斯型的巨大山系[84 -85 ] ;古生物研究发现高原腹地从中始新世到晚渐新世都存在东西向的大峡谷,该峡谷底部海拔在2 000 m左右,两侧为高大的冈底斯山脉和羌塘山脉[82 ,86 -88 ] ,动植物如巨犀(Paraceratherium )[89 ] 、攀鲈(Eoanabas )[90 -91 ] 、臭椿(Ailanthus )[88 ] 、椿榆(Cedrelospermum )[92 ] 、兔耳果(Lagokarpos )[93 ] 等在古近纪都可以通过青藏地区峡谷与周边动植物区系交流,新近纪之前的青藏地区虽然地形复杂,但是尚未隆升成今天的高原;横断山区某些区域则在晚始新世-早渐新世之交就已经获得现今的海拔高度[94 -95 ] ;高原北部和喜马拉雅山脉则在新近纪形成现在的地貌格局[25 ,96 -97 ] .古生物研究也发现青藏高原在晚上新世就已经存在高寒生境和冰缘环境,成为第四纪冰期动物忍耐寒冷环境的“训练营”,第四纪冰期来临之后这些适应寒冷环境的生物才扩散至北半球高纬度地区[34 ,98 -99 ] .因此,高原及周边地区高寒生物类群的起源时间表明在渐新世青藏高原部分区域就已经出现高寒生态系统,晚中新世以来青藏高原地貌与气候继续发生剧烈变化,促使这些物种的祖先类群迁入或就地演化积累适应高寒环境的基因和形态,并最终演化成适应高寒生境的新物种[25 ] .高寒区物种在晚上新世的加速分化可能与高原巨大山系和水系、冰川活动等造成的地理隔离有关[26 ] .随着第四纪冰期的到来,青藏高原及周边地区的高寒生物区也开始与北半球高纬度地区存在密切联系[34 ,98 -99 ] .同时,演化生物学研究也发现在第四纪冰期期间并未形成覆盖整个青藏高原的大冰盖,高原面上仍存在很多生物避难所[26 ] . ...
Extinction vs. Rapid radiation: The juxtaposed evolutionary histories of coelotine spiders support the Eocene-Oligocene orogenesis of the Tibetan Plateau
2
2017
... 小琵甲属(Gnaptorina )是琵甲族的高原特有属,已知的34种全部分布在青藏高原及周边地区海拔3 000~4 500 m的区域,该属昆虫后翅愈合,不能飞翔,是对高海拔极端环境的一种适应性状,小琵甲属生物地理学研究表明其祖先类群在始新世起源于青藏高原东南缘,高原隆升和河流切割造成的地理隔离阻断了小琵甲属物种或种群之间的基因交流,加速特有种的快速辐射演化[18 -19 ] ;隙蛛(Coelotinae )是北半球温带和亚热带地区常见蜘蛛种类,青藏高原东南缘晚始新世形成的高海拔地貌阻碍了南方隙蛛向北扩散,但是随之形成的复杂地貌创造的局部地理隔离也促使南方隙蛛在此阶段快速辐射演化,渐新世以来高原中部和北部的隆起导致北方隙蛛在早期多样化过程中经历了灭绝事件[59 ] . ...
... Biological evolution in the alpine region of Tibetan Plateau
Table 1 种属 分布区域 适应高海拔类群分化时间或多样性增加时间 文献来源 Sinopodophyllum hexandrum 青藏高原东部 在约(6.52±1.89) Ma期间出现在青藏高原东部,但是直到3~2 Ma以来才演化出适应高山环境的形态 [47 ] Nannoglottis 青藏高原 在23~32 Ma期间出现在青藏高原,随后多样性增高,其高山灌丛和针叶林生活类型分支时间约为3.4 Ma [48 ] Reaumuria soongarica 西北干旱区、青藏高原北部 高原内部分布类群演化时间2.0~6.3 Ma,高原东北缘与高原西北类群分化时间1.1~3.5 Ma [49 ] Orchidinae 青藏高原及喜马拉雅山脉 青藏高原特有种大多是在过去6 Ma里进化而来 [50 ] Oxyria sinensis 青藏高原东南部 高山类群出现时间7.54~24.36 Ma,种内多样化增加时间1.74~0.86 Ma [51 ] Juniperus microsperma 西藏波密 早中新世21.6~14 Ma期间与其他种群分化 [52 ] Rhododendron 北半球 横断山区概属多样性在晚中新世(约8 Ma)快速增加 [53 ] Roscoea, Cautleya 喜马拉雅-横断山区 在早渐新世(~32 Ma)两属出现,Roscoea在横断山区与喜马拉雅山脉的两个分支大概在渐新世-中新世之交分化 [54 ] Nanorana parkeri 青藏高原南部 高原东部和西部类群分化时间1.4~3.7 Ma [55 ] Paini 南亚、东南亚、青藏高原 在渐新世早期棘蛙族高原-喜马拉雅类群与中国第二阶梯上生活的类群分化,随后在在26.1~12.5 Ma高原生活类群与喜马拉雅山脉生活类群分化,高原内部生活类群在晚中新世到上新世期间分化速率加快 [22 ] Bufo tibetanus 青藏高原、四川、甘肃、陕西等 高原种类形成于上新世期间 [56 ] Phrynocephalus 亚洲干旱区 在7.2~13.0 Ma期间演化出胎生类群,3.2~7.3 Ma期间种间多样性增加 [57 ] Phrynocephalus erythrurus 羌塘盆地 在3.67 Ma左右北部类群与南部类群发生分化,约2.76 Ma北部类群内部进一步分化 [58 ] Gnaptorina 青藏高原及周边高海拔地区 始新世起源于青藏高原东南缘 [18 -19 ] Coelotinae 北半球温带、亚热带 晚始新世高原东南缘隆升阻碍南方隙蛛向北扩散,渐新世高原中部和北部隆起导致北方隙蛛早期多样化进程中的灭绝事件 [59 ] Montifringilla, Onychostruthus, Pyrgilauda 青藏高原 雪雀的共同祖先在约14 Ma就应经生活在青藏高原,并进化出适应高海拔极端环境的性状 [21 ] Parus major 亚洲、欧洲 蒙古分布类群与其他种类分化时间为2.8~0.7 Ma,喜马拉雅山脉东部与中国中东部类群分化时间为1.9~0.4 Ma [60 ] Pseudopodoces humilis 青藏高原、柴达木盆地周边高山 在9.9~7.7 Ma期间地山雀就与与其他山雀分化开来,并出现适应高海拔性状 [61 ] Phylloscopus, Seicercus 亚洲 喜马拉雅山脉分布类群在7 Ma加速进化 [62 -63 ] Perdix 西藏、甘肃、青海、四川、新疆 高原类群出现于3.63 (4.52~2.75) Ma [64 ] Pantholops hodgsonii 西藏、新疆、青海 上新世-更新世就已经出现在青藏高原 [65 ] Procapra picticaudata 西藏、四川、甘肃、青海、新疆 种内分化时间约2.2~4.4 Ma [66 ] Rhinopithecus 四川、甘肃、陕西、贵州、云南、缅甸、越南 北方类群川金丝猴、黔金丝猴与喜马拉雅类群滇金丝猴、缅甸金丝猴分化时间约1.6 Ma [67 ] Ochotona 北半球 分布于青藏高原的高山类群和灌丛草原类群与低海拔泛北极分布类群分化时间大约为2.4 Ma [68 ] Apodemus draco 四川、云南、甘肃、青海、陕西、湖北、湖南、福建、台湾 2.2 Ma (3.8~0.87 Ma)分化形成,后来由于高原的阶段性隆升以及冰川作用种系及遗传结构更加复杂 [69 -70 ] Eothenomys 东亚 高原种类与低海拔种类分裂时间约2.7 Ma [71 ] Schizothoracinae 青藏高原及周边地区 在晚中新世晚期或上新世裂腹鱼亚科鱼类就已经出现在青藏高原及周边水系中,在晚更新世期间种内分化加速,不同海拔之间的类群产生等级分化,种间多样性增加,第四纪以来青藏高原隆升导致的水系改组是推动裂腹鱼亚科鱼类演化的原动力 [31 ,76 -78 ]
3 讨论 最近,青藏高原古地貌重建工作取得了丰硕的成果(图3 ),但是各种指标重建结果往往存在矛盾(图4 )[25 ,79 -83 ] .同位素重建结果表明高原南部在古近纪早期就存在安第斯型的巨大山系[84 -85 ] ;古生物研究发现高原腹地从中始新世到晚渐新世都存在东西向的大峡谷,该峡谷底部海拔在2 000 m左右,两侧为高大的冈底斯山脉和羌塘山脉[82 ,86 -88 ] ,动植物如巨犀(Paraceratherium )[89 ] 、攀鲈(Eoanabas )[90 -91 ] 、臭椿(Ailanthus )[88 ] 、椿榆(Cedrelospermum )[92 ] 、兔耳果(Lagokarpos )[93 ] 等在古近纪都可以通过青藏地区峡谷与周边动植物区系交流,新近纪之前的青藏地区虽然地形复杂,但是尚未隆升成今天的高原;横断山区某些区域则在晚始新世-早渐新世之交就已经获得现今的海拔高度[94 -95 ] ;高原北部和喜马拉雅山脉则在新近纪形成现在的地貌格局[25 ,96 -97 ] .古生物研究也发现青藏高原在晚上新世就已经存在高寒生境和冰缘环境,成为第四纪冰期动物忍耐寒冷环境的“训练营”,第四纪冰期来临之后这些适应寒冷环境的生物才扩散至北半球高纬度地区[34 ,98 -99 ] .因此,高原及周边地区高寒生物类群的起源时间表明在渐新世青藏高原部分区域就已经出现高寒生态系统,晚中新世以来青藏高原地貌与气候继续发生剧烈变化,促使这些物种的祖先类群迁入或就地演化积累适应高寒环境的基因和形态,并最终演化成适应高寒生境的新物种[25 ] .高寒区物种在晚上新世的加速分化可能与高原巨大山系和水系、冰川活动等造成的地理隔离有关[26 ] .随着第四纪冰期的到来,青藏高原及周边地区的高寒生物区也开始与北半球高纬度地区存在密切联系[34 ,98 -99 ] .同时,演化生物学研究也发现在第四纪冰期期间并未形成覆盖整个青藏高原的大冰盖,高原面上仍存在很多生物避难所[26 ] . ...
Genetic responses to seasonal variation in altitudinal stress: whole-genome resequencing of great tit in eastern Himalayas
2
2015
... 鸟类中的雪雀(Montifringilla , Onychostruthus , Pyrgilauda )是在青藏高原低温、缺氧、强紫外线极端环境下繁衍壮大的类群,其全基因组测序与蛋白质功能研究发现雪雀的共同祖先在约14 Ma就应经生活在青藏高原,并进化出适应高海拔极端环境的性状[21 ] .大山雀(Parus major )在北半球广泛分布,第四纪以来高原周边地区构造隆升以及全球气候变干等形成地理隔离,使得高原分布类群与周边地区分布的姐妹类群发生分化,多样性升高[60 ] .地山雀(Pseudopodoces humilis )在青藏高原分布的海拔较高,一般超过4 000 m,系统发育研究发现在9.9~7.7 Ma期间地山雀就与其他山雀分化开来,并进化出适应高海拔生境的形态和习性[61 ] .对喜马拉雅山脉鸣禽的研究表明,山脉隆升和季风演化等原因导致柳莺属(Phylloscopus )和鹟莺属(Seicercus )在7 Ma左右开始加速分化[62 -63 ] .山鹑属(Perdix )广泛分布于中国西北和西南地区,其中高原山鹑(P. hodgsoniae )分布于青藏高原海拔2 500~5 000 m范围内,系统发育研究结果表明其适应高原生境的类群出现于4.52~2.75 Ma期间[64 ] . ...
... Biological evolution in the alpine region of Tibetan Plateau
Table 1 种属 分布区域 适应高海拔类群分化时间或多样性增加时间 文献来源 Sinopodophyllum hexandrum 青藏高原东部 在约(6.52±1.89) Ma期间出现在青藏高原东部,但是直到3~2 Ma以来才演化出适应高山环境的形态 [47 ] Nannoglottis 青藏高原 在23~32 Ma期间出现在青藏高原,随后多样性增高,其高山灌丛和针叶林生活类型分支时间约为3.4 Ma [48 ] Reaumuria soongarica 西北干旱区、青藏高原北部 高原内部分布类群演化时间2.0~6.3 Ma,高原东北缘与高原西北类群分化时间1.1~3.5 Ma [49 ] Orchidinae 青藏高原及喜马拉雅山脉 青藏高原特有种大多是在过去6 Ma里进化而来 [50 ] Oxyria sinensis 青藏高原东南部 高山类群出现时间7.54~24.36 Ma,种内多样化增加时间1.74~0.86 Ma [51 ] Juniperus microsperma 西藏波密 早中新世21.6~14 Ma期间与其他种群分化 [52 ] Rhododendron 北半球 横断山区概属多样性在晚中新世(约8 Ma)快速增加 [53 ] Roscoea, Cautleya 喜马拉雅-横断山区 在早渐新世(~32 Ma)两属出现,Roscoea在横断山区与喜马拉雅山脉的两个分支大概在渐新世-中新世之交分化 [54 ] Nanorana parkeri 青藏高原南部 高原东部和西部类群分化时间1.4~3.7 Ma [55 ] Paini 南亚、东南亚、青藏高原 在渐新世早期棘蛙族高原-喜马拉雅类群与中国第二阶梯上生活的类群分化,随后在在26.1~12.5 Ma高原生活类群与喜马拉雅山脉生活类群分化,高原内部生活类群在晚中新世到上新世期间分化速率加快 [22 ] Bufo tibetanus 青藏高原、四川、甘肃、陕西等 高原种类形成于上新世期间 [56 ] Phrynocephalus 亚洲干旱区 在7.2~13.0 Ma期间演化出胎生类群,3.2~7.3 Ma期间种间多样性增加 [57 ] Phrynocephalus erythrurus 羌塘盆地 在3.67 Ma左右北部类群与南部类群发生分化,约2.76 Ma北部类群内部进一步分化 [58 ] Gnaptorina 青藏高原及周边高海拔地区 始新世起源于青藏高原东南缘 [18 -19 ] Coelotinae 北半球温带、亚热带 晚始新世高原东南缘隆升阻碍南方隙蛛向北扩散,渐新世高原中部和北部隆起导致北方隙蛛早期多样化进程中的灭绝事件 [59 ] Montifringilla, Onychostruthus, Pyrgilauda 青藏高原 雪雀的共同祖先在约14 Ma就应经生活在青藏高原,并进化出适应高海拔极端环境的性状 [21 ] Parus major 亚洲、欧洲 蒙古分布类群与其他种类分化时间为2.8~0.7 Ma,喜马拉雅山脉东部与中国中东部类群分化时间为1.9~0.4 Ma [60 ] Pseudopodoces humilis 青藏高原、柴达木盆地周边高山 在9.9~7.7 Ma期间地山雀就与与其他山雀分化开来,并出现适应高海拔性状 [61 ] Phylloscopus, Seicercus 亚洲 喜马拉雅山脉分布类群在7 Ma加速进化 [62 -63 ] Perdix 西藏、甘肃、青海、四川、新疆 高原类群出现于3.63 (4.52~2.75) Ma [64 ] Pantholops hodgsonii 西藏、新疆、青海 上新世-更新世就已经出现在青藏高原 [65 ] Procapra picticaudata 西藏、四川、甘肃、青海、新疆 种内分化时间约2.2~4.4 Ma [66 ] Rhinopithecus 四川、甘肃、陕西、贵州、云南、缅甸、越南 北方类群川金丝猴、黔金丝猴与喜马拉雅类群滇金丝猴、缅甸金丝猴分化时间约1.6 Ma [67 ] Ochotona 北半球 分布于青藏高原的高山类群和灌丛草原类群与低海拔泛北极分布类群分化时间大约为2.4 Ma [68 ] Apodemus draco 四川、云南、甘肃、青海、陕西、湖北、湖南、福建、台湾 2.2 Ma (3.8~0.87 Ma)分化形成,后来由于高原的阶段性隆升以及冰川作用种系及遗传结构更加复杂 [69 -70 ] Eothenomys 东亚 高原种类与低海拔种类分裂时间约2.7 Ma [71 ] Schizothoracinae 青藏高原及周边地区 在晚中新世晚期或上新世裂腹鱼亚科鱼类就已经出现在青藏高原及周边水系中,在晚更新世期间种内分化加速,不同海拔之间的类群产生等级分化,种间多样性增加,第四纪以来青藏高原隆升导致的水系改组是推动裂腹鱼亚科鱼类演化的原动力 [31 ,76 -78 ]
3 讨论 最近,青藏高原古地貌重建工作取得了丰硕的成果(图3 ),但是各种指标重建结果往往存在矛盾(图4 )[25 ,79 -83 ] .同位素重建结果表明高原南部在古近纪早期就存在安第斯型的巨大山系[84 -85 ] ;古生物研究发现高原腹地从中始新世到晚渐新世都存在东西向的大峡谷,该峡谷底部海拔在2 000 m左右,两侧为高大的冈底斯山脉和羌塘山脉[82 ,86 -88 ] ,动植物如巨犀(Paraceratherium )[89 ] 、攀鲈(Eoanabas )[90 -91 ] 、臭椿(Ailanthus )[88 ] 、椿榆(Cedrelospermum )[92 ] 、兔耳果(Lagokarpos )[93 ] 等在古近纪都可以通过青藏地区峡谷与周边动植物区系交流,新近纪之前的青藏地区虽然地形复杂,但是尚未隆升成今天的高原;横断山区某些区域则在晚始新世-早渐新世之交就已经获得现今的海拔高度[94 -95 ] ;高原北部和喜马拉雅山脉则在新近纪形成现在的地貌格局[25 ,96 -97 ] .古生物研究也发现青藏高原在晚上新世就已经存在高寒生境和冰缘环境,成为第四纪冰期动物忍耐寒冷环境的“训练营”,第四纪冰期来临之后这些适应寒冷环境的生物才扩散至北半球高纬度地区[34 ,98 -99 ] .因此,高原及周边地区高寒生物类群的起源时间表明在渐新世青藏高原部分区域就已经出现高寒生态系统,晚中新世以来青藏高原地貌与气候继续发生剧烈变化,促使这些物种的祖先类群迁入或就地演化积累适应高寒环境的基因和形态,并最终演化成适应高寒生境的新物种[25 ] .高寒区物种在晚上新世的加速分化可能与高原巨大山系和水系、冰川活动等造成的地理隔离有关[26 ] .随着第四纪冰期的到来,青藏高原及周边地区的高寒生物区也开始与北半球高纬度地区存在密切联系[34 ,98 -99 ] .同时,演化生物学研究也发现在第四纪冰期期间并未形成覆盖整个青藏高原的大冰盖,高原面上仍存在很多生物避难所[26 ] . ...
Ground tit genome reveals avian adaptation to living at high altitudes in the Tibetan Plateau
2
2013
... 鸟类中的雪雀(Montifringilla , Onychostruthus , Pyrgilauda )是在青藏高原低温、缺氧、强紫外线极端环境下繁衍壮大的类群,其全基因组测序与蛋白质功能研究发现雪雀的共同祖先在约14 Ma就应经生活在青藏高原,并进化出适应高海拔极端环境的性状[21 ] .大山雀(Parus major )在北半球广泛分布,第四纪以来高原周边地区构造隆升以及全球气候变干等形成地理隔离,使得高原分布类群与周边地区分布的姐妹类群发生分化,多样性升高[60 ] .地山雀(Pseudopodoces humilis )在青藏高原分布的海拔较高,一般超过4 000 m,系统发育研究发现在9.9~7.7 Ma期间地山雀就与其他山雀分化开来,并进化出适应高海拔生境的形态和习性[61 ] .对喜马拉雅山脉鸣禽的研究表明,山脉隆升和季风演化等原因导致柳莺属(Phylloscopus )和鹟莺属(Seicercus )在7 Ma左右开始加速分化[62 -63 ] .山鹑属(Perdix )广泛分布于中国西北和西南地区,其中高原山鹑(P. hodgsoniae )分布于青藏高原海拔2 500~5 000 m范围内,系统发育研究结果表明其适应高原生境的类群出现于4.52~2.75 Ma期间[64 ] . ...
... Biological evolution in the alpine region of Tibetan Plateau
Table 1 种属 分布区域 适应高海拔类群分化时间或多样性增加时间 文献来源 Sinopodophyllum hexandrum 青藏高原东部 在约(6.52±1.89) Ma期间出现在青藏高原东部,但是直到3~2 Ma以来才演化出适应高山环境的形态 [47 ] Nannoglottis 青藏高原 在23~32 Ma期间出现在青藏高原,随后多样性增高,其高山灌丛和针叶林生活类型分支时间约为3.4 Ma [48 ] Reaumuria soongarica 西北干旱区、青藏高原北部 高原内部分布类群演化时间2.0~6.3 Ma,高原东北缘与高原西北类群分化时间1.1~3.5 Ma [49 ] Orchidinae 青藏高原及喜马拉雅山脉 青藏高原特有种大多是在过去6 Ma里进化而来 [50 ] Oxyria sinensis 青藏高原东南部 高山类群出现时间7.54~24.36 Ma,种内多样化增加时间1.74~0.86 Ma [51 ] Juniperus microsperma 西藏波密 早中新世21.6~14 Ma期间与其他种群分化 [52 ] Rhododendron 北半球 横断山区概属多样性在晚中新世(约8 Ma)快速增加 [53 ] Roscoea, Cautleya 喜马拉雅-横断山区 在早渐新世(~32 Ma)两属出现,Roscoea在横断山区与喜马拉雅山脉的两个分支大概在渐新世-中新世之交分化 [54 ] Nanorana parkeri 青藏高原南部 高原东部和西部类群分化时间1.4~3.7 Ma [55 ] Paini 南亚、东南亚、青藏高原 在渐新世早期棘蛙族高原-喜马拉雅类群与中国第二阶梯上生活的类群分化,随后在在26.1~12.5 Ma高原生活类群与喜马拉雅山脉生活类群分化,高原内部生活类群在晚中新世到上新世期间分化速率加快 [22 ] Bufo tibetanus 青藏高原、四川、甘肃、陕西等 高原种类形成于上新世期间 [56 ] Phrynocephalus 亚洲干旱区 在7.2~13.0 Ma期间演化出胎生类群,3.2~7.3 Ma期间种间多样性增加 [57 ] Phrynocephalus erythrurus 羌塘盆地 在3.67 Ma左右北部类群与南部类群发生分化,约2.76 Ma北部类群内部进一步分化 [58 ] Gnaptorina 青藏高原及周边高海拔地区 始新世起源于青藏高原东南缘 [18 -19 ] Coelotinae 北半球温带、亚热带 晚始新世高原东南缘隆升阻碍南方隙蛛向北扩散,渐新世高原中部和北部隆起导致北方隙蛛早期多样化进程中的灭绝事件 [59 ] Montifringilla, Onychostruthus, Pyrgilauda 青藏高原 雪雀的共同祖先在约14 Ma就应经生活在青藏高原,并进化出适应高海拔极端环境的性状 [21 ] Parus major 亚洲、欧洲 蒙古分布类群与其他种类分化时间为2.8~0.7 Ma,喜马拉雅山脉东部与中国中东部类群分化时间为1.9~0.4 Ma [60 ] Pseudopodoces humilis 青藏高原、柴达木盆地周边高山 在9.9~7.7 Ma期间地山雀就与与其他山雀分化开来,并出现适应高海拔性状 [61 ] Phylloscopus, Seicercus 亚洲 喜马拉雅山脉分布类群在7 Ma加速进化 [62 -63 ] Perdix 西藏、甘肃、青海、四川、新疆 高原类群出现于3.63 (4.52~2.75) Ma [64 ] Pantholops hodgsonii 西藏、新疆、青海 上新世-更新世就已经出现在青藏高原 [65 ] Procapra picticaudata 西藏、四川、甘肃、青海、新疆 种内分化时间约2.2~4.4 Ma [66 ] Rhinopithecus 四川、甘肃、陕西、贵州、云南、缅甸、越南 北方类群川金丝猴、黔金丝猴与喜马拉雅类群滇金丝猴、缅甸金丝猴分化时间约1.6 Ma [67 ] Ochotona 北半球 分布于青藏高原的高山类群和灌丛草原类群与低海拔泛北极分布类群分化时间大约为2.4 Ma [68 ] Apodemus draco 四川、云南、甘肃、青海、陕西、湖北、湖南、福建、台湾 2.2 Ma (3.8~0.87 Ma)分化形成,后来由于高原的阶段性隆升以及冰川作用种系及遗传结构更加复杂 [69 -70 ] Eothenomys 东亚 高原种类与低海拔种类分裂时间约2.7 Ma [71 ] Schizothoracinae 青藏高原及周边地区 在晚中新世晚期或上新世裂腹鱼亚科鱼类就已经出现在青藏高原及周边水系中,在晚更新世期间种内分化加速,不同海拔之间的类群产生等级分化,种间多样性增加,第四纪以来青藏高原隆升导致的水系改组是推动裂腹鱼亚科鱼类演化的原动力 [31 ,76 -78 ]
3 讨论 最近,青藏高原古地貌重建工作取得了丰硕的成果(图3 ),但是各种指标重建结果往往存在矛盾(图4 )[25 ,79 -83 ] .同位素重建结果表明高原南部在古近纪早期就存在安第斯型的巨大山系[84 -85 ] ;古生物研究发现高原腹地从中始新世到晚渐新世都存在东西向的大峡谷,该峡谷底部海拔在2 000 m左右,两侧为高大的冈底斯山脉和羌塘山脉[82 ,86 -88 ] ,动植物如巨犀(Paraceratherium )[89 ] 、攀鲈(Eoanabas )[90 -91 ] 、臭椿(Ailanthus )[88 ] 、椿榆(Cedrelospermum )[92 ] 、兔耳果(Lagokarpos )[93 ] 等在古近纪都可以通过青藏地区峡谷与周边动植物区系交流,新近纪之前的青藏地区虽然地形复杂,但是尚未隆升成今天的高原;横断山区某些区域则在晚始新世-早渐新世之交就已经获得现今的海拔高度[94 -95 ] ;高原北部和喜马拉雅山脉则在新近纪形成现在的地貌格局[25 ,96 -97 ] .古生物研究也发现青藏高原在晚上新世就已经存在高寒生境和冰缘环境,成为第四纪冰期动物忍耐寒冷环境的“训练营”,第四纪冰期来临之后这些适应寒冷环境的生物才扩散至北半球高纬度地区[34 ,98 -99 ] .因此,高原及周边地区高寒生物类群的起源时间表明在渐新世青藏高原部分区域就已经出现高寒生态系统,晚中新世以来青藏高原地貌与气候继续发生剧烈变化,促使这些物种的祖先类群迁入或就地演化积累适应高寒环境的基因和形态,并最终演化成适应高寒生境的新物种[25 ] .高寒区物种在晚上新世的加速分化可能与高原巨大山系和水系、冰川活动等造成的地理隔离有关[26 ] .随着第四纪冰期的到来,青藏高原及周边地区的高寒生物区也开始与北半球高纬度地区存在密切联系[34 ,98 -99 ] .同时,演化生物学研究也发现在第四纪冰期期间并未形成覆盖整个青藏高原的大冰盖,高原面上仍存在很多生物避难所[26 ] . ...
The roles of time and ecology in the continental radiation of the Old World leaf warblers (Phylloscopus and Seicercus )
2
2010
... 鸟类中的雪雀(Montifringilla , Onychostruthus , Pyrgilauda )是在青藏高原低温、缺氧、强紫外线极端环境下繁衍壮大的类群,其全基因组测序与蛋白质功能研究发现雪雀的共同祖先在约14 Ma就应经生活在青藏高原,并进化出适应高海拔极端环境的性状[21 ] .大山雀(Parus major )在北半球广泛分布,第四纪以来高原周边地区构造隆升以及全球气候变干等形成地理隔离,使得高原分布类群与周边地区分布的姐妹类群发生分化,多样性升高[60 ] .地山雀(Pseudopodoces humilis )在青藏高原分布的海拔较高,一般超过4 000 m,系统发育研究发现在9.9~7.7 Ma期间地山雀就与其他山雀分化开来,并进化出适应高海拔生境的形态和习性[61 ] .对喜马拉雅山脉鸣禽的研究表明,山脉隆升和季风演化等原因导致柳莺属(Phylloscopus )和鹟莺属(Seicercus )在7 Ma左右开始加速分化[62 -63 ] .山鹑属(Perdix )广泛分布于中国西北和西南地区,其中高原山鹑(P. hodgsoniae )分布于青藏高原海拔2 500~5 000 m范围内,系统发育研究结果表明其适应高原生境的类群出现于4.52~2.75 Ma期间[64 ] . ...
... Biological evolution in the alpine region of Tibetan Plateau
Table 1 种属 分布区域 适应高海拔类群分化时间或多样性增加时间 文献来源 Sinopodophyllum hexandrum 青藏高原东部 在约(6.52±1.89) Ma期间出现在青藏高原东部,但是直到3~2 Ma以来才演化出适应高山环境的形态 [47 ] Nannoglottis 青藏高原 在23~32 Ma期间出现在青藏高原,随后多样性增高,其高山灌丛和针叶林生活类型分支时间约为3.4 Ma [48 ] Reaumuria soongarica 西北干旱区、青藏高原北部 高原内部分布类群演化时间2.0~6.3 Ma,高原东北缘与高原西北类群分化时间1.1~3.5 Ma [49 ] Orchidinae 青藏高原及喜马拉雅山脉 青藏高原特有种大多是在过去6 Ma里进化而来 [50 ] Oxyria sinensis 青藏高原东南部 高山类群出现时间7.54~24.36 Ma,种内多样化增加时间1.74~0.86 Ma [51 ] Juniperus microsperma 西藏波密 早中新世21.6~14 Ma期间与其他种群分化 [52 ] Rhododendron 北半球 横断山区概属多样性在晚中新世(约8 Ma)快速增加 [53 ] Roscoea, Cautleya 喜马拉雅-横断山区 在早渐新世(~32 Ma)两属出现,Roscoea在横断山区与喜马拉雅山脉的两个分支大概在渐新世-中新世之交分化 [54 ] Nanorana parkeri 青藏高原南部 高原东部和西部类群分化时间1.4~3.7 Ma [55 ] Paini 南亚、东南亚、青藏高原 在渐新世早期棘蛙族高原-喜马拉雅类群与中国第二阶梯上生活的类群分化,随后在在26.1~12.5 Ma高原生活类群与喜马拉雅山脉生活类群分化,高原内部生活类群在晚中新世到上新世期间分化速率加快 [22 ] Bufo tibetanus 青藏高原、四川、甘肃、陕西等 高原种类形成于上新世期间 [56 ] Phrynocephalus 亚洲干旱区 在7.2~13.0 Ma期间演化出胎生类群,3.2~7.3 Ma期间种间多样性增加 [57 ] Phrynocephalus erythrurus 羌塘盆地 在3.67 Ma左右北部类群与南部类群发生分化,约2.76 Ma北部类群内部进一步分化 [58 ] Gnaptorina 青藏高原及周边高海拔地区 始新世起源于青藏高原东南缘 [18 -19 ] Coelotinae 北半球温带、亚热带 晚始新世高原东南缘隆升阻碍南方隙蛛向北扩散,渐新世高原中部和北部隆起导致北方隙蛛早期多样化进程中的灭绝事件 [59 ] Montifringilla, Onychostruthus, Pyrgilauda 青藏高原 雪雀的共同祖先在约14 Ma就应经生活在青藏高原,并进化出适应高海拔极端环境的性状 [21 ] Parus major 亚洲、欧洲 蒙古分布类群与其他种类分化时间为2.8~0.7 Ma,喜马拉雅山脉东部与中国中东部类群分化时间为1.9~0.4 Ma [60 ] Pseudopodoces humilis 青藏高原、柴达木盆地周边高山 在9.9~7.7 Ma期间地山雀就与与其他山雀分化开来,并出现适应高海拔性状 [61 ] Phylloscopus, Seicercus 亚洲 喜马拉雅山脉分布类群在7 Ma加速进化 [62 -63 ] Perdix 西藏、甘肃、青海、四川、新疆 高原类群出现于3.63 (4.52~2.75) Ma [64 ] Pantholops hodgsonii 西藏、新疆、青海 上新世-更新世就已经出现在青藏高原 [65 ] Procapra picticaudata 西藏、四川、甘肃、青海、新疆 种内分化时间约2.2~4.4 Ma [66 ] Rhinopithecus 四川、甘肃、陕西、贵州、云南、缅甸、越南 北方类群川金丝猴、黔金丝猴与喜马拉雅类群滇金丝猴、缅甸金丝猴分化时间约1.6 Ma [67 ] Ochotona 北半球 分布于青藏高原的高山类群和灌丛草原类群与低海拔泛北极分布类群分化时间大约为2.4 Ma [68 ] Apodemus draco 四川、云南、甘肃、青海、陕西、湖北、湖南、福建、台湾 2.2 Ma (3.8~0.87 Ma)分化形成,后来由于高原的阶段性隆升以及冰川作用种系及遗传结构更加复杂 [69 -70 ] Eothenomys 东亚 高原种类与低海拔种类分裂时间约2.7 Ma [71 ] Schizothoracinae 青藏高原及周边地区 在晚中新世晚期或上新世裂腹鱼亚科鱼类就已经出现在青藏高原及周边水系中,在晚更新世期间种内分化加速,不同海拔之间的类群产生等级分化,种间多样性增加,第四纪以来青藏高原隆升导致的水系改组是推动裂腹鱼亚科鱼类演化的原动力 [31 ,76 -78 ]
3 讨论 最近,青藏高原古地貌重建工作取得了丰硕的成果(图3 ),但是各种指标重建结果往往存在矛盾(图4 )[25 ,79 -83 ] .同位素重建结果表明高原南部在古近纪早期就存在安第斯型的巨大山系[84 -85 ] ;古生物研究发现高原腹地从中始新世到晚渐新世都存在东西向的大峡谷,该峡谷底部海拔在2 000 m左右,两侧为高大的冈底斯山脉和羌塘山脉[82 ,86 -88 ] ,动植物如巨犀(Paraceratherium )[89 ] 、攀鲈(Eoanabas )[90 -91 ] 、臭椿(Ailanthus )[88 ] 、椿榆(Cedrelospermum )[92 ] 、兔耳果(Lagokarpos )[93 ] 等在古近纪都可以通过青藏地区峡谷与周边动植物区系交流,新近纪之前的青藏地区虽然地形复杂,但是尚未隆升成今天的高原;横断山区某些区域则在晚始新世-早渐新世之交就已经获得现今的海拔高度[94 -95 ] ;高原北部和喜马拉雅山脉则在新近纪形成现在的地貌格局[25 ,96 -97 ] .古生物研究也发现青藏高原在晚上新世就已经存在高寒生境和冰缘环境,成为第四纪冰期动物忍耐寒冷环境的“训练营”,第四纪冰期来临之后这些适应寒冷环境的生物才扩散至北半球高纬度地区[34 ,98 -99 ] .因此,高原及周边地区高寒生物类群的起源时间表明在渐新世青藏高原部分区域就已经出现高寒生态系统,晚中新世以来青藏高原地貌与气候继续发生剧烈变化,促使这些物种的祖先类群迁入或就地演化积累适应高寒环境的基因和形态,并最终演化成适应高寒生境的新物种[25 ] .高寒区物种在晚上新世的加速分化可能与高原巨大山系和水系、冰川活动等造成的地理隔离有关[26 ] .随着第四纪冰期的到来,青藏高原及周边地区的高寒生物区也开始与北半球高纬度地区存在密切联系[34 ,98 -99 ] .同时,演化生物学研究也发现在第四纪冰期期间并未形成覆盖整个青藏高原的大冰盖,高原面上仍存在很多生物避难所[26 ] . ...
Niche filling slows the diversification of Himalayan songbirds
2
2014
... 鸟类中的雪雀(Montifringilla , Onychostruthus , Pyrgilauda )是在青藏高原低温、缺氧、强紫外线极端环境下繁衍壮大的类群,其全基因组测序与蛋白质功能研究发现雪雀的共同祖先在约14 Ma就应经生活在青藏高原,并进化出适应高海拔极端环境的性状[21 ] .大山雀(Parus major )在北半球广泛分布,第四纪以来高原周边地区构造隆升以及全球气候变干等形成地理隔离,使得高原分布类群与周边地区分布的姐妹类群发生分化,多样性升高[60 ] .地山雀(Pseudopodoces humilis )在青藏高原分布的海拔较高,一般超过4 000 m,系统发育研究发现在9.9~7.7 Ma期间地山雀就与其他山雀分化开来,并进化出适应高海拔生境的形态和习性[61 ] .对喜马拉雅山脉鸣禽的研究表明,山脉隆升和季风演化等原因导致柳莺属(Phylloscopus )和鹟莺属(Seicercus )在7 Ma左右开始加速分化[62 -63 ] .山鹑属(Perdix )广泛分布于中国西北和西南地区,其中高原山鹑(P. hodgsoniae )分布于青藏高原海拔2 500~5 000 m范围内,系统发育研究结果表明其适应高原生境的类群出现于4.52~2.75 Ma期间[64 ] . ...
... Biological evolution in the alpine region of Tibetan Plateau
Table 1 种属 分布区域 适应高海拔类群分化时间或多样性增加时间 文献来源 Sinopodophyllum hexandrum 青藏高原东部 在约(6.52±1.89) Ma期间出现在青藏高原东部,但是直到3~2 Ma以来才演化出适应高山环境的形态 [47 ] Nannoglottis 青藏高原 在23~32 Ma期间出现在青藏高原,随后多样性增高,其高山灌丛和针叶林生活类型分支时间约为3.4 Ma [48 ] Reaumuria soongarica 西北干旱区、青藏高原北部 高原内部分布类群演化时间2.0~6.3 Ma,高原东北缘与高原西北类群分化时间1.1~3.5 Ma [49 ] Orchidinae 青藏高原及喜马拉雅山脉 青藏高原特有种大多是在过去6 Ma里进化而来 [50 ] Oxyria sinensis 青藏高原东南部 高山类群出现时间7.54~24.36 Ma,种内多样化增加时间1.74~0.86 Ma [51 ] Juniperus microsperma 西藏波密 早中新世21.6~14 Ma期间与其他种群分化 [52 ] Rhododendron 北半球 横断山区概属多样性在晚中新世(约8 Ma)快速增加 [53 ] Roscoea, Cautleya 喜马拉雅-横断山区 在早渐新世(~32 Ma)两属出现,Roscoea在横断山区与喜马拉雅山脉的两个分支大概在渐新世-中新世之交分化 [54 ] Nanorana parkeri 青藏高原南部 高原东部和西部类群分化时间1.4~3.7 Ma [55 ] Paini 南亚、东南亚、青藏高原 在渐新世早期棘蛙族高原-喜马拉雅类群与中国第二阶梯上生活的类群分化,随后在在26.1~12.5 Ma高原生活类群与喜马拉雅山脉生活类群分化,高原内部生活类群在晚中新世到上新世期间分化速率加快 [22 ] Bufo tibetanus 青藏高原、四川、甘肃、陕西等 高原种类形成于上新世期间 [56 ] Phrynocephalus 亚洲干旱区 在7.2~13.0 Ma期间演化出胎生类群,3.2~7.3 Ma期间种间多样性增加 [57 ] Phrynocephalus erythrurus 羌塘盆地 在3.67 Ma左右北部类群与南部类群发生分化,约2.76 Ma北部类群内部进一步分化 [58 ] Gnaptorina 青藏高原及周边高海拔地区 始新世起源于青藏高原东南缘 [18 -19 ] Coelotinae 北半球温带、亚热带 晚始新世高原东南缘隆升阻碍南方隙蛛向北扩散,渐新世高原中部和北部隆起导致北方隙蛛早期多样化进程中的灭绝事件 [59 ] Montifringilla, Onychostruthus, Pyrgilauda 青藏高原 雪雀的共同祖先在约14 Ma就应经生活在青藏高原,并进化出适应高海拔极端环境的性状 [21 ] Parus major 亚洲、欧洲 蒙古分布类群与其他种类分化时间为2.8~0.7 Ma,喜马拉雅山脉东部与中国中东部类群分化时间为1.9~0.4 Ma [60 ] Pseudopodoces humilis 青藏高原、柴达木盆地周边高山 在9.9~7.7 Ma期间地山雀就与与其他山雀分化开来,并出现适应高海拔性状 [61 ] Phylloscopus, Seicercus 亚洲 喜马拉雅山脉分布类群在7 Ma加速进化 [62 -63 ] Perdix 西藏、甘肃、青海、四川、新疆 高原类群出现于3.63 (4.52~2.75) Ma [64 ] Pantholops hodgsonii 西藏、新疆、青海 上新世-更新世就已经出现在青藏高原 [65 ] Procapra picticaudata 西藏、四川、甘肃、青海、新疆 种内分化时间约2.2~4.4 Ma [66 ] Rhinopithecus 四川、甘肃、陕西、贵州、云南、缅甸、越南 北方类群川金丝猴、黔金丝猴与喜马拉雅类群滇金丝猴、缅甸金丝猴分化时间约1.6 Ma [67 ] Ochotona 北半球 分布于青藏高原的高山类群和灌丛草原类群与低海拔泛北极分布类群分化时间大约为2.4 Ma [68 ] Apodemus draco 四川、云南、甘肃、青海、陕西、湖北、湖南、福建、台湾 2.2 Ma (3.8~0.87 Ma)分化形成,后来由于高原的阶段性隆升以及冰川作用种系及遗传结构更加复杂 [69 -70 ] Eothenomys 东亚 高原种类与低海拔种类分裂时间约2.7 Ma [71 ] Schizothoracinae 青藏高原及周边地区 在晚中新世晚期或上新世裂腹鱼亚科鱼类就已经出现在青藏高原及周边水系中,在晚更新世期间种内分化加速,不同海拔之间的类群产生等级分化,种间多样性增加,第四纪以来青藏高原隆升导致的水系改组是推动裂腹鱼亚科鱼类演化的原动力 [31 ,76 -78 ]
3 讨论 最近,青藏高原古地貌重建工作取得了丰硕的成果(图3 ),但是各种指标重建结果往往存在矛盾(图4 )[25 ,79 -83 ] .同位素重建结果表明高原南部在古近纪早期就存在安第斯型的巨大山系[84 -85 ] ;古生物研究发现高原腹地从中始新世到晚渐新世都存在东西向的大峡谷,该峡谷底部海拔在2 000 m左右,两侧为高大的冈底斯山脉和羌塘山脉[82 ,86 -88 ] ,动植物如巨犀(Paraceratherium )[89 ] 、攀鲈(Eoanabas )[90 -91 ] 、臭椿(Ailanthus )[88 ] 、椿榆(Cedrelospermum )[92 ] 、兔耳果(Lagokarpos )[93 ] 等在古近纪都可以通过青藏地区峡谷与周边动植物区系交流,新近纪之前的青藏地区虽然地形复杂,但是尚未隆升成今天的高原;横断山区某些区域则在晚始新世-早渐新世之交就已经获得现今的海拔高度[94 -95 ] ;高原北部和喜马拉雅山脉则在新近纪形成现在的地貌格局[25 ,96 -97 ] .古生物研究也发现青藏高原在晚上新世就已经存在高寒生境和冰缘环境,成为第四纪冰期动物忍耐寒冷环境的“训练营”,第四纪冰期来临之后这些适应寒冷环境的生物才扩散至北半球高纬度地区[34 ,98 -99 ] .因此,高原及周边地区高寒生物类群的起源时间表明在渐新世青藏高原部分区域就已经出现高寒生态系统,晚中新世以来青藏高原地貌与气候继续发生剧烈变化,促使这些物种的祖先类群迁入或就地演化积累适应高寒环境的基因和形态,并最终演化成适应高寒生境的新物种[25 ] .高寒区物种在晚上新世的加速分化可能与高原巨大山系和水系、冰川活动等造成的地理隔离有关[26 ] .随着第四纪冰期的到来,青藏高原及周边地区的高寒生物区也开始与北半球高纬度地区存在密切联系[34 ,98 -99 ] .同时,演化生物学研究也发现在第四纪冰期期间并未形成覆盖整个青藏高原的大冰盖,高原面上仍存在很多生物避难所[26 ] . ...
The phylogenetic position and speciation dynamics of the genus Perdix (Phasianidae, Galliformes)
2
2010
... 鸟类中的雪雀(Montifringilla , Onychostruthus , Pyrgilauda )是在青藏高原低温、缺氧、强紫外线极端环境下繁衍壮大的类群,其全基因组测序与蛋白质功能研究发现雪雀的共同祖先在约14 Ma就应经生活在青藏高原,并进化出适应高海拔极端环境的性状[21 ] .大山雀(Parus major )在北半球广泛分布,第四纪以来高原周边地区构造隆升以及全球气候变干等形成地理隔离,使得高原分布类群与周边地区分布的姐妹类群发生分化,多样性升高[60 ] .地山雀(Pseudopodoces humilis )在青藏高原分布的海拔较高,一般超过4 000 m,系统发育研究发现在9.9~7.7 Ma期间地山雀就与其他山雀分化开来,并进化出适应高海拔生境的形态和习性[61 ] .对喜马拉雅山脉鸣禽的研究表明,山脉隆升和季风演化等原因导致柳莺属(Phylloscopus )和鹟莺属(Seicercus )在7 Ma左右开始加速分化[62 -63 ] .山鹑属(Perdix )广泛分布于中国西北和西南地区,其中高原山鹑(P. hodgsoniae )分布于青藏高原海拔2 500~5 000 m范围内,系统发育研究结果表明其适应高原生境的类群出现于4.52~2.75 Ma期间[64 ] . ...
... Biological evolution in the alpine region of Tibetan Plateau
Table 1 种属 分布区域 适应高海拔类群分化时间或多样性增加时间 文献来源 Sinopodophyllum hexandrum 青藏高原东部 在约(6.52±1.89) Ma期间出现在青藏高原东部,但是直到3~2 Ma以来才演化出适应高山环境的形态 [47 ] Nannoglottis 青藏高原 在23~32 Ma期间出现在青藏高原,随后多样性增高,其高山灌丛和针叶林生活类型分支时间约为3.4 Ma [48 ] Reaumuria soongarica 西北干旱区、青藏高原北部 高原内部分布类群演化时间2.0~6.3 Ma,高原东北缘与高原西北类群分化时间1.1~3.5 Ma [49 ] Orchidinae 青藏高原及喜马拉雅山脉 青藏高原特有种大多是在过去6 Ma里进化而来 [50 ] Oxyria sinensis 青藏高原东南部 高山类群出现时间7.54~24.36 Ma,种内多样化增加时间1.74~0.86 Ma [51 ] Juniperus microsperma 西藏波密 早中新世21.6~14 Ma期间与其他种群分化 [52 ] Rhododendron 北半球 横断山区概属多样性在晚中新世(约8 Ma)快速增加 [53 ] Roscoea, Cautleya 喜马拉雅-横断山区 在早渐新世(~32 Ma)两属出现,Roscoea在横断山区与喜马拉雅山脉的两个分支大概在渐新世-中新世之交分化 [54 ] Nanorana parkeri 青藏高原南部 高原东部和西部类群分化时间1.4~3.7 Ma [55 ] Paini 南亚、东南亚、青藏高原 在渐新世早期棘蛙族高原-喜马拉雅类群与中国第二阶梯上生活的类群分化,随后在在26.1~12.5 Ma高原生活类群与喜马拉雅山脉生活类群分化,高原内部生活类群在晚中新世到上新世期间分化速率加快 [22 ] Bufo tibetanus 青藏高原、四川、甘肃、陕西等 高原种类形成于上新世期间 [56 ] Phrynocephalus 亚洲干旱区 在7.2~13.0 Ma期间演化出胎生类群,3.2~7.3 Ma期间种间多样性增加 [57 ] Phrynocephalus erythrurus 羌塘盆地 在3.67 Ma左右北部类群与南部类群发生分化,约2.76 Ma北部类群内部进一步分化 [58 ] Gnaptorina 青藏高原及周边高海拔地区 始新世起源于青藏高原东南缘 [18 -19 ] Coelotinae 北半球温带、亚热带 晚始新世高原东南缘隆升阻碍南方隙蛛向北扩散,渐新世高原中部和北部隆起导致北方隙蛛早期多样化进程中的灭绝事件 [59 ] Montifringilla, Onychostruthus, Pyrgilauda 青藏高原 雪雀的共同祖先在约14 Ma就应经生活在青藏高原,并进化出适应高海拔极端环境的性状 [21 ] Parus major 亚洲、欧洲 蒙古分布类群与其他种类分化时间为2.8~0.7 Ma,喜马拉雅山脉东部与中国中东部类群分化时间为1.9~0.4 Ma [60 ] Pseudopodoces humilis 青藏高原、柴达木盆地周边高山 在9.9~7.7 Ma期间地山雀就与与其他山雀分化开来,并出现适应高海拔性状 [61 ] Phylloscopus, Seicercus 亚洲 喜马拉雅山脉分布类群在7 Ma加速进化 [62 -63 ] Perdix 西藏、甘肃、青海、四川、新疆 高原类群出现于3.63 (4.52~2.75) Ma [64 ] Pantholops hodgsonii 西藏、新疆、青海 上新世-更新世就已经出现在青藏高原 [65 ] Procapra picticaudata 西藏、四川、甘肃、青海、新疆 种内分化时间约2.2~4.4 Ma [66 ] Rhinopithecus 四川、甘肃、陕西、贵州、云南、缅甸、越南 北方类群川金丝猴、黔金丝猴与喜马拉雅类群滇金丝猴、缅甸金丝猴分化时间约1.6 Ma [67 ] Ochotona 北半球 分布于青藏高原的高山类群和灌丛草原类群与低海拔泛北极分布类群分化时间大约为2.4 Ma [68 ] Apodemus draco 四川、云南、甘肃、青海、陕西、湖北、湖南、福建、台湾 2.2 Ma (3.8~0.87 Ma)分化形成,后来由于高原的阶段性隆升以及冰川作用种系及遗传结构更加复杂 [69 -70 ] Eothenomys 东亚 高原种类与低海拔种类分裂时间约2.7 Ma [71 ] Schizothoracinae 青藏高原及周边地区 在晚中新世晚期或上新世裂腹鱼亚科鱼类就已经出现在青藏高原及周边水系中,在晚更新世期间种内分化加速,不同海拔之间的类群产生等级分化,种间多样性增加,第四纪以来青藏高原隆升导致的水系改组是推动裂腹鱼亚科鱼类演化的原动力 [31 ,76 -78 ]
3 讨论 最近,青藏高原古地貌重建工作取得了丰硕的成果(图3 ),但是各种指标重建结果往往存在矛盾(图4 )[25 ,79 -83 ] .同位素重建结果表明高原南部在古近纪早期就存在安第斯型的巨大山系[84 -85 ] ;古生物研究发现高原腹地从中始新世到晚渐新世都存在东西向的大峡谷,该峡谷底部海拔在2 000 m左右,两侧为高大的冈底斯山脉和羌塘山脉[82 ,86 -88 ] ,动植物如巨犀(Paraceratherium )[89 ] 、攀鲈(Eoanabas )[90 -91 ] 、臭椿(Ailanthus )[88 ] 、椿榆(Cedrelospermum )[92 ] 、兔耳果(Lagokarpos )[93 ] 等在古近纪都可以通过青藏地区峡谷与周边动植物区系交流,新近纪之前的青藏地区虽然地形复杂,但是尚未隆升成今天的高原;横断山区某些区域则在晚始新世-早渐新世之交就已经获得现今的海拔高度[94 -95 ] ;高原北部和喜马拉雅山脉则在新近纪形成现在的地貌格局[25 ,96 -97 ] .古生物研究也发现青藏高原在晚上新世就已经存在高寒生境和冰缘环境,成为第四纪冰期动物忍耐寒冷环境的“训练营”,第四纪冰期来临之后这些适应寒冷环境的生物才扩散至北半球高纬度地区[34 ,98 -99 ] .因此,高原及周边地区高寒生物类群的起源时间表明在渐新世青藏高原部分区域就已经出现高寒生态系统,晚中新世以来青藏高原地貌与气候继续发生剧烈变化,促使这些物种的祖先类群迁入或就地演化积累适应高寒环境的基因和形态,并最终演化成适应高寒生境的新物种[25 ] .高寒区物种在晚上新世的加速分化可能与高原巨大山系和水系、冰川活动等造成的地理隔离有关[26 ] .随着第四纪冰期的到来,青藏高原及周边地区的高寒生物区也开始与北半球高纬度地区存在密切联系[34 ,98 -99 ] .同时,演化生物学研究也发现在第四纪冰期期间并未形成覆盖整个青藏高原的大冰盖,高原面上仍存在很多生物避难所[26 ] . ...
Evolutionary history and current population relationships of the Chiru (Pantholops hodgsonii ) inferred from mtDNA variation
2
2005
... 哺乳动物中的藏羚羊(Pantholops hodgsonii )生活于西藏、青海和新疆等地海拔3 700~5 500 m的区域,能够成功适应高原缺氧环境,其系统发育学研究表明在上新世-早更新世期间该类群就已经生活在青藏高原地区[65 ] .藏原羚(Procapra picticaudata )现今生活于海拔3 000~5 100 m地区,其种内分化时间约为4.4~2.2 Ma[66 ] .灵长目仰鼻猴属(Rhinopithecus )鼻孔结构是对高原缺氧环境的适应,系统发育研究表明在1.6 Ma左右其分布于四川和云南的金丝猴北方类群与分布于喜马拉雅的金丝猴类群发生分化[67 ] .兔形目鼠兔属(Ochotona )广泛分布于北半球草原和灌丛,其分布于青藏高原的类群演化出适应高寒生境的习性,在2.4 Ma左右,适应高寒生境类群与低海拔泛北极分布类群分化[68 ] .中华姬鼠(Apodemus draco )分布于青藏高原及周边地区以及湖南、湖北和台湾等地,其在3.8~0.87 Ma期间分化形成适应高海拔生境的物种,其种系及遗传结构由于高原阶段性隆升和冰川活动的影响而变得更加复杂[69 -70 ] .绒鼠属(Eothenomys )高原类群与低海拔类群分化时间约为2.7 Ma[71 ] .基于4 966种现生陆栖脊椎动物和1 278个哺乳动物化石属的研究也表明受印度-欧亚板块碰撞、高原阶段性隆升、亚洲内陆干旱化等事件的影响,青藏高原及其邻近地区动物区系在新生代发生显著变化,其区系成分在早期更接近东洋界,并最终在中新世-上新世之交形成与现在接近的动物区系格局[72 ] .对174种青藏高原特有陆栖脊椎动物祖先分布区、来源和分化时间的估算,发现青藏高原特有种早在55 Ma就开始从周边区域扩散至高原,扩散事件约从15 Ma开始增加,并在6 Ma之后达到顶峰,高原隆升和历史气候变化对该地区动物区系的形成与演化具有重要作用[73 ] . ...
... Biological evolution in the alpine region of Tibetan Plateau
Table 1 种属 分布区域 适应高海拔类群分化时间或多样性增加时间 文献来源 Sinopodophyllum hexandrum 青藏高原东部 在约(6.52±1.89) Ma期间出现在青藏高原东部,但是直到3~2 Ma以来才演化出适应高山环境的形态 [47 ] Nannoglottis 青藏高原 在23~32 Ma期间出现在青藏高原,随后多样性增高,其高山灌丛和针叶林生活类型分支时间约为3.4 Ma [48 ] Reaumuria soongarica 西北干旱区、青藏高原北部 高原内部分布类群演化时间2.0~6.3 Ma,高原东北缘与高原西北类群分化时间1.1~3.5 Ma [49 ] Orchidinae 青藏高原及喜马拉雅山脉 青藏高原特有种大多是在过去6 Ma里进化而来 [50 ] Oxyria sinensis 青藏高原东南部 高山类群出现时间7.54~24.36 Ma,种内多样化增加时间1.74~0.86 Ma [51 ] Juniperus microsperma 西藏波密 早中新世21.6~14 Ma期间与其他种群分化 [52 ] Rhododendron 北半球 横断山区概属多样性在晚中新世(约8 Ma)快速增加 [53 ] Roscoea, Cautleya 喜马拉雅-横断山区 在早渐新世(~32 Ma)两属出现,Roscoea在横断山区与喜马拉雅山脉的两个分支大概在渐新世-中新世之交分化 [54 ] Nanorana parkeri 青藏高原南部 高原东部和西部类群分化时间1.4~3.7 Ma [55 ] Paini 南亚、东南亚、青藏高原 在渐新世早期棘蛙族高原-喜马拉雅类群与中国第二阶梯上生活的类群分化,随后在在26.1~12.5 Ma高原生活类群与喜马拉雅山脉生活类群分化,高原内部生活类群在晚中新世到上新世期间分化速率加快 [22 ] Bufo tibetanus 青藏高原、四川、甘肃、陕西等 高原种类形成于上新世期间 [56 ] Phrynocephalus 亚洲干旱区 在7.2~13.0 Ma期间演化出胎生类群,3.2~7.3 Ma期间种间多样性增加 [57 ] Phrynocephalus erythrurus 羌塘盆地 在3.67 Ma左右北部类群与南部类群发生分化,约2.76 Ma北部类群内部进一步分化 [58 ] Gnaptorina 青藏高原及周边高海拔地区 始新世起源于青藏高原东南缘 [18 -19 ] Coelotinae 北半球温带、亚热带 晚始新世高原东南缘隆升阻碍南方隙蛛向北扩散,渐新世高原中部和北部隆起导致北方隙蛛早期多样化进程中的灭绝事件 [59 ] Montifringilla, Onychostruthus, Pyrgilauda 青藏高原 雪雀的共同祖先在约14 Ma就应经生活在青藏高原,并进化出适应高海拔极端环境的性状 [21 ] Parus major 亚洲、欧洲 蒙古分布类群与其他种类分化时间为2.8~0.7 Ma,喜马拉雅山脉东部与中国中东部类群分化时间为1.9~0.4 Ma [60 ] Pseudopodoces humilis 青藏高原、柴达木盆地周边高山 在9.9~7.7 Ma期间地山雀就与与其他山雀分化开来,并出现适应高海拔性状 [61 ] Phylloscopus, Seicercus 亚洲 喜马拉雅山脉分布类群在7 Ma加速进化 [62 -63 ] Perdix 西藏、甘肃、青海、四川、新疆 高原类群出现于3.63 (4.52~2.75) Ma [64 ] Pantholops hodgsonii 西藏、新疆、青海 上新世-更新世就已经出现在青藏高原 [65 ] Procapra picticaudata 西藏、四川、甘肃、青海、新疆 种内分化时间约2.2~4.4 Ma [66 ] Rhinopithecus 四川、甘肃、陕西、贵州、云南、缅甸、越南 北方类群川金丝猴、黔金丝猴与喜马拉雅类群滇金丝猴、缅甸金丝猴分化时间约1.6 Ma [67 ] Ochotona 北半球 分布于青藏高原的高山类群和灌丛草原类群与低海拔泛北极分布类群分化时间大约为2.4 Ma [68 ] Apodemus draco 四川、云南、甘肃、青海、陕西、湖北、湖南、福建、台湾 2.2 Ma (3.8~0.87 Ma)分化形成,后来由于高原的阶段性隆升以及冰川作用种系及遗传结构更加复杂 [69 -70 ] Eothenomys 东亚 高原种类与低海拔种类分裂时间约2.7 Ma [71 ] Schizothoracinae 青藏高原及周边地区 在晚中新世晚期或上新世裂腹鱼亚科鱼类就已经出现在青藏高原及周边水系中,在晚更新世期间种内分化加速,不同海拔之间的类群产生等级分化,种间多样性增加,第四纪以来青藏高原隆升导致的水系改组是推动裂腹鱼亚科鱼类演化的原动力 [31 ,76 -78 ]
3 讨论 最近,青藏高原古地貌重建工作取得了丰硕的成果(图3 ),但是各种指标重建结果往往存在矛盾(图4 )[25 ,79 -83 ] .同位素重建结果表明高原南部在古近纪早期就存在安第斯型的巨大山系[84 -85 ] ;古生物研究发现高原腹地从中始新世到晚渐新世都存在东西向的大峡谷,该峡谷底部海拔在2 000 m左右,两侧为高大的冈底斯山脉和羌塘山脉[82 ,86 -88 ] ,动植物如巨犀(Paraceratherium )[89 ] 、攀鲈(Eoanabas )[90 -91 ] 、臭椿(Ailanthus )[88 ] 、椿榆(Cedrelospermum )[92 ] 、兔耳果(Lagokarpos )[93 ] 等在古近纪都可以通过青藏地区峡谷与周边动植物区系交流,新近纪之前的青藏地区虽然地形复杂,但是尚未隆升成今天的高原;横断山区某些区域则在晚始新世-早渐新世之交就已经获得现今的海拔高度[94 -95 ] ;高原北部和喜马拉雅山脉则在新近纪形成现在的地貌格局[25 ,96 -97 ] .古生物研究也发现青藏高原在晚上新世就已经存在高寒生境和冰缘环境,成为第四纪冰期动物忍耐寒冷环境的“训练营”,第四纪冰期来临之后这些适应寒冷环境的生物才扩散至北半球高纬度地区[34 ,98 -99 ] .因此,高原及周边地区高寒生物类群的起源时间表明在渐新世青藏高原部分区域就已经出现高寒生态系统,晚中新世以来青藏高原地貌与气候继续发生剧烈变化,促使这些物种的祖先类群迁入或就地演化积累适应高寒环境的基因和形态,并最终演化成适应高寒生境的新物种[25 ] .高寒区物种在晚上新世的加速分化可能与高原巨大山系和水系、冰川活动等造成的地理隔离有关[26 ] .随着第四纪冰期的到来,青藏高原及周边地区的高寒生物区也开始与北半球高纬度地区存在密切联系[34 ,98 -99 ] .同时,演化生物学研究也发现在第四纪冰期期间并未形成覆盖整个青藏高原的大冰盖,高原面上仍存在很多生物避难所[26 ] . ...
Mitochondrial phylogeography and genetic diversity of Tibetan gazelle (Procapra picticaudata ): implications for conservation
2
2006
... 哺乳动物中的藏羚羊(Pantholops hodgsonii )生活于西藏、青海和新疆等地海拔3 700~5 500 m的区域,能够成功适应高原缺氧环境,其系统发育学研究表明在上新世-早更新世期间该类群就已经生活在青藏高原地区[65 ] .藏原羚(Procapra picticaudata )现今生活于海拔3 000~5 100 m地区,其种内分化时间约为4.4~2.2 Ma[66 ] .灵长目仰鼻猴属(Rhinopithecus )鼻孔结构是对高原缺氧环境的适应,系统发育研究表明在1.6 Ma左右其分布于四川和云南的金丝猴北方类群与分布于喜马拉雅的金丝猴类群发生分化[67 ] .兔形目鼠兔属(Ochotona )广泛分布于北半球草原和灌丛,其分布于青藏高原的类群演化出适应高寒生境的习性,在2.4 Ma左右,适应高寒生境类群与低海拔泛北极分布类群分化[68 ] .中华姬鼠(Apodemus draco )分布于青藏高原及周边地区以及湖南、湖北和台湾等地,其在3.8~0.87 Ma期间分化形成适应高海拔生境的物种,其种系及遗传结构由于高原阶段性隆升和冰川活动的影响而变得更加复杂[69 -70 ] .绒鼠属(Eothenomys )高原类群与低海拔类群分化时间约为2.7 Ma[71 ] .基于4 966种现生陆栖脊椎动物和1 278个哺乳动物化石属的研究也表明受印度-欧亚板块碰撞、高原阶段性隆升、亚洲内陆干旱化等事件的影响,青藏高原及其邻近地区动物区系在新生代发生显著变化,其区系成分在早期更接近东洋界,并最终在中新世-上新世之交形成与现在接近的动物区系格局[72 ] .对174种青藏高原特有陆栖脊椎动物祖先分布区、来源和分化时间的估算,发现青藏高原特有种早在55 Ma就开始从周边区域扩散至高原,扩散事件约从15 Ma开始增加,并在6 Ma之后达到顶峰,高原隆升和历史气候变化对该地区动物区系的形成与演化具有重要作用[73 ] . ...
... Biological evolution in the alpine region of Tibetan Plateau
Table 1 种属 分布区域 适应高海拔类群分化时间或多样性增加时间 文献来源 Sinopodophyllum hexandrum 青藏高原东部 在约(6.52±1.89) Ma期间出现在青藏高原东部,但是直到3~2 Ma以来才演化出适应高山环境的形态 [47 ] Nannoglottis 青藏高原 在23~32 Ma期间出现在青藏高原,随后多样性增高,其高山灌丛和针叶林生活类型分支时间约为3.4 Ma [48 ] Reaumuria soongarica 西北干旱区、青藏高原北部 高原内部分布类群演化时间2.0~6.3 Ma,高原东北缘与高原西北类群分化时间1.1~3.5 Ma [49 ] Orchidinae 青藏高原及喜马拉雅山脉 青藏高原特有种大多是在过去6 Ma里进化而来 [50 ] Oxyria sinensis 青藏高原东南部 高山类群出现时间7.54~24.36 Ma,种内多样化增加时间1.74~0.86 Ma [51 ] Juniperus microsperma 西藏波密 早中新世21.6~14 Ma期间与其他种群分化 [52 ] Rhododendron 北半球 横断山区概属多样性在晚中新世(约8 Ma)快速增加 [53 ] Roscoea, Cautleya 喜马拉雅-横断山区 在早渐新世(~32 Ma)两属出现,Roscoea在横断山区与喜马拉雅山脉的两个分支大概在渐新世-中新世之交分化 [54 ] Nanorana parkeri 青藏高原南部 高原东部和西部类群分化时间1.4~3.7 Ma [55 ] Paini 南亚、东南亚、青藏高原 在渐新世早期棘蛙族高原-喜马拉雅类群与中国第二阶梯上生活的类群分化,随后在在26.1~12.5 Ma高原生活类群与喜马拉雅山脉生活类群分化,高原内部生活类群在晚中新世到上新世期间分化速率加快 [22 ] Bufo tibetanus 青藏高原、四川、甘肃、陕西等 高原种类形成于上新世期间 [56 ] Phrynocephalus 亚洲干旱区 在7.2~13.0 Ma期间演化出胎生类群,3.2~7.3 Ma期间种间多样性增加 [57 ] Phrynocephalus erythrurus 羌塘盆地 在3.67 Ma左右北部类群与南部类群发生分化,约2.76 Ma北部类群内部进一步分化 [58 ] Gnaptorina 青藏高原及周边高海拔地区 始新世起源于青藏高原东南缘 [18 -19 ] Coelotinae 北半球温带、亚热带 晚始新世高原东南缘隆升阻碍南方隙蛛向北扩散,渐新世高原中部和北部隆起导致北方隙蛛早期多样化进程中的灭绝事件 [59 ] Montifringilla, Onychostruthus, Pyrgilauda 青藏高原 雪雀的共同祖先在约14 Ma就应经生活在青藏高原,并进化出适应高海拔极端环境的性状 [21 ] Parus major 亚洲、欧洲 蒙古分布类群与其他种类分化时间为2.8~0.7 Ma,喜马拉雅山脉东部与中国中东部类群分化时间为1.9~0.4 Ma [60 ] Pseudopodoces humilis 青藏高原、柴达木盆地周边高山 在9.9~7.7 Ma期间地山雀就与与其他山雀分化开来,并出现适应高海拔性状 [61 ] Phylloscopus, Seicercus 亚洲 喜马拉雅山脉分布类群在7 Ma加速进化 [62 -63 ] Perdix 西藏、甘肃、青海、四川、新疆 高原类群出现于3.63 (4.52~2.75) Ma [64 ] Pantholops hodgsonii 西藏、新疆、青海 上新世-更新世就已经出现在青藏高原 [65 ] Procapra picticaudata 西藏、四川、甘肃、青海、新疆 种内分化时间约2.2~4.4 Ma [66 ] Rhinopithecus 四川、甘肃、陕西、贵州、云南、缅甸、越南 北方类群川金丝猴、黔金丝猴与喜马拉雅类群滇金丝猴、缅甸金丝猴分化时间约1.6 Ma [67 ] Ochotona 北半球 分布于青藏高原的高山类群和灌丛草原类群与低海拔泛北极分布类群分化时间大约为2.4 Ma [68 ] Apodemus draco 四川、云南、甘肃、青海、陕西、湖北、湖南、福建、台湾 2.2 Ma (3.8~0.87 Ma)分化形成,后来由于高原的阶段性隆升以及冰川作用种系及遗传结构更加复杂 [69 -70 ] Eothenomys 东亚 高原种类与低海拔种类分裂时间约2.7 Ma [71 ] Schizothoracinae 青藏高原及周边地区 在晚中新世晚期或上新世裂腹鱼亚科鱼类就已经出现在青藏高原及周边水系中,在晚更新世期间种内分化加速,不同海拔之间的类群产生等级分化,种间多样性增加,第四纪以来青藏高原隆升导致的水系改组是推动裂腹鱼亚科鱼类演化的原动力 [31 ,76 -78 ]
3 讨论 最近,青藏高原古地貌重建工作取得了丰硕的成果(图3 ),但是各种指标重建结果往往存在矛盾(图4 )[25 ,79 -83 ] .同位素重建结果表明高原南部在古近纪早期就存在安第斯型的巨大山系[84 -85 ] ;古生物研究发现高原腹地从中始新世到晚渐新世都存在东西向的大峡谷,该峡谷底部海拔在2 000 m左右,两侧为高大的冈底斯山脉和羌塘山脉[82 ,86 -88 ] ,动植物如巨犀(Paraceratherium )[89 ] 、攀鲈(Eoanabas )[90 -91 ] 、臭椿(Ailanthus )[88 ] 、椿榆(Cedrelospermum )[92 ] 、兔耳果(Lagokarpos )[93 ] 等在古近纪都可以通过青藏地区峡谷与周边动植物区系交流,新近纪之前的青藏地区虽然地形复杂,但是尚未隆升成今天的高原;横断山区某些区域则在晚始新世-早渐新世之交就已经获得现今的海拔高度[94 -95 ] ;高原北部和喜马拉雅山脉则在新近纪形成现在的地貌格局[25 ,96 -97 ] .古生物研究也发现青藏高原在晚上新世就已经存在高寒生境和冰缘环境,成为第四纪冰期动物忍耐寒冷环境的“训练营”,第四纪冰期来临之后这些适应寒冷环境的生物才扩散至北半球高纬度地区[34 ,98 -99 ] .因此,高原及周边地区高寒生物类群的起源时间表明在渐新世青藏高原部分区域就已经出现高寒生态系统,晚中新世以来青藏高原地貌与气候继续发生剧烈变化,促使这些物种的祖先类群迁入或就地演化积累适应高寒环境的基因和形态,并最终演化成适应高寒生境的新物种[25 ] .高寒区物种在晚上新世的加速分化可能与高原巨大山系和水系、冰川活动等造成的地理隔离有关[26 ] .随着第四纪冰期的到来,青藏高原及周边地区的高寒生物区也开始与北半球高纬度地区存在密切联系[34 ,98 -99 ] .同时,演化生物学研究也发现在第四纪冰期期间并未形成覆盖整个青藏高原的大冰盖,高原面上仍存在很多生物避难所[26 ] . ...
Whole-genome sequencing of the snub-nosed monkey provides insights into folivory and evolutionary history
2
2014
... 哺乳动物中的藏羚羊(Pantholops hodgsonii )生活于西藏、青海和新疆等地海拔3 700~5 500 m的区域,能够成功适应高原缺氧环境,其系统发育学研究表明在上新世-早更新世期间该类群就已经生活在青藏高原地区[65 ] .藏原羚(Procapra picticaudata )现今生活于海拔3 000~5 100 m地区,其种内分化时间约为4.4~2.2 Ma[66 ] .灵长目仰鼻猴属(Rhinopithecus )鼻孔结构是对高原缺氧环境的适应,系统发育研究表明在1.6 Ma左右其分布于四川和云南的金丝猴北方类群与分布于喜马拉雅的金丝猴类群发生分化[67 ] .兔形目鼠兔属(Ochotona )广泛分布于北半球草原和灌丛,其分布于青藏高原的类群演化出适应高寒生境的习性,在2.4 Ma左右,适应高寒生境类群与低海拔泛北极分布类群分化[68 ] .中华姬鼠(Apodemus draco )分布于青藏高原及周边地区以及湖南、湖北和台湾等地,其在3.8~0.87 Ma期间分化形成适应高海拔生境的物种,其种系及遗传结构由于高原阶段性隆升和冰川活动的影响而变得更加复杂[69 -70 ] .绒鼠属(Eothenomys )高原类群与低海拔类群分化时间约为2.7 Ma[71 ] .基于4 966种现生陆栖脊椎动物和1 278个哺乳动物化石属的研究也表明受印度-欧亚板块碰撞、高原阶段性隆升、亚洲内陆干旱化等事件的影响,青藏高原及其邻近地区动物区系在新生代发生显著变化,其区系成分在早期更接近东洋界,并最终在中新世-上新世之交形成与现在接近的动物区系格局[72 ] .对174种青藏高原特有陆栖脊椎动物祖先分布区、来源和分化时间的估算,发现青藏高原特有种早在55 Ma就开始从周边区域扩散至高原,扩散事件约从15 Ma开始增加,并在6 Ma之后达到顶峰,高原隆升和历史气候变化对该地区动物区系的形成与演化具有重要作用[73 ] . ...
... Biological evolution in the alpine region of Tibetan Plateau
Table 1 种属 分布区域 适应高海拔类群分化时间或多样性增加时间 文献来源 Sinopodophyllum hexandrum 青藏高原东部 在约(6.52±1.89) Ma期间出现在青藏高原东部,但是直到3~2 Ma以来才演化出适应高山环境的形态 [47 ] Nannoglottis 青藏高原 在23~32 Ma期间出现在青藏高原,随后多样性增高,其高山灌丛和针叶林生活类型分支时间约为3.4 Ma [48 ] Reaumuria soongarica 西北干旱区、青藏高原北部 高原内部分布类群演化时间2.0~6.3 Ma,高原东北缘与高原西北类群分化时间1.1~3.5 Ma [49 ] Orchidinae 青藏高原及喜马拉雅山脉 青藏高原特有种大多是在过去6 Ma里进化而来 [50 ] Oxyria sinensis 青藏高原东南部 高山类群出现时间7.54~24.36 Ma,种内多样化增加时间1.74~0.86 Ma [51 ] Juniperus microsperma 西藏波密 早中新世21.6~14 Ma期间与其他种群分化 [52 ] Rhododendron 北半球 横断山区概属多样性在晚中新世(约8 Ma)快速增加 [53 ] Roscoea, Cautleya 喜马拉雅-横断山区 在早渐新世(~32 Ma)两属出现,Roscoea在横断山区与喜马拉雅山脉的两个分支大概在渐新世-中新世之交分化 [54 ] Nanorana parkeri 青藏高原南部 高原东部和西部类群分化时间1.4~3.7 Ma [55 ] Paini 南亚、东南亚、青藏高原 在渐新世早期棘蛙族高原-喜马拉雅类群与中国第二阶梯上生活的类群分化,随后在在26.1~12.5 Ma高原生活类群与喜马拉雅山脉生活类群分化,高原内部生活类群在晚中新世到上新世期间分化速率加快 [22 ] Bufo tibetanus 青藏高原、四川、甘肃、陕西等 高原种类形成于上新世期间 [56 ] Phrynocephalus 亚洲干旱区 在7.2~13.0 Ma期间演化出胎生类群,3.2~7.3 Ma期间种间多样性增加 [57 ] Phrynocephalus erythrurus 羌塘盆地 在3.67 Ma左右北部类群与南部类群发生分化,约2.76 Ma北部类群内部进一步分化 [58 ] Gnaptorina 青藏高原及周边高海拔地区 始新世起源于青藏高原东南缘 [18 -19 ] Coelotinae 北半球温带、亚热带 晚始新世高原东南缘隆升阻碍南方隙蛛向北扩散,渐新世高原中部和北部隆起导致北方隙蛛早期多样化进程中的灭绝事件 [59 ] Montifringilla, Onychostruthus, Pyrgilauda 青藏高原 雪雀的共同祖先在约14 Ma就应经生活在青藏高原,并进化出适应高海拔极端环境的性状 [21 ] Parus major 亚洲、欧洲 蒙古分布类群与其他种类分化时间为2.8~0.7 Ma,喜马拉雅山脉东部与中国中东部类群分化时间为1.9~0.4 Ma [60 ] Pseudopodoces humilis 青藏高原、柴达木盆地周边高山 在9.9~7.7 Ma期间地山雀就与与其他山雀分化开来,并出现适应高海拔性状 [61 ] Phylloscopus, Seicercus 亚洲 喜马拉雅山脉分布类群在7 Ma加速进化 [62 -63 ] Perdix 西藏、甘肃、青海、四川、新疆 高原类群出现于3.63 (4.52~2.75) Ma [64 ] Pantholops hodgsonii 西藏、新疆、青海 上新世-更新世就已经出现在青藏高原 [65 ] Procapra picticaudata 西藏、四川、甘肃、青海、新疆 种内分化时间约2.2~4.4 Ma [66 ] Rhinopithecus 四川、甘肃、陕西、贵州、云南、缅甸、越南 北方类群川金丝猴、黔金丝猴与喜马拉雅类群滇金丝猴、缅甸金丝猴分化时间约1.6 Ma [67 ] Ochotona 北半球 分布于青藏高原的高山类群和灌丛草原类群与低海拔泛北极分布类群分化时间大约为2.4 Ma [68 ] Apodemus draco 四川、云南、甘肃、青海、陕西、湖北、湖南、福建、台湾 2.2 Ma (3.8~0.87 Ma)分化形成,后来由于高原的阶段性隆升以及冰川作用种系及遗传结构更加复杂 [69 -70 ] Eothenomys 东亚 高原种类与低海拔种类分裂时间约2.7 Ma [71 ] Schizothoracinae 青藏高原及周边地区 在晚中新世晚期或上新世裂腹鱼亚科鱼类就已经出现在青藏高原及周边水系中,在晚更新世期间种内分化加速,不同海拔之间的类群产生等级分化,种间多样性增加,第四纪以来青藏高原隆升导致的水系改组是推动裂腹鱼亚科鱼类演化的原动力 [31 ,76 -78 ]
3 讨论 最近,青藏高原古地貌重建工作取得了丰硕的成果(图3 ),但是各种指标重建结果往往存在矛盾(图4 )[25 ,79 -83 ] .同位素重建结果表明高原南部在古近纪早期就存在安第斯型的巨大山系[84 -85 ] ;古生物研究发现高原腹地从中始新世到晚渐新世都存在东西向的大峡谷,该峡谷底部海拔在2 000 m左右,两侧为高大的冈底斯山脉和羌塘山脉[82 ,86 -88 ] ,动植物如巨犀(Paraceratherium )[89 ] 、攀鲈(Eoanabas )[90 -91 ] 、臭椿(Ailanthus )[88 ] 、椿榆(Cedrelospermum )[92 ] 、兔耳果(Lagokarpos )[93 ] 等在古近纪都可以通过青藏地区峡谷与周边动植物区系交流,新近纪之前的青藏地区虽然地形复杂,但是尚未隆升成今天的高原;横断山区某些区域则在晚始新世-早渐新世之交就已经获得现今的海拔高度[94 -95 ] ;高原北部和喜马拉雅山脉则在新近纪形成现在的地貌格局[25 ,96 -97 ] .古生物研究也发现青藏高原在晚上新世就已经存在高寒生境和冰缘环境,成为第四纪冰期动物忍耐寒冷环境的“训练营”,第四纪冰期来临之后这些适应寒冷环境的生物才扩散至北半球高纬度地区[34 ,98 -99 ] .因此,高原及周边地区高寒生物类群的起源时间表明在渐新世青藏高原部分区域就已经出现高寒生态系统,晚中新世以来青藏高原地貌与气候继续发生剧烈变化,促使这些物种的祖先类群迁入或就地演化积累适应高寒环境的基因和形态,并最终演化成适应高寒生境的新物种[25 ] .高寒区物种在晚上新世的加速分化可能与高原巨大山系和水系、冰川活动等造成的地理隔离有关[26 ] .随着第四纪冰期的到来,青藏高原及周边地区的高寒生物区也开始与北半球高纬度地区存在密切联系[34 ,98 -99 ] .同时,演化生物学研究也发现在第四纪冰期期间并未形成覆盖整个青藏高原的大冰盖,高原面上仍存在很多生物避难所[26 ] . ...
Molecular systematics of pikas (genus Ochotona ) inferred from mitochondrial DNA sequences
2
2000
... 哺乳动物中的藏羚羊(Pantholops hodgsonii )生活于西藏、青海和新疆等地海拔3 700~5 500 m的区域,能够成功适应高原缺氧环境,其系统发育学研究表明在上新世-早更新世期间该类群就已经生活在青藏高原地区[65 ] .藏原羚(Procapra picticaudata )现今生活于海拔3 000~5 100 m地区,其种内分化时间约为4.4~2.2 Ma[66 ] .灵长目仰鼻猴属(Rhinopithecus )鼻孔结构是对高原缺氧环境的适应,系统发育研究表明在1.6 Ma左右其分布于四川和云南的金丝猴北方类群与分布于喜马拉雅的金丝猴类群发生分化[67 ] .兔形目鼠兔属(Ochotona )广泛分布于北半球草原和灌丛,其分布于青藏高原的类群演化出适应高寒生境的习性,在2.4 Ma左右,适应高寒生境类群与低海拔泛北极分布类群分化[68 ] .中华姬鼠(Apodemus draco )分布于青藏高原及周边地区以及湖南、湖北和台湾等地,其在3.8~0.87 Ma期间分化形成适应高海拔生境的物种,其种系及遗传结构由于高原阶段性隆升和冰川活动的影响而变得更加复杂[69 -70 ] .绒鼠属(Eothenomys )高原类群与低海拔类群分化时间约为2.7 Ma[71 ] .基于4 966种现生陆栖脊椎动物和1 278个哺乳动物化石属的研究也表明受印度-欧亚板块碰撞、高原阶段性隆升、亚洲内陆干旱化等事件的影响,青藏高原及其邻近地区动物区系在新生代发生显著变化,其区系成分在早期更接近东洋界,并最终在中新世-上新世之交形成与现在接近的动物区系格局[72 ] .对174种青藏高原特有陆栖脊椎动物祖先分布区、来源和分化时间的估算,发现青藏高原特有种早在55 Ma就开始从周边区域扩散至高原,扩散事件约从15 Ma开始增加,并在6 Ma之后达到顶峰,高原隆升和历史气候变化对该地区动物区系的形成与演化具有重要作用[73 ] . ...
... Biological evolution in the alpine region of Tibetan Plateau
Table 1 种属 分布区域 适应高海拔类群分化时间或多样性增加时间 文献来源 Sinopodophyllum hexandrum 青藏高原东部 在约(6.52±1.89) Ma期间出现在青藏高原东部,但是直到3~2 Ma以来才演化出适应高山环境的形态 [47 ] Nannoglottis 青藏高原 在23~32 Ma期间出现在青藏高原,随后多样性增高,其高山灌丛和针叶林生活类型分支时间约为3.4 Ma [48 ] Reaumuria soongarica 西北干旱区、青藏高原北部 高原内部分布类群演化时间2.0~6.3 Ma,高原东北缘与高原西北类群分化时间1.1~3.5 Ma [49 ] Orchidinae 青藏高原及喜马拉雅山脉 青藏高原特有种大多是在过去6 Ma里进化而来 [50 ] Oxyria sinensis 青藏高原东南部 高山类群出现时间7.54~24.36 Ma,种内多样化增加时间1.74~0.86 Ma [51 ] Juniperus microsperma 西藏波密 早中新世21.6~14 Ma期间与其他种群分化 [52 ] Rhododendron 北半球 横断山区概属多样性在晚中新世(约8 Ma)快速增加 [53 ] Roscoea, Cautleya 喜马拉雅-横断山区 在早渐新世(~32 Ma)两属出现,Roscoea在横断山区与喜马拉雅山脉的两个分支大概在渐新世-中新世之交分化 [54 ] Nanorana parkeri 青藏高原南部 高原东部和西部类群分化时间1.4~3.7 Ma [55 ] Paini 南亚、东南亚、青藏高原 在渐新世早期棘蛙族高原-喜马拉雅类群与中国第二阶梯上生活的类群分化,随后在在26.1~12.5 Ma高原生活类群与喜马拉雅山脉生活类群分化,高原内部生活类群在晚中新世到上新世期间分化速率加快 [22 ] Bufo tibetanus 青藏高原、四川、甘肃、陕西等 高原种类形成于上新世期间 [56 ] Phrynocephalus 亚洲干旱区 在7.2~13.0 Ma期间演化出胎生类群,3.2~7.3 Ma期间种间多样性增加 [57 ] Phrynocephalus erythrurus 羌塘盆地 在3.67 Ma左右北部类群与南部类群发生分化,约2.76 Ma北部类群内部进一步分化 [58 ] Gnaptorina 青藏高原及周边高海拔地区 始新世起源于青藏高原东南缘 [18 -19 ] Coelotinae 北半球温带、亚热带 晚始新世高原东南缘隆升阻碍南方隙蛛向北扩散,渐新世高原中部和北部隆起导致北方隙蛛早期多样化进程中的灭绝事件 [59 ] Montifringilla, Onychostruthus, Pyrgilauda 青藏高原 雪雀的共同祖先在约14 Ma就应经生活在青藏高原,并进化出适应高海拔极端环境的性状 [21 ] Parus major 亚洲、欧洲 蒙古分布类群与其他种类分化时间为2.8~0.7 Ma,喜马拉雅山脉东部与中国中东部类群分化时间为1.9~0.4 Ma [60 ] Pseudopodoces humilis 青藏高原、柴达木盆地周边高山 在9.9~7.7 Ma期间地山雀就与与其他山雀分化开来,并出现适应高海拔性状 [61 ] Phylloscopus, Seicercus 亚洲 喜马拉雅山脉分布类群在7 Ma加速进化 [62 -63 ] Perdix 西藏、甘肃、青海、四川、新疆 高原类群出现于3.63 (4.52~2.75) Ma [64 ] Pantholops hodgsonii 西藏、新疆、青海 上新世-更新世就已经出现在青藏高原 [65 ] Procapra picticaudata 西藏、四川、甘肃、青海、新疆 种内分化时间约2.2~4.4 Ma [66 ] Rhinopithecus 四川、甘肃、陕西、贵州、云南、缅甸、越南 北方类群川金丝猴、黔金丝猴与喜马拉雅类群滇金丝猴、缅甸金丝猴分化时间约1.6 Ma [67 ] Ochotona 北半球 分布于青藏高原的高山类群和灌丛草原类群与低海拔泛北极分布类群分化时间大约为2.4 Ma [68 ] Apodemus draco 四川、云南、甘肃、青海、陕西、湖北、湖南、福建、台湾 2.2 Ma (3.8~0.87 Ma)分化形成,后来由于高原的阶段性隆升以及冰川作用种系及遗传结构更加复杂 [69 -70 ] Eothenomys 东亚 高原种类与低海拔种类分裂时间约2.7 Ma [71 ] Schizothoracinae 青藏高原及周边地区 在晚中新世晚期或上新世裂腹鱼亚科鱼类就已经出现在青藏高原及周边水系中,在晚更新世期间种内分化加速,不同海拔之间的类群产生等级分化,种间多样性增加,第四纪以来青藏高原隆升导致的水系改组是推动裂腹鱼亚科鱼类演化的原动力 [31 ,76 -78 ]
3 讨论 最近,青藏高原古地貌重建工作取得了丰硕的成果(图3 ),但是各种指标重建结果往往存在矛盾(图4 )[25 ,79 -83 ] .同位素重建结果表明高原南部在古近纪早期就存在安第斯型的巨大山系[84 -85 ] ;古生物研究发现高原腹地从中始新世到晚渐新世都存在东西向的大峡谷,该峡谷底部海拔在2 000 m左右,两侧为高大的冈底斯山脉和羌塘山脉[82 ,86 -88 ] ,动植物如巨犀(Paraceratherium )[89 ] 、攀鲈(Eoanabas )[90 -91 ] 、臭椿(Ailanthus )[88 ] 、椿榆(Cedrelospermum )[92 ] 、兔耳果(Lagokarpos )[93 ] 等在古近纪都可以通过青藏地区峡谷与周边动植物区系交流,新近纪之前的青藏地区虽然地形复杂,但是尚未隆升成今天的高原;横断山区某些区域则在晚始新世-早渐新世之交就已经获得现今的海拔高度[94 -95 ] ;高原北部和喜马拉雅山脉则在新近纪形成现在的地貌格局[25 ,96 -97 ] .古生物研究也发现青藏高原在晚上新世就已经存在高寒生境和冰缘环境,成为第四纪冰期动物忍耐寒冷环境的“训练营”,第四纪冰期来临之后这些适应寒冷环境的生物才扩散至北半球高纬度地区[34 ,98 -99 ] .因此,高原及周边地区高寒生物类群的起源时间表明在渐新世青藏高原部分区域就已经出现高寒生态系统,晚中新世以来青藏高原地貌与气候继续发生剧烈变化,促使这些物种的祖先类群迁入或就地演化积累适应高寒环境的基因和形态,并最终演化成适应高寒生境的新物种[25 ] .高寒区物种在晚上新世的加速分化可能与高原巨大山系和水系、冰川活动等造成的地理隔离有关[26 ] .随着第四纪冰期的到来,青藏高原及周边地区的高寒生物区也开始与北半球高纬度地区存在密切联系[34 ,98 -99 ] .同时,演化生物学研究也发现在第四纪冰期期间并未形成覆盖整个青藏高原的大冰盖,高原面上仍存在很多生物避难所[26 ] . ...
Comparative phylogeography of four Apodemus species (Mammalia: Rodentia) in the Asian Far East: evidence of Quaternary climatic changes in their genetic structure
2
2010
... 哺乳动物中的藏羚羊(Pantholops hodgsonii )生活于西藏、青海和新疆等地海拔3 700~5 500 m的区域,能够成功适应高原缺氧环境,其系统发育学研究表明在上新世-早更新世期间该类群就已经生活在青藏高原地区[65 ] .藏原羚(Procapra picticaudata )现今生活于海拔3 000~5 100 m地区,其种内分化时间约为4.4~2.2 Ma[66 ] .灵长目仰鼻猴属(Rhinopithecus )鼻孔结构是对高原缺氧环境的适应,系统发育研究表明在1.6 Ma左右其分布于四川和云南的金丝猴北方类群与分布于喜马拉雅的金丝猴类群发生分化[67 ] .兔形目鼠兔属(Ochotona )广泛分布于北半球草原和灌丛,其分布于青藏高原的类群演化出适应高寒生境的习性,在2.4 Ma左右,适应高寒生境类群与低海拔泛北极分布类群分化[68 ] .中华姬鼠(Apodemus draco )分布于青藏高原及周边地区以及湖南、湖北和台湾等地,其在3.8~0.87 Ma期间分化形成适应高海拔生境的物种,其种系及遗传结构由于高原阶段性隆升和冰川活动的影响而变得更加复杂[69 -70 ] .绒鼠属(Eothenomys )高原类群与低海拔类群分化时间约为2.7 Ma[71 ] .基于4 966种现生陆栖脊椎动物和1 278个哺乳动物化石属的研究也表明受印度-欧亚板块碰撞、高原阶段性隆升、亚洲内陆干旱化等事件的影响,青藏高原及其邻近地区动物区系在新生代发生显著变化,其区系成分在早期更接近东洋界,并最终在中新世-上新世之交形成与现在接近的动物区系格局[72 ] .对174种青藏高原特有陆栖脊椎动物祖先分布区、来源和分化时间的估算,发现青藏高原特有种早在55 Ma就开始从周边区域扩散至高原,扩散事件约从15 Ma开始增加,并在6 Ma之后达到顶峰,高原隆升和历史气候变化对该地区动物区系的形成与演化具有重要作用[73 ] . ...
... Biological evolution in the alpine region of Tibetan Plateau
Table 1 种属 分布区域 适应高海拔类群分化时间或多样性增加时间 文献来源 Sinopodophyllum hexandrum 青藏高原东部 在约(6.52±1.89) Ma期间出现在青藏高原东部,但是直到3~2 Ma以来才演化出适应高山环境的形态 [47 ] Nannoglottis 青藏高原 在23~32 Ma期间出现在青藏高原,随后多样性增高,其高山灌丛和针叶林生活类型分支时间约为3.4 Ma [48 ] Reaumuria soongarica 西北干旱区、青藏高原北部 高原内部分布类群演化时间2.0~6.3 Ma,高原东北缘与高原西北类群分化时间1.1~3.5 Ma [49 ] Orchidinae 青藏高原及喜马拉雅山脉 青藏高原特有种大多是在过去6 Ma里进化而来 [50 ] Oxyria sinensis 青藏高原东南部 高山类群出现时间7.54~24.36 Ma,种内多样化增加时间1.74~0.86 Ma [51 ] Juniperus microsperma 西藏波密 早中新世21.6~14 Ma期间与其他种群分化 [52 ] Rhododendron 北半球 横断山区概属多样性在晚中新世(约8 Ma)快速增加 [53 ] Roscoea, Cautleya 喜马拉雅-横断山区 在早渐新世(~32 Ma)两属出现,Roscoea在横断山区与喜马拉雅山脉的两个分支大概在渐新世-中新世之交分化 [54 ] Nanorana parkeri 青藏高原南部 高原东部和西部类群分化时间1.4~3.7 Ma [55 ] Paini 南亚、东南亚、青藏高原 在渐新世早期棘蛙族高原-喜马拉雅类群与中国第二阶梯上生活的类群分化,随后在在26.1~12.5 Ma高原生活类群与喜马拉雅山脉生活类群分化,高原内部生活类群在晚中新世到上新世期间分化速率加快 [22 ] Bufo tibetanus 青藏高原、四川、甘肃、陕西等 高原种类形成于上新世期间 [56 ] Phrynocephalus 亚洲干旱区 在7.2~13.0 Ma期间演化出胎生类群,3.2~7.3 Ma期间种间多样性增加 [57 ] Phrynocephalus erythrurus 羌塘盆地 在3.67 Ma左右北部类群与南部类群发生分化,约2.76 Ma北部类群内部进一步分化 [58 ] Gnaptorina 青藏高原及周边高海拔地区 始新世起源于青藏高原东南缘 [18 -19 ] Coelotinae 北半球温带、亚热带 晚始新世高原东南缘隆升阻碍南方隙蛛向北扩散,渐新世高原中部和北部隆起导致北方隙蛛早期多样化进程中的灭绝事件 [59 ] Montifringilla, Onychostruthus, Pyrgilauda 青藏高原 雪雀的共同祖先在约14 Ma就应经生活在青藏高原,并进化出适应高海拔极端环境的性状 [21 ] Parus major 亚洲、欧洲 蒙古分布类群与其他种类分化时间为2.8~0.7 Ma,喜马拉雅山脉东部与中国中东部类群分化时间为1.9~0.4 Ma [60 ] Pseudopodoces humilis 青藏高原、柴达木盆地周边高山 在9.9~7.7 Ma期间地山雀就与与其他山雀分化开来,并出现适应高海拔性状 [61 ] Phylloscopus, Seicercus 亚洲 喜马拉雅山脉分布类群在7 Ma加速进化 [62 -63 ] Perdix 西藏、甘肃、青海、四川、新疆 高原类群出现于3.63 (4.52~2.75) Ma [64 ] Pantholops hodgsonii 西藏、新疆、青海 上新世-更新世就已经出现在青藏高原 [65 ] Procapra picticaudata 西藏、四川、甘肃、青海、新疆 种内分化时间约2.2~4.4 Ma [66 ] Rhinopithecus 四川、甘肃、陕西、贵州、云南、缅甸、越南 北方类群川金丝猴、黔金丝猴与喜马拉雅类群滇金丝猴、缅甸金丝猴分化时间约1.6 Ma [67 ] Ochotona 北半球 分布于青藏高原的高山类群和灌丛草原类群与低海拔泛北极分布类群分化时间大约为2.4 Ma [68 ] Apodemus draco 四川、云南、甘肃、青海、陕西、湖北、湖南、福建、台湾 2.2 Ma (3.8~0.87 Ma)分化形成,后来由于高原的阶段性隆升以及冰川作用种系及遗传结构更加复杂 [69 -70 ] Eothenomys 东亚 高原种类与低海拔种类分裂时间约2.7 Ma [71 ] Schizothoracinae 青藏高原及周边地区 在晚中新世晚期或上新世裂腹鱼亚科鱼类就已经出现在青藏高原及周边水系中,在晚更新世期间种内分化加速,不同海拔之间的类群产生等级分化,种间多样性增加,第四纪以来青藏高原隆升导致的水系改组是推动裂腹鱼亚科鱼类演化的原动力 [31 ,76 -78 ]
3 讨论 最近,青藏高原古地貌重建工作取得了丰硕的成果(图3 ),但是各种指标重建结果往往存在矛盾(图4 )[25 ,79 -83 ] .同位素重建结果表明高原南部在古近纪早期就存在安第斯型的巨大山系[84 -85 ] ;古生物研究发现高原腹地从中始新世到晚渐新世都存在东西向的大峡谷,该峡谷底部海拔在2 000 m左右,两侧为高大的冈底斯山脉和羌塘山脉[82 ,86 -88 ] ,动植物如巨犀(Paraceratherium )[89 ] 、攀鲈(Eoanabas )[90 -91 ] 、臭椿(Ailanthus )[88 ] 、椿榆(Cedrelospermum )[92 ] 、兔耳果(Lagokarpos )[93 ] 等在古近纪都可以通过青藏地区峡谷与周边动植物区系交流,新近纪之前的青藏地区虽然地形复杂,但是尚未隆升成今天的高原;横断山区某些区域则在晚始新世-早渐新世之交就已经获得现今的海拔高度[94 -95 ] ;高原北部和喜马拉雅山脉则在新近纪形成现在的地貌格局[25 ,96 -97 ] .古生物研究也发现青藏高原在晚上新世就已经存在高寒生境和冰缘环境,成为第四纪冰期动物忍耐寒冷环境的“训练营”,第四纪冰期来临之后这些适应寒冷环境的生物才扩散至北半球高纬度地区[34 ,98 -99 ] .因此,高原及周边地区高寒生物类群的起源时间表明在渐新世青藏高原部分区域就已经出现高寒生态系统,晚中新世以来青藏高原地貌与气候继续发生剧烈变化,促使这些物种的祖先类群迁入或就地演化积累适应高寒环境的基因和形态,并最终演化成适应高寒生境的新物种[25 ] .高寒区物种在晚上新世的加速分化可能与高原巨大山系和水系、冰川活动等造成的地理隔离有关[26 ] .随着第四纪冰期的到来,青藏高原及周边地区的高寒生物区也开始与北半球高纬度地区存在密切联系[34 ,98 -99 ] .同时,演化生物学研究也发现在第四纪冰期期间并未形成覆盖整个青藏高原的大冰盖,高原面上仍存在很多生物避难所[26 ] . ...
Phylogeography of the South China field mouse (Apodemus draco ) on the Southeastern Tibetan Plateau reveals high genetic diversity and glacial refugia
2
2012
... 哺乳动物中的藏羚羊(Pantholops hodgsonii )生活于西藏、青海和新疆等地海拔3 700~5 500 m的区域,能够成功适应高原缺氧环境,其系统发育学研究表明在上新世-早更新世期间该类群就已经生活在青藏高原地区[65 ] .藏原羚(Procapra picticaudata )现今生活于海拔3 000~5 100 m地区,其种内分化时间约为4.4~2.2 Ma[66 ] .灵长目仰鼻猴属(Rhinopithecus )鼻孔结构是对高原缺氧环境的适应,系统发育研究表明在1.6 Ma左右其分布于四川和云南的金丝猴北方类群与分布于喜马拉雅的金丝猴类群发生分化[67 ] .兔形目鼠兔属(Ochotona )广泛分布于北半球草原和灌丛,其分布于青藏高原的类群演化出适应高寒生境的习性,在2.4 Ma左右,适应高寒生境类群与低海拔泛北极分布类群分化[68 ] .中华姬鼠(Apodemus draco )分布于青藏高原及周边地区以及湖南、湖北和台湾等地,其在3.8~0.87 Ma期间分化形成适应高海拔生境的物种,其种系及遗传结构由于高原阶段性隆升和冰川活动的影响而变得更加复杂[69 -70 ] .绒鼠属(Eothenomys )高原类群与低海拔类群分化时间约为2.7 Ma[71 ] .基于4 966种现生陆栖脊椎动物和1 278个哺乳动物化石属的研究也表明受印度-欧亚板块碰撞、高原阶段性隆升、亚洲内陆干旱化等事件的影响,青藏高原及其邻近地区动物区系在新生代发生显著变化,其区系成分在早期更接近东洋界,并最终在中新世-上新世之交形成与现在接近的动物区系格局[72 ] .对174种青藏高原特有陆栖脊椎动物祖先分布区、来源和分化时间的估算,发现青藏高原特有种早在55 Ma就开始从周边区域扩散至高原,扩散事件约从15 Ma开始增加,并在6 Ma之后达到顶峰,高原隆升和历史气候变化对该地区动物区系的形成与演化具有重要作用[73 ] . ...
... Biological evolution in the alpine region of Tibetan Plateau
Table 1 种属 分布区域 适应高海拔类群分化时间或多样性增加时间 文献来源 Sinopodophyllum hexandrum 青藏高原东部 在约(6.52±1.89) Ma期间出现在青藏高原东部,但是直到3~2 Ma以来才演化出适应高山环境的形态 [47 ] Nannoglottis 青藏高原 在23~32 Ma期间出现在青藏高原,随后多样性增高,其高山灌丛和针叶林生活类型分支时间约为3.4 Ma [48 ] Reaumuria soongarica 西北干旱区、青藏高原北部 高原内部分布类群演化时间2.0~6.3 Ma,高原东北缘与高原西北类群分化时间1.1~3.5 Ma [49 ] Orchidinae 青藏高原及喜马拉雅山脉 青藏高原特有种大多是在过去6 Ma里进化而来 [50 ] Oxyria sinensis 青藏高原东南部 高山类群出现时间7.54~24.36 Ma,种内多样化增加时间1.74~0.86 Ma [51 ] Juniperus microsperma 西藏波密 早中新世21.6~14 Ma期间与其他种群分化 [52 ] Rhododendron 北半球 横断山区概属多样性在晚中新世(约8 Ma)快速增加 [53 ] Roscoea, Cautleya 喜马拉雅-横断山区 在早渐新世(~32 Ma)两属出现,Roscoea在横断山区与喜马拉雅山脉的两个分支大概在渐新世-中新世之交分化 [54 ] Nanorana parkeri 青藏高原南部 高原东部和西部类群分化时间1.4~3.7 Ma [55 ] Paini 南亚、东南亚、青藏高原 在渐新世早期棘蛙族高原-喜马拉雅类群与中国第二阶梯上生活的类群分化,随后在在26.1~12.5 Ma高原生活类群与喜马拉雅山脉生活类群分化,高原内部生活类群在晚中新世到上新世期间分化速率加快 [22 ] Bufo tibetanus 青藏高原、四川、甘肃、陕西等 高原种类形成于上新世期间 [56 ] Phrynocephalus 亚洲干旱区 在7.2~13.0 Ma期间演化出胎生类群,3.2~7.3 Ma期间种间多样性增加 [57 ] Phrynocephalus erythrurus 羌塘盆地 在3.67 Ma左右北部类群与南部类群发生分化,约2.76 Ma北部类群内部进一步分化 [58 ] Gnaptorina 青藏高原及周边高海拔地区 始新世起源于青藏高原东南缘 [18 -19 ] Coelotinae 北半球温带、亚热带 晚始新世高原东南缘隆升阻碍南方隙蛛向北扩散,渐新世高原中部和北部隆起导致北方隙蛛早期多样化进程中的灭绝事件 [59 ] Montifringilla, Onychostruthus, Pyrgilauda 青藏高原 雪雀的共同祖先在约14 Ma就应经生活在青藏高原,并进化出适应高海拔极端环境的性状 [21 ] Parus major 亚洲、欧洲 蒙古分布类群与其他种类分化时间为2.8~0.7 Ma,喜马拉雅山脉东部与中国中东部类群分化时间为1.9~0.4 Ma [60 ] Pseudopodoces humilis 青藏高原、柴达木盆地周边高山 在9.9~7.7 Ma期间地山雀就与与其他山雀分化开来,并出现适应高海拔性状 [61 ] Phylloscopus, Seicercus 亚洲 喜马拉雅山脉分布类群在7 Ma加速进化 [62 -63 ] Perdix 西藏、甘肃、青海、四川、新疆 高原类群出现于3.63 (4.52~2.75) Ma [64 ] Pantholops hodgsonii 西藏、新疆、青海 上新世-更新世就已经出现在青藏高原 [65 ] Procapra picticaudata 西藏、四川、甘肃、青海、新疆 种内分化时间约2.2~4.4 Ma [66 ] Rhinopithecus 四川、甘肃、陕西、贵州、云南、缅甸、越南 北方类群川金丝猴、黔金丝猴与喜马拉雅类群滇金丝猴、缅甸金丝猴分化时间约1.6 Ma [67 ] Ochotona 北半球 分布于青藏高原的高山类群和灌丛草原类群与低海拔泛北极分布类群分化时间大约为2.4 Ma [68 ] Apodemus draco 四川、云南、甘肃、青海、陕西、湖北、湖南、福建、台湾 2.2 Ma (3.8~0.87 Ma)分化形成,后来由于高原的阶段性隆升以及冰川作用种系及遗传结构更加复杂 [69 -70 ] Eothenomys 东亚 高原种类与低海拔种类分裂时间约2.7 Ma [71 ] Schizothoracinae 青藏高原及周边地区 在晚中新世晚期或上新世裂腹鱼亚科鱼类就已经出现在青藏高原及周边水系中,在晚更新世期间种内分化加速,不同海拔之间的类群产生等级分化,种间多样性增加,第四纪以来青藏高原隆升导致的水系改组是推动裂腹鱼亚科鱼类演化的原动力 [31 ,76 -78 ]
3 讨论 最近,青藏高原古地貌重建工作取得了丰硕的成果(图3 ),但是各种指标重建结果往往存在矛盾(图4 )[25 ,79 -83 ] .同位素重建结果表明高原南部在古近纪早期就存在安第斯型的巨大山系[84 -85 ] ;古生物研究发现高原腹地从中始新世到晚渐新世都存在东西向的大峡谷,该峡谷底部海拔在2 000 m左右,两侧为高大的冈底斯山脉和羌塘山脉[82 ,86 -88 ] ,动植物如巨犀(Paraceratherium )[89 ] 、攀鲈(Eoanabas )[90 -91 ] 、臭椿(Ailanthus )[88 ] 、椿榆(Cedrelospermum )[92 ] 、兔耳果(Lagokarpos )[93 ] 等在古近纪都可以通过青藏地区峡谷与周边动植物区系交流,新近纪之前的青藏地区虽然地形复杂,但是尚未隆升成今天的高原;横断山区某些区域则在晚始新世-早渐新世之交就已经获得现今的海拔高度[94 -95 ] ;高原北部和喜马拉雅山脉则在新近纪形成现在的地貌格局[25 ,96 -97 ] .古生物研究也发现青藏高原在晚上新世就已经存在高寒生境和冰缘环境,成为第四纪冰期动物忍耐寒冷环境的“训练营”,第四纪冰期来临之后这些适应寒冷环境的生物才扩散至北半球高纬度地区[34 ,98 -99 ] .因此,高原及周边地区高寒生物类群的起源时间表明在渐新世青藏高原部分区域就已经出现高寒生态系统,晚中新世以来青藏高原地貌与气候继续发生剧烈变化,促使这些物种的祖先类群迁入或就地演化积累适应高寒环境的基因和形态,并最终演化成适应高寒生境的新物种[25 ] .高寒区物种在晚上新世的加速分化可能与高原巨大山系和水系、冰川活动等造成的地理隔离有关[26 ] .随着第四纪冰期的到来,青藏高原及周边地区的高寒生物区也开始与北半球高纬度地区存在密切联系[34 ,98 -99 ] .同时,演化生物学研究也发现在第四纪冰期期间并未形成覆盖整个青藏高原的大冰盖,高原面上仍存在很多生物避难所[26 ] . ...
Molecular phylogeny and biogeography of Oriental voles: genus Eothenomys (Muridae, Mammalia)
2
2004
... 哺乳动物中的藏羚羊(Pantholops hodgsonii )生活于西藏、青海和新疆等地海拔3 700~5 500 m的区域,能够成功适应高原缺氧环境,其系统发育学研究表明在上新世-早更新世期间该类群就已经生活在青藏高原地区[65 ] .藏原羚(Procapra picticaudata )现今生活于海拔3 000~5 100 m地区,其种内分化时间约为4.4~2.2 Ma[66 ] .灵长目仰鼻猴属(Rhinopithecus )鼻孔结构是对高原缺氧环境的适应,系统发育研究表明在1.6 Ma左右其分布于四川和云南的金丝猴北方类群与分布于喜马拉雅的金丝猴类群发生分化[67 ] .兔形目鼠兔属(Ochotona )广泛分布于北半球草原和灌丛,其分布于青藏高原的类群演化出适应高寒生境的习性,在2.4 Ma左右,适应高寒生境类群与低海拔泛北极分布类群分化[68 ] .中华姬鼠(Apodemus draco )分布于青藏高原及周边地区以及湖南、湖北和台湾等地,其在3.8~0.87 Ma期间分化形成适应高海拔生境的物种,其种系及遗传结构由于高原阶段性隆升和冰川活动的影响而变得更加复杂[69 -70 ] .绒鼠属(Eothenomys )高原类群与低海拔类群分化时间约为2.7 Ma[71 ] .基于4 966种现生陆栖脊椎动物和1 278个哺乳动物化石属的研究也表明受印度-欧亚板块碰撞、高原阶段性隆升、亚洲内陆干旱化等事件的影响,青藏高原及其邻近地区动物区系在新生代发生显著变化,其区系成分在早期更接近东洋界,并最终在中新世-上新世之交形成与现在接近的动物区系格局[72 ] .对174种青藏高原特有陆栖脊椎动物祖先分布区、来源和分化时间的估算,发现青藏高原特有种早在55 Ma就开始从周边区域扩散至高原,扩散事件约从15 Ma开始增加,并在6 Ma之后达到顶峰,高原隆升和历史气候变化对该地区动物区系的形成与演化具有重要作用[73 ] . ...
... Biological evolution in the alpine region of Tibetan Plateau
Table 1 种属 分布区域 适应高海拔类群分化时间或多样性增加时间 文献来源 Sinopodophyllum hexandrum 青藏高原东部 在约(6.52±1.89) Ma期间出现在青藏高原东部,但是直到3~2 Ma以来才演化出适应高山环境的形态 [47 ] Nannoglottis 青藏高原 在23~32 Ma期间出现在青藏高原,随后多样性增高,其高山灌丛和针叶林生活类型分支时间约为3.4 Ma [48 ] Reaumuria soongarica 西北干旱区、青藏高原北部 高原内部分布类群演化时间2.0~6.3 Ma,高原东北缘与高原西北类群分化时间1.1~3.5 Ma [49 ] Orchidinae 青藏高原及喜马拉雅山脉 青藏高原特有种大多是在过去6 Ma里进化而来 [50 ] Oxyria sinensis 青藏高原东南部 高山类群出现时间7.54~24.36 Ma,种内多样化增加时间1.74~0.86 Ma [51 ] Juniperus microsperma 西藏波密 早中新世21.6~14 Ma期间与其他种群分化 [52 ] Rhododendron 北半球 横断山区概属多样性在晚中新世(约8 Ma)快速增加 [53 ] Roscoea, Cautleya 喜马拉雅-横断山区 在早渐新世(~32 Ma)两属出现,Roscoea在横断山区与喜马拉雅山脉的两个分支大概在渐新世-中新世之交分化 [54 ] Nanorana parkeri 青藏高原南部 高原东部和西部类群分化时间1.4~3.7 Ma [55 ] Paini 南亚、东南亚、青藏高原 在渐新世早期棘蛙族高原-喜马拉雅类群与中国第二阶梯上生活的类群分化,随后在在26.1~12.5 Ma高原生活类群与喜马拉雅山脉生活类群分化,高原内部生活类群在晚中新世到上新世期间分化速率加快 [22 ] Bufo tibetanus 青藏高原、四川、甘肃、陕西等 高原种类形成于上新世期间 [56 ] Phrynocephalus 亚洲干旱区 在7.2~13.0 Ma期间演化出胎生类群,3.2~7.3 Ma期间种间多样性增加 [57 ] Phrynocephalus erythrurus 羌塘盆地 在3.67 Ma左右北部类群与南部类群发生分化,约2.76 Ma北部类群内部进一步分化 [58 ] Gnaptorina 青藏高原及周边高海拔地区 始新世起源于青藏高原东南缘 [18 -19 ] Coelotinae 北半球温带、亚热带 晚始新世高原东南缘隆升阻碍南方隙蛛向北扩散,渐新世高原中部和北部隆起导致北方隙蛛早期多样化进程中的灭绝事件 [59 ] Montifringilla, Onychostruthus, Pyrgilauda 青藏高原 雪雀的共同祖先在约14 Ma就应经生活在青藏高原,并进化出适应高海拔极端环境的性状 [21 ] Parus major 亚洲、欧洲 蒙古分布类群与其他种类分化时间为2.8~0.7 Ma,喜马拉雅山脉东部与中国中东部类群分化时间为1.9~0.4 Ma [60 ] Pseudopodoces humilis 青藏高原、柴达木盆地周边高山 在9.9~7.7 Ma期间地山雀就与与其他山雀分化开来,并出现适应高海拔性状 [61 ] Phylloscopus, Seicercus 亚洲 喜马拉雅山脉分布类群在7 Ma加速进化 [62 -63 ] Perdix 西藏、甘肃、青海、四川、新疆 高原类群出现于3.63 (4.52~2.75) Ma [64 ] Pantholops hodgsonii 西藏、新疆、青海 上新世-更新世就已经出现在青藏高原 [65 ] Procapra picticaudata 西藏、四川、甘肃、青海、新疆 种内分化时间约2.2~4.4 Ma [66 ] Rhinopithecus 四川、甘肃、陕西、贵州、云南、缅甸、越南 北方类群川金丝猴、黔金丝猴与喜马拉雅类群滇金丝猴、缅甸金丝猴分化时间约1.6 Ma [67 ] Ochotona 北半球 分布于青藏高原的高山类群和灌丛草原类群与低海拔泛北极分布类群分化时间大约为2.4 Ma [68 ] Apodemus draco 四川、云南、甘肃、青海、陕西、湖北、湖南、福建、台湾 2.2 Ma (3.8~0.87 Ma)分化形成,后来由于高原的阶段性隆升以及冰川作用种系及遗传结构更加复杂 [69 -70 ] Eothenomys 东亚 高原种类与低海拔种类分裂时间约2.7 Ma [71 ] Schizothoracinae 青藏高原及周边地区 在晚中新世晚期或上新世裂腹鱼亚科鱼类就已经出现在青藏高原及周边水系中,在晚更新世期间种内分化加速,不同海拔之间的类群产生等级分化,种间多样性增加,第四纪以来青藏高原隆升导致的水系改组是推动裂腹鱼亚科鱼类演化的原动力 [31 ,76 -78 ]
3 讨论 最近,青藏高原古地貌重建工作取得了丰硕的成果(图3 ),但是各种指标重建结果往往存在矛盾(图4 )[25 ,79 -83 ] .同位素重建结果表明高原南部在古近纪早期就存在安第斯型的巨大山系[84 -85 ] ;古生物研究发现高原腹地从中始新世到晚渐新世都存在东西向的大峡谷,该峡谷底部海拔在2 000 m左右,两侧为高大的冈底斯山脉和羌塘山脉[82 ,86 -88 ] ,动植物如巨犀(Paraceratherium )[89 ] 、攀鲈(Eoanabas )[90 -91 ] 、臭椿(Ailanthus )[88 ] 、椿榆(Cedrelospermum )[92 ] 、兔耳果(Lagokarpos )[93 ] 等在古近纪都可以通过青藏地区峡谷与周边动植物区系交流,新近纪之前的青藏地区虽然地形复杂,但是尚未隆升成今天的高原;横断山区某些区域则在晚始新世-早渐新世之交就已经获得现今的海拔高度[94 -95 ] ;高原北部和喜马拉雅山脉则在新近纪形成现在的地貌格局[25 ,96 -97 ] .古生物研究也发现青藏高原在晚上新世就已经存在高寒生境和冰缘环境,成为第四纪冰期动物忍耐寒冷环境的“训练营”,第四纪冰期来临之后这些适应寒冷环境的生物才扩散至北半球高纬度地区[34 ,98 -99 ] .因此,高原及周边地区高寒生物类群的起源时间表明在渐新世青藏高原部分区域就已经出现高寒生态系统,晚中新世以来青藏高原地貌与气候继续发生剧烈变化,促使这些物种的祖先类群迁入或就地演化积累适应高寒环境的基因和形态,并最终演化成适应高寒生境的新物种[25 ] .高寒区物种在晚上新世的加速分化可能与高原巨大山系和水系、冰川活动等造成的地理隔离有关[26 ] .随着第四纪冰期的到来,青藏高原及周边地区的高寒生物区也开始与北半球高纬度地区存在密切联系[34 ,98 -99 ] .同时,演化生物学研究也发现在第四纪冰期期间并未形成覆盖整个青藏高原的大冰盖,高原面上仍存在很多生物避难所[26 ] . ...
Evolutionary history of zoogeographical regions surrounding the Tibetan Plateau
1
2020
... 哺乳动物中的藏羚羊(Pantholops hodgsonii )生活于西藏、青海和新疆等地海拔3 700~5 500 m的区域,能够成功适应高原缺氧环境,其系统发育学研究表明在上新世-早更新世期间该类群就已经生活在青藏高原地区[65 ] .藏原羚(Procapra picticaudata )现今生活于海拔3 000~5 100 m地区,其种内分化时间约为4.4~2.2 Ma[66 ] .灵长目仰鼻猴属(Rhinopithecus )鼻孔结构是对高原缺氧环境的适应,系统发育研究表明在1.6 Ma左右其分布于四川和云南的金丝猴北方类群与分布于喜马拉雅的金丝猴类群发生分化[67 ] .兔形目鼠兔属(Ochotona )广泛分布于北半球草原和灌丛,其分布于青藏高原的类群演化出适应高寒生境的习性,在2.4 Ma左右,适应高寒生境类群与低海拔泛北极分布类群分化[68 ] .中华姬鼠(Apodemus draco )分布于青藏高原及周边地区以及湖南、湖北和台湾等地,其在3.8~0.87 Ma期间分化形成适应高海拔生境的物种,其种系及遗传结构由于高原阶段性隆升和冰川活动的影响而变得更加复杂[69 -70 ] .绒鼠属(Eothenomys )高原类群与低海拔类群分化时间约为2.7 Ma[71 ] .基于4 966种现生陆栖脊椎动物和1 278个哺乳动物化石属的研究也表明受印度-欧亚板块碰撞、高原阶段性隆升、亚洲内陆干旱化等事件的影响,青藏高原及其邻近地区动物区系在新生代发生显著变化,其区系成分在早期更接近东洋界,并最终在中新世-上新世之交形成与现在接近的动物区系格局[72 ] .对174种青藏高原特有陆栖脊椎动物祖先分布区、来源和分化时间的估算,发现青藏高原特有种早在55 Ma就开始从周边区域扩散至高原,扩散事件约从15 Ma开始增加,并在6 Ma之后达到顶峰,高原隆升和历史气候变化对该地区动物区系的形成与演化具有重要作用[73 ] . ...
Geological and climatic histories likely shaped the origins of terrestrial vertebrates endemic to the Tibetan Plateau
1
2021
... 哺乳动物中的藏羚羊(Pantholops hodgsonii )生活于西藏、青海和新疆等地海拔3 700~5 500 m的区域,能够成功适应高原缺氧环境,其系统发育学研究表明在上新世-早更新世期间该类群就已经生活在青藏高原地区[65 ] .藏原羚(Procapra picticaudata )现今生活于海拔3 000~5 100 m地区,其种内分化时间约为4.4~2.2 Ma[66 ] .灵长目仰鼻猴属(Rhinopithecus )鼻孔结构是对高原缺氧环境的适应,系统发育研究表明在1.6 Ma左右其分布于四川和云南的金丝猴北方类群与分布于喜马拉雅的金丝猴类群发生分化[67 ] .兔形目鼠兔属(Ochotona )广泛分布于北半球草原和灌丛,其分布于青藏高原的类群演化出适应高寒生境的习性,在2.4 Ma左右,适应高寒生境类群与低海拔泛北极分布类群分化[68 ] .中华姬鼠(Apodemus draco )分布于青藏高原及周边地区以及湖南、湖北和台湾等地,其在3.8~0.87 Ma期间分化形成适应高海拔生境的物种,其种系及遗传结构由于高原阶段性隆升和冰川活动的影响而变得更加复杂[69 -70 ] .绒鼠属(Eothenomys )高原类群与低海拔类群分化时间约为2.7 Ma[71 ] .基于4 966种现生陆栖脊椎动物和1 278个哺乳动物化石属的研究也表明受印度-欧亚板块碰撞、高原阶段性隆升、亚洲内陆干旱化等事件的影响,青藏高原及其邻近地区动物区系在新生代发生显著变化,其区系成分在早期更接近东洋界,并最终在中新世-上新世之交形成与现在接近的动物区系格局[72 ] .对174种青藏高原特有陆栖脊椎动物祖先分布区、来源和分化时间的估算,发现青藏高原特有种早在55 Ma就开始从周边区域扩散至高原,扩散事件约从15 Ma开始增加,并在6 Ma之后达到顶峰,高原隆升和历史气候变化对该地区动物区系的形成与演化具有重要作用[73 ] . ...
Origin and evolution of schizothoracine fishes in relation to the upheaval of the Xizang Plateau
2
1981
... 鲤形目裂腹鱼亚科(Schizothoracinae)鱼类是青藏高原及其周边地区的特有鱼类[74 ] ,地貌和水系格局的改变对裂腹鱼亚科的系统演化和分布产生深刻影响,而地质活动和气候事件制约着地貌和水系格局的形成和演化,因此可以通过裂腹鱼亚科系统演化、历史地理分布和化石解剖形态来推断地质活动和气候事件发生的时间,为青藏高原的隆升研究提供证据[74 -75 ] .裂腹鱼亚科系统发育研究表明在晚中新世晚期或上新世裂腹鱼亚科鱼类就已经出现在青藏高原及周边水系中,在晚更新世期间种内分化加速,不同海拔之间的类群产生等级分化,种间多样性增加,第四纪以来青藏高原隆升导致的水系改组是推动裂腹鱼亚科鱼类演化的原动力[31 ,76 -78 ] . ...
... [74 -75 ].裂腹鱼亚科系统发育研究表明在晚中新世晚期或上新世裂腹鱼亚科鱼类就已经出现在青藏高原及周边水系中,在晚更新世期间种内分化加速,不同海拔之间的类群产生等级分化,种间多样性增加,第四纪以来青藏高原隆升导致的水系改组是推动裂腹鱼亚科鱼类演化的原动力[31 ,76 -78 ] . ...
Origin and evolution of schizothoracine fishes in relation to the upheaval of the Xizang Plateau
2
1981
... 鲤形目裂腹鱼亚科(Schizothoracinae)鱼类是青藏高原及其周边地区的特有鱼类[74 ] ,地貌和水系格局的改变对裂腹鱼亚科的系统演化和分布产生深刻影响,而地质活动和气候事件制约着地貌和水系格局的形成和演化,因此可以通过裂腹鱼亚科系统演化、历史地理分布和化石解剖形态来推断地质活动和气候事件发生的时间,为青藏高原的隆升研究提供证据[74 -75 ] .裂腹鱼亚科系统发育研究表明在晚中新世晚期或上新世裂腹鱼亚科鱼类就已经出现在青藏高原及周边水系中,在晚更新世期间种内分化加速,不同海拔之间的类群产生等级分化,种间多样性增加,第四纪以来青藏高原隆升导致的水系改组是推动裂腹鱼亚科鱼类演化的原动力[31 ,76 -78 ] . ...
... [74 -75 ].裂腹鱼亚科系统发育研究表明在晚中新世晚期或上新世裂腹鱼亚科鱼类就已经出现在青藏高原及周边水系中,在晚更新世期间种内分化加速,不同海拔之间的类群产生等级分化,种间多样性增加,第四纪以来青藏高原隆升导致的水系改组是推动裂腹鱼亚科鱼类演化的原动力[31 ,76 -78 ] . ...
青藏高原的新生代鱼化石及其古环境意义
1
2016
... 鲤形目裂腹鱼亚科(Schizothoracinae)鱼类是青藏高原及其周边地区的特有鱼类[74 ] ,地貌和水系格局的改变对裂腹鱼亚科的系统演化和分布产生深刻影响,而地质活动和气候事件制约着地貌和水系格局的形成和演化,因此可以通过裂腹鱼亚科系统演化、历史地理分布和化石解剖形态来推断地质活动和气候事件发生的时间,为青藏高原的隆升研究提供证据[74 -75 ] .裂腹鱼亚科系统发育研究表明在晚中新世晚期或上新世裂腹鱼亚科鱼类就已经出现在青藏高原及周边水系中,在晚更新世期间种内分化加速,不同海拔之间的类群产生等级分化,种间多样性增加,第四纪以来青藏高原隆升导致的水系改组是推动裂腹鱼亚科鱼类演化的原动力[31 ,76 -78 ] . ...
青藏高原的新生代鱼化石及其古环境意义
1
2016
... 鲤形目裂腹鱼亚科(Schizothoracinae)鱼类是青藏高原及其周边地区的特有鱼类[74 ] ,地貌和水系格局的改变对裂腹鱼亚科的系统演化和分布产生深刻影响,而地质活动和气候事件制约着地貌和水系格局的形成和演化,因此可以通过裂腹鱼亚科系统演化、历史地理分布和化石解剖形态来推断地质活动和气候事件发生的时间,为青藏高原的隆升研究提供证据[74 -75 ] .裂腹鱼亚科系统发育研究表明在晚中新世晚期或上新世裂腹鱼亚科鱼类就已经出现在青藏高原及周边水系中,在晚更新世期间种内分化加速,不同海拔之间的类群产生等级分化,种间多样性增加,第四纪以来青藏高原隆升导致的水系改组是推动裂腹鱼亚科鱼类演化的原动力[31 ,76 -78 ] . ...
High intra-population genetic variability and inter-population differentiation in a plateau specialized fish, Triplophysa orientalis
2
2012
... 鲤形目裂腹鱼亚科(Schizothoracinae)鱼类是青藏高原及其周边地区的特有鱼类[74 ] ,地貌和水系格局的改变对裂腹鱼亚科的系统演化和分布产生深刻影响,而地质活动和气候事件制约着地貌和水系格局的形成和演化,因此可以通过裂腹鱼亚科系统演化、历史地理分布和化石解剖形态来推断地质活动和气候事件发生的时间,为青藏高原的隆升研究提供证据[74 -75 ] .裂腹鱼亚科系统发育研究表明在晚中新世晚期或上新世裂腹鱼亚科鱼类就已经出现在青藏高原及周边水系中,在晚更新世期间种内分化加速,不同海拔之间的类群产生等级分化,种间多样性增加,第四纪以来青藏高原隆升导致的水系改组是推动裂腹鱼亚科鱼类演化的原动力[31 ,76 -78 ] . ...
... Biological evolution in the alpine region of Tibetan Plateau
Table 1 种属 分布区域 适应高海拔类群分化时间或多样性增加时间 文献来源 Sinopodophyllum hexandrum 青藏高原东部 在约(6.52±1.89) Ma期间出现在青藏高原东部,但是直到3~2 Ma以来才演化出适应高山环境的形态 [47 ] Nannoglottis 青藏高原 在23~32 Ma期间出现在青藏高原,随后多样性增高,其高山灌丛和针叶林生活类型分支时间约为3.4 Ma [48 ] Reaumuria soongarica 西北干旱区、青藏高原北部 高原内部分布类群演化时间2.0~6.3 Ma,高原东北缘与高原西北类群分化时间1.1~3.5 Ma [49 ] Orchidinae 青藏高原及喜马拉雅山脉 青藏高原特有种大多是在过去6 Ma里进化而来 [50 ] Oxyria sinensis 青藏高原东南部 高山类群出现时间7.54~24.36 Ma,种内多样化增加时间1.74~0.86 Ma [51 ] Juniperus microsperma 西藏波密 早中新世21.6~14 Ma期间与其他种群分化 [52 ] Rhododendron 北半球 横断山区概属多样性在晚中新世(约8 Ma)快速增加 [53 ] Roscoea, Cautleya 喜马拉雅-横断山区 在早渐新世(~32 Ma)两属出现,Roscoea在横断山区与喜马拉雅山脉的两个分支大概在渐新世-中新世之交分化 [54 ] Nanorana parkeri 青藏高原南部 高原东部和西部类群分化时间1.4~3.7 Ma [55 ] Paini 南亚、东南亚、青藏高原 在渐新世早期棘蛙族高原-喜马拉雅类群与中国第二阶梯上生活的类群分化,随后在在26.1~12.5 Ma高原生活类群与喜马拉雅山脉生活类群分化,高原内部生活类群在晚中新世到上新世期间分化速率加快 [22 ] Bufo tibetanus 青藏高原、四川、甘肃、陕西等 高原种类形成于上新世期间 [56 ] Phrynocephalus 亚洲干旱区 在7.2~13.0 Ma期间演化出胎生类群,3.2~7.3 Ma期间种间多样性增加 [57 ] Phrynocephalus erythrurus 羌塘盆地 在3.67 Ma左右北部类群与南部类群发生分化,约2.76 Ma北部类群内部进一步分化 [58 ] Gnaptorina 青藏高原及周边高海拔地区 始新世起源于青藏高原东南缘 [18 -19 ] Coelotinae 北半球温带、亚热带 晚始新世高原东南缘隆升阻碍南方隙蛛向北扩散,渐新世高原中部和北部隆起导致北方隙蛛早期多样化进程中的灭绝事件 [59 ] Montifringilla, Onychostruthus, Pyrgilauda 青藏高原 雪雀的共同祖先在约14 Ma就应经生活在青藏高原,并进化出适应高海拔极端环境的性状 [21 ] Parus major 亚洲、欧洲 蒙古分布类群与其他种类分化时间为2.8~0.7 Ma,喜马拉雅山脉东部与中国中东部类群分化时间为1.9~0.4 Ma [60 ] Pseudopodoces humilis 青藏高原、柴达木盆地周边高山 在9.9~7.7 Ma期间地山雀就与与其他山雀分化开来,并出现适应高海拔性状 [61 ] Phylloscopus, Seicercus 亚洲 喜马拉雅山脉分布类群在7 Ma加速进化 [62 -63 ] Perdix 西藏、甘肃、青海、四川、新疆 高原类群出现于3.63 (4.52~2.75) Ma [64 ] Pantholops hodgsonii 西藏、新疆、青海 上新世-更新世就已经出现在青藏高原 [65 ] Procapra picticaudata 西藏、四川、甘肃、青海、新疆 种内分化时间约2.2~4.4 Ma [66 ] Rhinopithecus 四川、甘肃、陕西、贵州、云南、缅甸、越南 北方类群川金丝猴、黔金丝猴与喜马拉雅类群滇金丝猴、缅甸金丝猴分化时间约1.6 Ma [67 ] Ochotona 北半球 分布于青藏高原的高山类群和灌丛草原类群与低海拔泛北极分布类群分化时间大约为2.4 Ma [68 ] Apodemus draco 四川、云南、甘肃、青海、陕西、湖北、湖南、福建、台湾 2.2 Ma (3.8~0.87 Ma)分化形成,后来由于高原的阶段性隆升以及冰川作用种系及遗传结构更加复杂 [69 -70 ] Eothenomys 东亚 高原种类与低海拔种类分裂时间约2.7 Ma [71 ] Schizothoracinae 青藏高原及周边地区 在晚中新世晚期或上新世裂腹鱼亚科鱼类就已经出现在青藏高原及周边水系中,在晚更新世期间种内分化加速,不同海拔之间的类群产生等级分化,种间多样性增加,第四纪以来青藏高原隆升导致的水系改组是推动裂腹鱼亚科鱼类演化的原动力 [31 ,76 -78 ]
3 讨论 最近,青藏高原古地貌重建工作取得了丰硕的成果(图3 ),但是各种指标重建结果往往存在矛盾(图4 )[25 ,79 -83 ] .同位素重建结果表明高原南部在古近纪早期就存在安第斯型的巨大山系[84 -85 ] ;古生物研究发现高原腹地从中始新世到晚渐新世都存在东西向的大峡谷,该峡谷底部海拔在2 000 m左右,两侧为高大的冈底斯山脉和羌塘山脉[82 ,86 -88 ] ,动植物如巨犀(Paraceratherium )[89 ] 、攀鲈(Eoanabas )[90 -91 ] 、臭椿(Ailanthus )[88 ] 、椿榆(Cedrelospermum )[92 ] 、兔耳果(Lagokarpos )[93 ] 等在古近纪都可以通过青藏地区峡谷与周边动植物区系交流,新近纪之前的青藏地区虽然地形复杂,但是尚未隆升成今天的高原;横断山区某些区域则在晚始新世-早渐新世之交就已经获得现今的海拔高度[94 -95 ] ;高原北部和喜马拉雅山脉则在新近纪形成现在的地貌格局[25 ,96 -97 ] .古生物研究也发现青藏高原在晚上新世就已经存在高寒生境和冰缘环境,成为第四纪冰期动物忍耐寒冷环境的“训练营”,第四纪冰期来临之后这些适应寒冷环境的生物才扩散至北半球高纬度地区[34 ,98 -99 ] .因此,高原及周边地区高寒生物类群的起源时间表明在渐新世青藏高原部分区域就已经出现高寒生态系统,晚中新世以来青藏高原地貌与气候继续发生剧烈变化,促使这些物种的祖先类群迁入或就地演化积累适应高寒环境的基因和形态,并最终演化成适应高寒生境的新物种[25 ] .高寒区物种在晚上新世的加速分化可能与高原巨大山系和水系、冰川活动等造成的地理隔离有关[26 ] .随着第四纪冰期的到来,青藏高原及周边地区的高寒生物区也开始与北半球高纬度地区存在密切联系[34 ,98 -99 ] .同时,演化生物学研究也发现在第四纪冰期期间并未形成覆盖整个青藏高原的大冰盖,高原面上仍存在很多生物避难所[26 ] . ...
Strong population structure of Schizopygopsis chengi and the origin of S. chengi baoxingensis revealed by mtDNA and microsatellite markers
0
2015
The biogeography and phylogeny of schizothoracine fishes (Schizopygopsis ) in the Qinghai‐Tibetan Plateau
2
2015
... 鲤形目裂腹鱼亚科(Schizothoracinae)鱼类是青藏高原及其周边地区的特有鱼类[74 ] ,地貌和水系格局的改变对裂腹鱼亚科的系统演化和分布产生深刻影响,而地质活动和气候事件制约着地貌和水系格局的形成和演化,因此可以通过裂腹鱼亚科系统演化、历史地理分布和化石解剖形态来推断地质活动和气候事件发生的时间,为青藏高原的隆升研究提供证据[74 -75 ] .裂腹鱼亚科系统发育研究表明在晚中新世晚期或上新世裂腹鱼亚科鱼类就已经出现在青藏高原及周边水系中,在晚更新世期间种内分化加速,不同海拔之间的类群产生等级分化,种间多样性增加,第四纪以来青藏高原隆升导致的水系改组是推动裂腹鱼亚科鱼类演化的原动力[31 ,76 -78 ] . ...
... Biological evolution in the alpine region of Tibetan Plateau
Table 1 种属 分布区域 适应高海拔类群分化时间或多样性增加时间 文献来源 Sinopodophyllum hexandrum 青藏高原东部 在约(6.52±1.89) Ma期间出现在青藏高原东部,但是直到3~2 Ma以来才演化出适应高山环境的形态 [47 ] Nannoglottis 青藏高原 在23~32 Ma期间出现在青藏高原,随后多样性增高,其高山灌丛和针叶林生活类型分支时间约为3.4 Ma [48 ] Reaumuria soongarica 西北干旱区、青藏高原北部 高原内部分布类群演化时间2.0~6.3 Ma,高原东北缘与高原西北类群分化时间1.1~3.5 Ma [49 ] Orchidinae 青藏高原及喜马拉雅山脉 青藏高原特有种大多是在过去6 Ma里进化而来 [50 ] Oxyria sinensis 青藏高原东南部 高山类群出现时间7.54~24.36 Ma,种内多样化增加时间1.74~0.86 Ma [51 ] Juniperus microsperma 西藏波密 早中新世21.6~14 Ma期间与其他种群分化 [52 ] Rhododendron 北半球 横断山区概属多样性在晚中新世(约8 Ma)快速增加 [53 ] Roscoea, Cautleya 喜马拉雅-横断山区 在早渐新世(~32 Ma)两属出现,Roscoea在横断山区与喜马拉雅山脉的两个分支大概在渐新世-中新世之交分化 [54 ] Nanorana parkeri 青藏高原南部 高原东部和西部类群分化时间1.4~3.7 Ma [55 ] Paini 南亚、东南亚、青藏高原 在渐新世早期棘蛙族高原-喜马拉雅类群与中国第二阶梯上生活的类群分化,随后在在26.1~12.5 Ma高原生活类群与喜马拉雅山脉生活类群分化,高原内部生活类群在晚中新世到上新世期间分化速率加快 [22 ] Bufo tibetanus 青藏高原、四川、甘肃、陕西等 高原种类形成于上新世期间 [56 ] Phrynocephalus 亚洲干旱区 在7.2~13.0 Ma期间演化出胎生类群,3.2~7.3 Ma期间种间多样性增加 [57 ] Phrynocephalus erythrurus 羌塘盆地 在3.67 Ma左右北部类群与南部类群发生分化,约2.76 Ma北部类群内部进一步分化 [58 ] Gnaptorina 青藏高原及周边高海拔地区 始新世起源于青藏高原东南缘 [18 -19 ] Coelotinae 北半球温带、亚热带 晚始新世高原东南缘隆升阻碍南方隙蛛向北扩散,渐新世高原中部和北部隆起导致北方隙蛛早期多样化进程中的灭绝事件 [59 ] Montifringilla, Onychostruthus, Pyrgilauda 青藏高原 雪雀的共同祖先在约14 Ma就应经生活在青藏高原,并进化出适应高海拔极端环境的性状 [21 ] Parus major 亚洲、欧洲 蒙古分布类群与其他种类分化时间为2.8~0.7 Ma,喜马拉雅山脉东部与中国中东部类群分化时间为1.9~0.4 Ma [60 ] Pseudopodoces humilis 青藏高原、柴达木盆地周边高山 在9.9~7.7 Ma期间地山雀就与与其他山雀分化开来,并出现适应高海拔性状 [61 ] Phylloscopus, Seicercus 亚洲 喜马拉雅山脉分布类群在7 Ma加速进化 [62 -63 ] Perdix 西藏、甘肃、青海、四川、新疆 高原类群出现于3.63 (4.52~2.75) Ma [64 ] Pantholops hodgsonii 西藏、新疆、青海 上新世-更新世就已经出现在青藏高原 [65 ] Procapra picticaudata 西藏、四川、甘肃、青海、新疆 种内分化时间约2.2~4.4 Ma [66 ] Rhinopithecus 四川、甘肃、陕西、贵州、云南、缅甸、越南 北方类群川金丝猴、黔金丝猴与喜马拉雅类群滇金丝猴、缅甸金丝猴分化时间约1.6 Ma [67 ] Ochotona 北半球 分布于青藏高原的高山类群和灌丛草原类群与低海拔泛北极分布类群分化时间大约为2.4 Ma [68 ] Apodemus draco 四川、云南、甘肃、青海、陕西、湖北、湖南、福建、台湾 2.2 Ma (3.8~0.87 Ma)分化形成,后来由于高原的阶段性隆升以及冰川作用种系及遗传结构更加复杂 [69 -70 ] Eothenomys 东亚 高原种类与低海拔种类分裂时间约2.7 Ma [71 ] Schizothoracinae 青藏高原及周边地区 在晚中新世晚期或上新世裂腹鱼亚科鱼类就已经出现在青藏高原及周边水系中,在晚更新世期间种内分化加速,不同海拔之间的类群产生等级分化,种间多样性增加,第四纪以来青藏高原隆升导致的水系改组是推动裂腹鱼亚科鱼类演化的原动力 [31 ,76 -78 ]
3 讨论 最近,青藏高原古地貌重建工作取得了丰硕的成果(图3 ),但是各种指标重建结果往往存在矛盾(图4 )[25 ,79 -83 ] .同位素重建结果表明高原南部在古近纪早期就存在安第斯型的巨大山系[84 -85 ] ;古生物研究发现高原腹地从中始新世到晚渐新世都存在东西向的大峡谷,该峡谷底部海拔在2 000 m左右,两侧为高大的冈底斯山脉和羌塘山脉[82 ,86 -88 ] ,动植物如巨犀(Paraceratherium )[89 ] 、攀鲈(Eoanabas )[90 -91 ] 、臭椿(Ailanthus )[88 ] 、椿榆(Cedrelospermum )[92 ] 、兔耳果(Lagokarpos )[93 ] 等在古近纪都可以通过青藏地区峡谷与周边动植物区系交流,新近纪之前的青藏地区虽然地形复杂,但是尚未隆升成今天的高原;横断山区某些区域则在晚始新世-早渐新世之交就已经获得现今的海拔高度[94 -95 ] ;高原北部和喜马拉雅山脉则在新近纪形成现在的地貌格局[25 ,96 -97 ] .古生物研究也发现青藏高原在晚上新世就已经存在高寒生境和冰缘环境,成为第四纪冰期动物忍耐寒冷环境的“训练营”,第四纪冰期来临之后这些适应寒冷环境的生物才扩散至北半球高纬度地区[34 ,98 -99 ] .因此,高原及周边地区高寒生物类群的起源时间表明在渐新世青藏高原部分区域就已经出现高寒生态系统,晚中新世以来青藏高原地貌与气候继续发生剧烈变化,促使这些物种的祖先类群迁入或就地演化积累适应高寒环境的基因和形态,并最终演化成适应高寒生境的新物种[25 ] .高寒区物种在晚上新世的加速分化可能与高原巨大山系和水系、冰川活动等造成的地理隔离有关[26 ] .随着第四纪冰期的到来,青藏高原及周边地区的高寒生物区也开始与北半球高纬度地区存在密切联系[34 ,98 -99 ] .同时,演化生物学研究也发现在第四纪冰期期间并未形成覆盖整个青藏高原的大冰盖,高原面上仍存在很多生物避难所[26 ] . ...
Revised paleoaltimetry data show low Tibetan Plateau elevation during the Eocene
1
2019
... 最近,青藏高原古地貌重建工作取得了丰硕的成果(图3 ),但是各种指标重建结果往往存在矛盾(图4 )[25 ,79 -83 ] .同位素重建结果表明高原南部在古近纪早期就存在安第斯型的巨大山系[84 -85 ] ;古生物研究发现高原腹地从中始新世到晚渐新世都存在东西向的大峡谷,该峡谷底部海拔在2 000 m左右,两侧为高大的冈底斯山脉和羌塘山脉[82 ,86 -88 ] ,动植物如巨犀(Paraceratherium )[89 ] 、攀鲈(Eoanabas )[90 -91 ] 、臭椿(Ailanthus )[88 ] 、椿榆(Cedrelospermum )[92 ] 、兔耳果(Lagokarpos )[93 ] 等在古近纪都可以通过青藏地区峡谷与周边动植物区系交流,新近纪之前的青藏地区虽然地形复杂,但是尚未隆升成今天的高原;横断山区某些区域则在晚始新世-早渐新世之交就已经获得现今的海拔高度[94 -95 ] ;高原北部和喜马拉雅山脉则在新近纪形成现在的地貌格局[25 ,96 -97 ] .古生物研究也发现青藏高原在晚上新世就已经存在高寒生境和冰缘环境,成为第四纪冰期动物忍耐寒冷环境的“训练营”,第四纪冰期来临之后这些适应寒冷环境的生物才扩散至北半球高纬度地区[34 ,98 -99 ] .因此,高原及周边地区高寒生物类群的起源时间表明在渐新世青藏高原部分区域就已经出现高寒生态系统,晚中新世以来青藏高原地貌与气候继续发生剧烈变化,促使这些物种的祖先类群迁入或就地演化积累适应高寒环境的基因和形态,并最终演化成适应高寒生境的新物种[25 ] .高寒区物种在晚上新世的加速分化可能与高原巨大山系和水系、冰川活动等造成的地理隔离有关[26 ] .随着第四纪冰期的到来,青藏高原及周边地区的高寒生物区也开始与北半球高纬度地区存在密切联系[34 ,98 -99 ] .同时,演化生物学研究也发现在第四纪冰期期间并未形成覆盖整个青藏高原的大冰盖,高原面上仍存在很多生物避难所[26 ] . ...
Palaeo-altimetry of the late Eocene to Miocene Lunpola basin, central Tibet
0
2006
Why the ‘Uplift of the Tibetan Plateau’ is a myth
3
2021
... 随着DNA测序技术和比较系统学方法的发展,大量涌现的青藏高寒区生物地理学研究为我们理解高原隆升过程及其气候效应提供了生物演化方面的证据[25 ] .但是,演化生物学家通常仅对单一特定类群的系统发育过程开展研究,很少进行系统的总结工作[25 ] ,对某一类群遗传数据的解释也往往与最新的青藏高原隆升和气候变化研究进展脱节,不能很好地将最新的关于高原隆升和气候变化的研究与生物类群演化联系在一起[101 ] ,同时也没有认识到青藏高原的隆升过程是阶段性的,没有对青藏高原的不同组成部分进行区分[26 ,53 ,81 ] .地质学研究大多认为高原地貌格局形成于中新世之前,但是综合近年来的生物地理学研究结果可以发现,青藏高寒生物类群的分化时间多集中在晚中新世,种内分化加速和多样性增加主要发生在晚上新世到第四纪期间(图5 ),这些不同类群在分化时间上的巧合可能反映青藏高寒区生物演化受到晚中新世以来构造和气候事件的影响,但是并未引起地质学家足够的重视[25 ] . ...
... 注: 青藏高原隆升示意图从左到右依次为古近纪早期高原南部安第斯型山脉,古近纪中晚期高原中部东西向峡谷模型,新近纪早期高原中部和南部隆升,新近纪晚期高原北部生长[81 -82 ,84 -85 ,87 ,94 ,96 -97 ] ...
... Note: The tectonic history of the Tibetan Plateau shows the Andean-type mountains in the southern plateau in the early Paleogene, the east-west canyon model in the middle plateau until the late Paleogene, the uplift of the central and southern plateau in the early Neogene, and the growth of the northern plateau during the late Neogene[81 -82 ,84 -85 ,87 ,94 ,96 -97 ] ...
No high Tibetan Plateau until the Neogene
4
2019
... 最近,青藏高原古地貌重建工作取得了丰硕的成果(图3 ),但是各种指标重建结果往往存在矛盾(图4 )[25 ,79 -83 ] .同位素重建结果表明高原南部在古近纪早期就存在安第斯型的巨大山系[84 -85 ] ;古生物研究发现高原腹地从中始新世到晚渐新世都存在东西向的大峡谷,该峡谷底部海拔在2 000 m左右,两侧为高大的冈底斯山脉和羌塘山脉[82 ,86 -88 ] ,动植物如巨犀(Paraceratherium )[89 ] 、攀鲈(Eoanabas )[90 -91 ] 、臭椿(Ailanthus )[88 ] 、椿榆(Cedrelospermum )[92 ] 、兔耳果(Lagokarpos )[93 ] 等在古近纪都可以通过青藏地区峡谷与周边动植物区系交流,新近纪之前的青藏地区虽然地形复杂,但是尚未隆升成今天的高原;横断山区某些区域则在晚始新世-早渐新世之交就已经获得现今的海拔高度[94 -95 ] ;高原北部和喜马拉雅山脉则在新近纪形成现在的地貌格局[25 ,96 -97 ] .古生物研究也发现青藏高原在晚上新世就已经存在高寒生境和冰缘环境,成为第四纪冰期动物忍耐寒冷环境的“训练营”,第四纪冰期来临之后这些适应寒冷环境的生物才扩散至北半球高纬度地区[34 ,98 -99 ] .因此,高原及周边地区高寒生物类群的起源时间表明在渐新世青藏高原部分区域就已经出现高寒生态系统,晚中新世以来青藏高原地貌与气候继续发生剧烈变化,促使这些物种的祖先类群迁入或就地演化积累适应高寒环境的基因和形态,并最终演化成适应高寒生境的新物种[25 ] .高寒区物种在晚上新世的加速分化可能与高原巨大山系和水系、冰川活动等造成的地理隔离有关[26 ] .随着第四纪冰期的到来,青藏高原及周边地区的高寒生物区也开始与北半球高纬度地区存在密切联系[34 ,98 -99 ] .同时,演化生物学研究也发现在第四纪冰期期间并未形成覆盖整个青藏高原的大冰盖,高原面上仍存在很多生物避难所[26 ] . ...
... ]、[
82 ]、[
87 ]、[
100 ]修改)
North-south paleoelevation profiles on the Tibetan Plateau (Revised from References [25],[34],[82],[87],[100]) Fig.4 ![]()
随着DNA测序技术和比较系统学方法的发展,大量涌现的青藏高寒区生物地理学研究为我们理解高原隆升过程及其气候效应提供了生物演化方面的证据[25 ] .但是,演化生物学家通常仅对单一特定类群的系统发育过程开展研究,很少进行系统的总结工作[25 ] ,对某一类群遗传数据的解释也往往与最新的青藏高原隆升和气候变化研究进展脱节,不能很好地将最新的关于高原隆升和气候变化的研究与生物类群演化联系在一起[101 ] ,同时也没有认识到青藏高原的隆升过程是阶段性的,没有对青藏高原的不同组成部分进行区分[26 ,53 ,81 ] .地质学研究大多认为高原地貌格局形成于中新世之前,但是综合近年来的生物地理学研究结果可以发现,青藏高寒生物类群的分化时间多集中在晚中新世,种内分化加速和多样性增加主要发生在晚上新世到第四纪期间(图5 ),这些不同类群在分化时间上的巧合可能反映青藏高寒区生物演化受到晚中新世以来构造和气候事件的影响,但是并未引起地质学家足够的重视[25 ] . ...
... 注: 青藏高原隆升示意图从左到右依次为古近纪早期高原南部安第斯型山脉,古近纪中晚期高原中部东西向峡谷模型,新近纪早期高原中部和南部隆升,新近纪晚期高原北部生长[81 -82 ,84 -85 ,87 ,94 ,96 -97 ] ...
... Note: The tectonic history of the Tibetan Plateau shows the Andean-type mountains in the southern plateau in the early Paleogene, the east-west canyon model in the middle plateau until the late Paleogene, the uplift of the central and southern plateau in the early Neogene, and the growth of the northern plateau during the late Neogene[81 -82 ,84 -85 ,87 ,94 ,96 -97 ] ...
Comment on “Revised paleoaltimetry data show low Tibetan Plateau elevation during the Eocene”
1
2019
... 最近,青藏高原古地貌重建工作取得了丰硕的成果(图3 ),但是各种指标重建结果往往存在矛盾(图4 )[25 ,79 -83 ] .同位素重建结果表明高原南部在古近纪早期就存在安第斯型的巨大山系[84 -85 ] ;古生物研究发现高原腹地从中始新世到晚渐新世都存在东西向的大峡谷,该峡谷底部海拔在2 000 m左右,两侧为高大的冈底斯山脉和羌塘山脉[82 ,86 -88 ] ,动植物如巨犀(Paraceratherium )[89 ] 、攀鲈(Eoanabas )[90 -91 ] 、臭椿(Ailanthus )[88 ] 、椿榆(Cedrelospermum )[92 ] 、兔耳果(Lagokarpos )[93 ] 等在古近纪都可以通过青藏地区峡谷与周边动植物区系交流,新近纪之前的青藏地区虽然地形复杂,但是尚未隆升成今天的高原;横断山区某些区域则在晚始新世-早渐新世之交就已经获得现今的海拔高度[94 -95 ] ;高原北部和喜马拉雅山脉则在新近纪形成现在的地貌格局[25 ,96 -97 ] .古生物研究也发现青藏高原在晚上新世就已经存在高寒生境和冰缘环境,成为第四纪冰期动物忍耐寒冷环境的“训练营”,第四纪冰期来临之后这些适应寒冷环境的生物才扩散至北半球高纬度地区[34 ,98 -99 ] .因此,高原及周边地区高寒生物类群的起源时间表明在渐新世青藏高原部分区域就已经出现高寒生态系统,晚中新世以来青藏高原地貌与气候继续发生剧烈变化,促使这些物种的祖先类群迁入或就地演化积累适应高寒环境的基因和形态,并最终演化成适应高寒生境的新物种[25 ] .高寒区物种在晚上新世的加速分化可能与高原巨大山系和水系、冰川活动等造成的地理隔离有关[26 ] .随着第四纪冰期的到来,青藏高原及周边地区的高寒生物区也开始与北半球高纬度地区存在密切联系[34 ,98 -99 ] .同时,演化生物学研究也发现在第四纪冰期期间并未形成覆盖整个青藏高原的大冰盖,高原面上仍存在很多生物避难所[26 ] . ...
Quantifying the rise of the Himalaya orogen and implications for the South Asian monsoon
3
2017
... 最近,青藏高原古地貌重建工作取得了丰硕的成果(图3 ),但是各种指标重建结果往往存在矛盾(图4 )[25 ,79 -83 ] .同位素重建结果表明高原南部在古近纪早期就存在安第斯型的巨大山系[84 -85 ] ;古生物研究发现高原腹地从中始新世到晚渐新世都存在东西向的大峡谷,该峡谷底部海拔在2 000 m左右,两侧为高大的冈底斯山脉和羌塘山脉[82 ,86 -88 ] ,动植物如巨犀(Paraceratherium )[89 ] 、攀鲈(Eoanabas )[90 -91 ] 、臭椿(Ailanthus )[88 ] 、椿榆(Cedrelospermum )[92 ] 、兔耳果(Lagokarpos )[93 ] 等在古近纪都可以通过青藏地区峡谷与周边动植物区系交流,新近纪之前的青藏地区虽然地形复杂,但是尚未隆升成今天的高原;横断山区某些区域则在晚始新世-早渐新世之交就已经获得现今的海拔高度[94 -95 ] ;高原北部和喜马拉雅山脉则在新近纪形成现在的地貌格局[25 ,96 -97 ] .古生物研究也发现青藏高原在晚上新世就已经存在高寒生境和冰缘环境,成为第四纪冰期动物忍耐寒冷环境的“训练营”,第四纪冰期来临之后这些适应寒冷环境的生物才扩散至北半球高纬度地区[34 ,98 -99 ] .因此,高原及周边地区高寒生物类群的起源时间表明在渐新世青藏高原部分区域就已经出现高寒生态系统,晚中新世以来青藏高原地貌与气候继续发生剧烈变化,促使这些物种的祖先类群迁入或就地演化积累适应高寒环境的基因和形态,并最终演化成适应高寒生境的新物种[25 ] .高寒区物种在晚上新世的加速分化可能与高原巨大山系和水系、冰川活动等造成的地理隔离有关[26 ] .随着第四纪冰期的到来,青藏高原及周边地区的高寒生物区也开始与北半球高纬度地区存在密切联系[34 ,98 -99 ] .同时,演化生物学研究也发现在第四纪冰期期间并未形成覆盖整个青藏高原的大冰盖,高原面上仍存在很多生物避难所[26 ] . ...
... 注: 青藏高原隆升示意图从左到右依次为古近纪早期高原南部安第斯型山脉,古近纪中晚期高原中部东西向峡谷模型,新近纪早期高原中部和南部隆升,新近纪晚期高原北部生长[81 -82 ,84 -85 ,87 ,94 ,96 -97 ] ...
... Note: The tectonic history of the Tibetan Plateau shows the Andean-type mountains in the southern plateau in the early Paleogene, the east-west canyon model in the middle plateau until the late Paleogene, the uplift of the central and southern plateau in the early Neogene, and the growth of the northern plateau during the late Neogene[81 -82 ,84 -85 ,87 ,94 ,96 -97 ] ...
The Andean-type Gangdese Mountains: paleoelevation record from the Paleocene-Eocene Linzhou Basin
3
2014
... 最近,青藏高原古地貌重建工作取得了丰硕的成果(图3 ),但是各种指标重建结果往往存在矛盾(图4 )[25 ,79 -83 ] .同位素重建结果表明高原南部在古近纪早期就存在安第斯型的巨大山系[84 -85 ] ;古生物研究发现高原腹地从中始新世到晚渐新世都存在东西向的大峡谷,该峡谷底部海拔在2 000 m左右,两侧为高大的冈底斯山脉和羌塘山脉[82 ,86 -88 ] ,动植物如巨犀(Paraceratherium )[89 ] 、攀鲈(Eoanabas )[90 -91 ] 、臭椿(Ailanthus )[88 ] 、椿榆(Cedrelospermum )[92 ] 、兔耳果(Lagokarpos )[93 ] 等在古近纪都可以通过青藏地区峡谷与周边动植物区系交流,新近纪之前的青藏地区虽然地形复杂,但是尚未隆升成今天的高原;横断山区某些区域则在晚始新世-早渐新世之交就已经获得现今的海拔高度[94 -95 ] ;高原北部和喜马拉雅山脉则在新近纪形成现在的地貌格局[25 ,96 -97 ] .古生物研究也发现青藏高原在晚上新世就已经存在高寒生境和冰缘环境,成为第四纪冰期动物忍耐寒冷环境的“训练营”,第四纪冰期来临之后这些适应寒冷环境的生物才扩散至北半球高纬度地区[34 ,98 -99 ] .因此,高原及周边地区高寒生物类群的起源时间表明在渐新世青藏高原部分区域就已经出现高寒生态系统,晚中新世以来青藏高原地貌与气候继续发生剧烈变化,促使这些物种的祖先类群迁入或就地演化积累适应高寒环境的基因和形态,并最终演化成适应高寒生境的新物种[25 ] .高寒区物种在晚上新世的加速分化可能与高原巨大山系和水系、冰川活动等造成的地理隔离有关[26 ] .随着第四纪冰期的到来,青藏高原及周边地区的高寒生物区也开始与北半球高纬度地区存在密切联系[34 ,98 -99 ] .同时,演化生物学研究也发现在第四纪冰期期间并未形成覆盖整个青藏高原的大冰盖,高原面上仍存在很多生物避难所[26 ] . ...
... 注: 青藏高原隆升示意图从左到右依次为古近纪早期高原南部安第斯型山脉,古近纪中晚期高原中部东西向峡谷模型,新近纪早期高原中部和南部隆升,新近纪晚期高原北部生长[81 -82 ,84 -85 ,87 ,94 ,96 -97 ] ...
... Note: The tectonic history of the Tibetan Plateau shows the Andean-type mountains in the southern plateau in the early Paleogene, the east-west canyon model in the middle plateau until the late Paleogene, the uplift of the central and southern plateau in the early Neogene, and the growth of the northern plateau during the late Neogene[81 -82 ,84 -85 ,87 ,94 ,96 -97 ] ...
A Middle Eocene lowland humid subtropical “Shangri-La” ecosystem in central Tibet
1
2020
... 最近,青藏高原古地貌重建工作取得了丰硕的成果(图3 ),但是各种指标重建结果往往存在矛盾(图4 )[25 ,79 -83 ] .同位素重建结果表明高原南部在古近纪早期就存在安第斯型的巨大山系[84 -85 ] ;古生物研究发现高原腹地从中始新世到晚渐新世都存在东西向的大峡谷,该峡谷底部海拔在2 000 m左右,两侧为高大的冈底斯山脉和羌塘山脉[82 ,86 -88 ] ,动植物如巨犀(Paraceratherium )[89 ] 、攀鲈(Eoanabas )[90 -91 ] 、臭椿(Ailanthus )[88 ] 、椿榆(Cedrelospermum )[92 ] 、兔耳果(Lagokarpos )[93 ] 等在古近纪都可以通过青藏地区峡谷与周边动植物区系交流,新近纪之前的青藏地区虽然地形复杂,但是尚未隆升成今天的高原;横断山区某些区域则在晚始新世-早渐新世之交就已经获得现今的海拔高度[94 -95 ] ;高原北部和喜马拉雅山脉则在新近纪形成现在的地貌格局[25 ,96 -97 ] .古生物研究也发现青藏高原在晚上新世就已经存在高寒生境和冰缘环境,成为第四纪冰期动物忍耐寒冷环境的“训练营”,第四纪冰期来临之后这些适应寒冷环境的生物才扩散至北半球高纬度地区[34 ,98 -99 ] .因此,高原及周边地区高寒生物类群的起源时间表明在渐新世青藏高原部分区域就已经出现高寒生态系统,晚中新世以来青藏高原地貌与气候继续发生剧烈变化,促使这些物种的祖先类群迁入或就地演化积累适应高寒环境的基因和形态,并最终演化成适应高寒生境的新物种[25 ] .高寒区物种在晚上新世的加速分化可能与高原巨大山系和水系、冰川活动等造成的地理隔离有关[26 ] .随着第四纪冰期的到来,青藏高原及周边地区的高寒生物区也开始与北半球高纬度地区存在密切联系[34 ,98 -99 ] .同时,演化生物学研究也发现在第四纪冰期期间并未形成覆盖整个青藏高原的大冰盖,高原面上仍存在很多生物避难所[26 ] . ...
Revised chronology of central Tibet uplift (Lunpola Basin)
3
2020
... 最近,青藏高原古地貌重建工作取得了丰硕的成果(
图3 ),但是各种指标重建结果往往存在矛盾(
图4 )
[25 ,79 -83 ] .同位素重建结果表明高原南部在古近纪早期就存在安第斯型的巨大山系
[84 -85 ] ;古生物研究发现高原腹地从中始新世到晚渐新世都存在东西向的大峡谷,该峡谷底部海拔在2 000 m左右,两侧为高大的冈底斯山脉和羌塘山脉
[82 ,86 -88 ] ,动植物如巨犀(
Paraceratherium )
[89 ] 、攀鲈(
Eoanabas )
[90 -91 ] 、臭椿(
Ailanthus )
[88 ] 、椿榆(
Cedrelospermum )
[92 ] 、兔耳果(
Lagokarpos )
[93 ] 等在古近纪都可以通过青藏地区峡谷与周边动植物区系交流,新近纪之前的青藏地区虽然地形复杂,但是尚未隆升成今天的高原;横断山区某些区域则在晚始新世-早渐新世之交就已经获得现今的海拔高度
[94 -95 ] ;高原北部和喜马拉雅山脉则在新近纪形成现在的地貌格局
[25 ,96 -97 ] .古生物研究也发现青藏高原在晚上新世就已经存在高寒生境和冰缘环境,成为第四纪冰期动物忍耐寒冷环境的“训练营”,第四纪冰期来临之后这些适应寒冷环境的生物才扩散至北半球高纬度地区
[34 ,98 -99 ] .因此,高原及周边地区高寒生物类群的起源时间表明在渐新世青藏高原部分区域就已经出现高寒生态系统,晚中新世以来青藏高原地貌与气候继续发生剧烈变化,促使这些物种的祖先类群迁入或就地演化积累适应高寒环境的基因和形态,并最终演化成适应高寒生境的新物种
[25 ] .高寒区物种在晚上新世的加速分化可能与高原巨大山系和水系、冰川活动等造成的地理隔离有关
[26 ] .随着第四纪冰期的到来,青藏高原及周边地区的高寒生物区也开始与北半球高纬度地区存在密切联系
[34 ,98 -99 ] .同时,演化生物学研究也发现在第四纪冰期期间并未形成覆盖整个青藏高原的大冰盖,高原面上仍存在很多生物避难所
[26 ] .
图3 青藏高原新生代古高程定量重建研究点(根据文献[<xref ref-type="bibr" rid="R25">25</xref>]修改) Quantitative reconstruction paleoelevation sites on Tibetan Plateau (Revised from Reference [<xref ref-type="bibr" rid="R25">25</xref>]) Fig.3 ![]()
图4 青藏高原南北剖面古高程变化示意图(根据文献[<xref ref-type="bibr" rid="R25">25</xref>]、[<xref ref-type="bibr" rid="R34">34</xref>]、[<xref ref-type="bibr" rid="R82">82</xref>]、[<xref ref-type="bibr" rid="R87">87</xref>]、[<xref ref-type="bibr" rid="R100">100</xref>]修改) North-south paleoelevation profiles on the Tibetan Plateau (Revised from References [25],[34],[82],[87],[100]) Fig.4 ![]()
随着DNA测序技术和比较系统学方法的发展,大量涌现的青藏高寒区生物地理学研究为我们理解高原隆升过程及其气候效应提供了生物演化方面的证据[25 ] .但是,演化生物学家通常仅对单一特定类群的系统发育过程开展研究,很少进行系统的总结工作[25 ] ,对某一类群遗传数据的解释也往往与最新的青藏高原隆升和气候变化研究进展脱节,不能很好地将最新的关于高原隆升和气候变化的研究与生物类群演化联系在一起[101 ] ,同时也没有认识到青藏高原的隆升过程是阶段性的,没有对青藏高原的不同组成部分进行区分[26 ,53 ,81 ] .地质学研究大多认为高原地貌格局形成于中新世之前,但是综合近年来的生物地理学研究结果可以发现,青藏高寒生物类群的分化时间多集中在晚中新世,种内分化加速和多样性增加主要发生在晚上新世到第四纪期间(图5 ),这些不同类群在分化时间上的巧合可能反映青藏高寒区生物演化受到晚中新世以来构造和气候事件的影响,但是并未引起地质学家足够的重视[25 ] . ...
... 注: 青藏高原隆升示意图从左到右依次为古近纪早期高原南部安第斯型山脉,古近纪中晚期高原中部东西向峡谷模型,新近纪早期高原中部和南部隆升,新近纪晚期高原北部生长[81 -82 ,84 -85 ,87 ,94 ,96 -97 ] ...
... Note: The tectonic history of the Tibetan Plateau shows the Andean-type mountains in the southern plateau in the early Paleogene, the east-west canyon model in the middle plateau until the late Paleogene, the uplift of the central and southern plateau in the early Neogene, and the growth of the northern plateau during the late Neogene[81 -82 ,84 -85 ,87 ,94 ,96 -97 ] ...
Biotic interchange through lowlands of Tibetan Plateau suture zones during Paleogene
2
2019
... 最近,青藏高原古地貌重建工作取得了丰硕的成果(图3 ),但是各种指标重建结果往往存在矛盾(图4 )[25 ,79 -83 ] .同位素重建结果表明高原南部在古近纪早期就存在安第斯型的巨大山系[84 -85 ] ;古生物研究发现高原腹地从中始新世到晚渐新世都存在东西向的大峡谷,该峡谷底部海拔在2 000 m左右,两侧为高大的冈底斯山脉和羌塘山脉[82 ,86 -88 ] ,动植物如巨犀(Paraceratherium )[89 ] 、攀鲈(Eoanabas )[90 -91 ] 、臭椿(Ailanthus )[88 ] 、椿榆(Cedrelospermum )[92 ] 、兔耳果(Lagokarpos )[93 ] 等在古近纪都可以通过青藏地区峡谷与周边动植物区系交流,新近纪之前的青藏地区虽然地形复杂,但是尚未隆升成今天的高原;横断山区某些区域则在晚始新世-早渐新世之交就已经获得现今的海拔高度[94 -95 ] ;高原北部和喜马拉雅山脉则在新近纪形成现在的地貌格局[25 ,96 -97 ] .古生物研究也发现青藏高原在晚上新世就已经存在高寒生境和冰缘环境,成为第四纪冰期动物忍耐寒冷环境的“训练营”,第四纪冰期来临之后这些适应寒冷环境的生物才扩散至北半球高纬度地区[34 ,98 -99 ] .因此,高原及周边地区高寒生物类群的起源时间表明在渐新世青藏高原部分区域就已经出现高寒生态系统,晚中新世以来青藏高原地貌与气候继续发生剧烈变化,促使这些物种的祖先类群迁入或就地演化积累适应高寒环境的基因和形态,并最终演化成适应高寒生境的新物种[25 ] .高寒区物种在晚上新世的加速分化可能与高原巨大山系和水系、冰川活动等造成的地理隔离有关[26 ] .随着第四纪冰期的到来,青藏高原及周边地区的高寒生物区也开始与北半球高纬度地区存在密切联系[34 ,98 -99 ] .同时,演化生物学研究也发现在第四纪冰期期间并未形成覆盖整个青藏高原的大冰盖,高原面上仍存在很多生物避难所[26 ] . ...
... [88 ]、椿榆(Cedrelospermum )[92 ] 、兔耳果(Lagokarpos )[93 ] 等在古近纪都可以通过青藏地区峡谷与周边动植物区系交流,新近纪之前的青藏地区虽然地形复杂,但是尚未隆升成今天的高原;横断山区某些区域则在晚始新世-早渐新世之交就已经获得现今的海拔高度[94 -95 ] ;高原北部和喜马拉雅山脉则在新近纪形成现在的地貌格局[25 ,96 -97 ] .古生物研究也发现青藏高原在晚上新世就已经存在高寒生境和冰缘环境,成为第四纪冰期动物忍耐寒冷环境的“训练营”,第四纪冰期来临之后这些适应寒冷环境的生物才扩散至北半球高纬度地区[34 ,98 -99 ] .因此,高原及周边地区高寒生物类群的起源时间表明在渐新世青藏高原部分区域就已经出现高寒生态系统,晚中新世以来青藏高原地貌与气候继续发生剧烈变化,促使这些物种的祖先类群迁入或就地演化积累适应高寒环境的基因和形态,并最终演化成适应高寒生境的新物种[25 ] .高寒区物种在晚上新世的加速分化可能与高原巨大山系和水系、冰川活动等造成的地理隔离有关[26 ] .随着第四纪冰期的到来,青藏高原及周边地区的高寒生物区也开始与北半球高纬度地区存在密切联系[34 ,98 -99 ] .同时,演化生物学研究也发现在第四纪冰期期间并未形成覆盖整个青藏高原的大冰盖,高原面上仍存在很多生物避难所[26 ] . ...
An Oligocene giant rhino provides insights into Paraceratherium evolution
1
2021
... 最近,青藏高原古地貌重建工作取得了丰硕的成果(图3 ),但是各种指标重建结果往往存在矛盾(图4 )[25 ,79 -83 ] .同位素重建结果表明高原南部在古近纪早期就存在安第斯型的巨大山系[84 -85 ] ;古生物研究发现高原腹地从中始新世到晚渐新世都存在东西向的大峡谷,该峡谷底部海拔在2 000 m左右,两侧为高大的冈底斯山脉和羌塘山脉[82 ,86 -88 ] ,动植物如巨犀(Paraceratherium )[89 ] 、攀鲈(Eoanabas )[90 -91 ] 、臭椿(Ailanthus )[88 ] 、椿榆(Cedrelospermum )[92 ] 、兔耳果(Lagokarpos )[93 ] 等在古近纪都可以通过青藏地区峡谷与周边动植物区系交流,新近纪之前的青藏地区虽然地形复杂,但是尚未隆升成今天的高原;横断山区某些区域则在晚始新世-早渐新世之交就已经获得现今的海拔高度[94 -95 ] ;高原北部和喜马拉雅山脉则在新近纪形成现在的地貌格局[25 ,96 -97 ] .古生物研究也发现青藏高原在晚上新世就已经存在高寒生境和冰缘环境,成为第四纪冰期动物忍耐寒冷环境的“训练营”,第四纪冰期来临之后这些适应寒冷环境的生物才扩散至北半球高纬度地区[34 ,98 -99 ] .因此,高原及周边地区高寒生物类群的起源时间表明在渐新世青藏高原部分区域就已经出现高寒生态系统,晚中新世以来青藏高原地貌与气候继续发生剧烈变化,促使这些物种的祖先类群迁入或就地演化积累适应高寒环境的基因和形态,并最终演化成适应高寒生境的新物种[25 ] .高寒区物种在晚上新世的加速分化可能与高原巨大山系和水系、冰川活动等造成的地理隔离有关[26 ] .随着第四纪冰期的到来,青藏高原及周边地区的高寒生物区也开始与北半球高纬度地区存在密切联系[34 ,98 -99 ] .同时,演化生物学研究也发现在第四纪冰期期间并未形成覆盖整个青藏高原的大冰盖,高原面上仍存在很多生物避难所[26 ] . ...
Into Africa via docked India: a fossil climbing perch from the Oligocene of Tibet helps solve the anabantid biogeographical puzzle
1
2019
... 最近,青藏高原古地貌重建工作取得了丰硕的成果(图3 ),但是各种指标重建结果往往存在矛盾(图4 )[25 ,79 -83 ] .同位素重建结果表明高原南部在古近纪早期就存在安第斯型的巨大山系[84 -85 ] ;古生物研究发现高原腹地从中始新世到晚渐新世都存在东西向的大峡谷,该峡谷底部海拔在2 000 m左右,两侧为高大的冈底斯山脉和羌塘山脉[82 ,86 -88 ] ,动植物如巨犀(Paraceratherium )[89 ] 、攀鲈(Eoanabas )[90 -91 ] 、臭椿(Ailanthus )[88 ] 、椿榆(Cedrelospermum )[92 ] 、兔耳果(Lagokarpos )[93 ] 等在古近纪都可以通过青藏地区峡谷与周边动植物区系交流,新近纪之前的青藏地区虽然地形复杂,但是尚未隆升成今天的高原;横断山区某些区域则在晚始新世-早渐新世之交就已经获得现今的海拔高度[94 -95 ] ;高原北部和喜马拉雅山脉则在新近纪形成现在的地貌格局[25 ,96 -97 ] .古生物研究也发现青藏高原在晚上新世就已经存在高寒生境和冰缘环境,成为第四纪冰期动物忍耐寒冷环境的“训练营”,第四纪冰期来临之后这些适应寒冷环境的生物才扩散至北半球高纬度地区[34 ,98 -99 ] .因此,高原及周边地区高寒生物类群的起源时间表明在渐新世青藏高原部分区域就已经出现高寒生态系统,晚中新世以来青藏高原地貌与气候继续发生剧烈变化,促使这些物种的祖先类群迁入或就地演化积累适应高寒环境的基因和形态,并最终演化成适应高寒生境的新物种[25 ] .高寒区物种在晚上新世的加速分化可能与高原巨大山系和水系、冰川活动等造成的地理隔离有关[26 ] .随着第四纪冰期的到来,青藏高原及周边地区的高寒生物区也开始与北半球高纬度地区存在密切联系[34 ,98 -99 ] .同时,演化生物学研究也发现在第四纪冰期期间并未形成覆盖整个青藏高原的大冰盖,高原面上仍存在很多生物避难所[26 ] . ...
Fossil climbing perch and associated plant megafossils indicate a warm and wet central Tibet during the late Oligocene
1
2017
... 最近,青藏高原古地貌重建工作取得了丰硕的成果(图3 ),但是各种指标重建结果往往存在矛盾(图4 )[25 ,79 -83 ] .同位素重建结果表明高原南部在古近纪早期就存在安第斯型的巨大山系[84 -85 ] ;古生物研究发现高原腹地从中始新世到晚渐新世都存在东西向的大峡谷,该峡谷底部海拔在2 000 m左右,两侧为高大的冈底斯山脉和羌塘山脉[82 ,86 -88 ] ,动植物如巨犀(Paraceratherium )[89 ] 、攀鲈(Eoanabas )[90 -91 ] 、臭椿(Ailanthus )[88 ] 、椿榆(Cedrelospermum )[92 ] 、兔耳果(Lagokarpos )[93 ] 等在古近纪都可以通过青藏地区峡谷与周边动植物区系交流,新近纪之前的青藏地区虽然地形复杂,但是尚未隆升成今天的高原;横断山区某些区域则在晚始新世-早渐新世之交就已经获得现今的海拔高度[94 -95 ] ;高原北部和喜马拉雅山脉则在新近纪形成现在的地貌格局[25 ,96 -97 ] .古生物研究也发现青藏高原在晚上新世就已经存在高寒生境和冰缘环境,成为第四纪冰期动物忍耐寒冷环境的“训练营”,第四纪冰期来临之后这些适应寒冷环境的生物才扩散至北半球高纬度地区[34 ,98 -99 ] .因此,高原及周边地区高寒生物类群的起源时间表明在渐新世青藏高原部分区域就已经出现高寒生态系统,晚中新世以来青藏高原地貌与气候继续发生剧烈变化,促使这些物种的祖先类群迁入或就地演化积累适应高寒环境的基因和形态,并最终演化成适应高寒生境的新物种[25 ] .高寒区物种在晚上新世的加速分化可能与高原巨大山系和水系、冰川活动等造成的地理隔离有关[26 ] .随着第四纪冰期的到来,青藏高原及周边地区的高寒生物区也开始与北半球高纬度地区存在密切联系[34 ,98 -99 ] .同时,演化生物学研究也发现在第四纪冰期期间并未形成覆盖整个青藏高原的大冰盖,高原面上仍存在很多生物避难所[26 ] . ...
First fossil record of Cedrelospermum (Ulmaceae) from the Qinghai‐Tibetan Plateau: implications for morphological evolution and biogeography
1
2019
... 最近,青藏高原古地貌重建工作取得了丰硕的成果(图3 ),但是各种指标重建结果往往存在矛盾(图4 )[25 ,79 -83 ] .同位素重建结果表明高原南部在古近纪早期就存在安第斯型的巨大山系[84 -85 ] ;古生物研究发现高原腹地从中始新世到晚渐新世都存在东西向的大峡谷,该峡谷底部海拔在2 000 m左右,两侧为高大的冈底斯山脉和羌塘山脉[82 ,86 -88 ] ,动植物如巨犀(Paraceratherium )[89 ] 、攀鲈(Eoanabas )[90 -91 ] 、臭椿(Ailanthus )[88 ] 、椿榆(Cedrelospermum )[92 ] 、兔耳果(Lagokarpos )[93 ] 等在古近纪都可以通过青藏地区峡谷与周边动植物区系交流,新近纪之前的青藏地区虽然地形复杂,但是尚未隆升成今天的高原;横断山区某些区域则在晚始新世-早渐新世之交就已经获得现今的海拔高度[94 -95 ] ;高原北部和喜马拉雅山脉则在新近纪形成现在的地貌格局[25 ,96 -97 ] .古生物研究也发现青藏高原在晚上新世就已经存在高寒生境和冰缘环境,成为第四纪冰期动物忍耐寒冷环境的“训练营”,第四纪冰期来临之后这些适应寒冷环境的生物才扩散至北半球高纬度地区[34 ,98 -99 ] .因此,高原及周边地区高寒生物类群的起源时间表明在渐新世青藏高原部分区域就已经出现高寒生态系统,晚中新世以来青藏高原地貌与气候继续发生剧烈变化,促使这些物种的祖先类群迁入或就地演化积累适应高寒环境的基因和形态,并最终演化成适应高寒生境的新物种[25 ] .高寒区物种在晚上新世的加速分化可能与高原巨大山系和水系、冰川活动等造成的地理隔离有关[26 ] .随着第四纪冰期的到来,青藏高原及周边地区的高寒生物区也开始与北半球高纬度地区存在密切联系[34 ,98 -99 ] .同时,演化生物学研究也发现在第四纪冰期期间并未形成覆盖整个青藏高原的大冰盖,高原面上仍存在很多生物避难所[26 ] . ...
Extinct genus Lagokarpos reveals a biogeographic connection between Tibet and other regions in the Northern Hemisphere during the Paleogene
1
2019
... 最近,青藏高原古地貌重建工作取得了丰硕的成果(图3 ),但是各种指标重建结果往往存在矛盾(图4 )[25 ,79 -83 ] .同位素重建结果表明高原南部在古近纪早期就存在安第斯型的巨大山系[84 -85 ] ;古生物研究发现高原腹地从中始新世到晚渐新世都存在东西向的大峡谷,该峡谷底部海拔在2 000 m左右,两侧为高大的冈底斯山脉和羌塘山脉[82 ,86 -88 ] ,动植物如巨犀(Paraceratherium )[89 ] 、攀鲈(Eoanabas )[90 -91 ] 、臭椿(Ailanthus )[88 ] 、椿榆(Cedrelospermum )[92 ] 、兔耳果(Lagokarpos )[93 ] 等在古近纪都可以通过青藏地区峡谷与周边动植物区系交流,新近纪之前的青藏地区虽然地形复杂,但是尚未隆升成今天的高原;横断山区某些区域则在晚始新世-早渐新世之交就已经获得现今的海拔高度[94 -95 ] ;高原北部和喜马拉雅山脉则在新近纪形成现在的地貌格局[25 ,96 -97 ] .古生物研究也发现青藏高原在晚上新世就已经存在高寒生境和冰缘环境,成为第四纪冰期动物忍耐寒冷环境的“训练营”,第四纪冰期来临之后这些适应寒冷环境的生物才扩散至北半球高纬度地区[34 ,98 -99 ] .因此,高原及周边地区高寒生物类群的起源时间表明在渐新世青藏高原部分区域就已经出现高寒生态系统,晚中新世以来青藏高原地貌与气候继续发生剧烈变化,促使这些物种的祖先类群迁入或就地演化积累适应高寒环境的基因和形态,并最终演化成适应高寒生境的新物种[25 ] .高寒区物种在晚上新世的加速分化可能与高原巨大山系和水系、冰川活动等造成的地理隔离有关[26 ] .随着第四纪冰期的到来,青藏高原及周边地区的高寒生物区也开始与北半球高纬度地区存在密切联系[34 ,98 -99 ] .同时,演化生物学研究也发现在第四纪冰期期间并未形成覆盖整个青藏高原的大冰盖,高原面上仍存在很多生物避难所[26 ] . ...
Uplift, climate and biotic changes at the eocene-oligocene transition in Southeast Tibet
4
2019
... 最近,青藏高原古地貌重建工作取得了丰硕的成果(图3 ),但是各种指标重建结果往往存在矛盾(图4 )[25 ,79 -83 ] .同位素重建结果表明高原南部在古近纪早期就存在安第斯型的巨大山系[84 -85 ] ;古生物研究发现高原腹地从中始新世到晚渐新世都存在东西向的大峡谷,该峡谷底部海拔在2 000 m左右,两侧为高大的冈底斯山脉和羌塘山脉[82 ,86 -88 ] ,动植物如巨犀(Paraceratherium )[89 ] 、攀鲈(Eoanabas )[90 -91 ] 、臭椿(Ailanthus )[88 ] 、椿榆(Cedrelospermum )[92 ] 、兔耳果(Lagokarpos )[93 ] 等在古近纪都可以通过青藏地区峡谷与周边动植物区系交流,新近纪之前的青藏地区虽然地形复杂,但是尚未隆升成今天的高原;横断山区某些区域则在晚始新世-早渐新世之交就已经获得现今的海拔高度[94 -95 ] ;高原北部和喜马拉雅山脉则在新近纪形成现在的地貌格局[25 ,96 -97 ] .古生物研究也发现青藏高原在晚上新世就已经存在高寒生境和冰缘环境,成为第四纪冰期动物忍耐寒冷环境的“训练营”,第四纪冰期来临之后这些适应寒冷环境的生物才扩散至北半球高纬度地区[34 ,98 -99 ] .因此,高原及周边地区高寒生物类群的起源时间表明在渐新世青藏高原部分区域就已经出现高寒生态系统,晚中新世以来青藏高原地貌与气候继续发生剧烈变化,促使这些物种的祖先类群迁入或就地演化积累适应高寒环境的基因和形态,并最终演化成适应高寒生境的新物种[25 ] .高寒区物种在晚上新世的加速分化可能与高原巨大山系和水系、冰川活动等造成的地理隔离有关[26 ] .随着第四纪冰期的到来,青藏高原及周边地区的高寒生物区也开始与北半球高纬度地区存在密切联系[34 ,98 -99 ] .同时,演化生物学研究也发现在第四纪冰期期间并未形成覆盖整个青藏高原的大冰盖,高原面上仍存在很多生物避难所[26 ] . ...
... 注: 青藏高原隆升示意图从左到右依次为古近纪早期高原南部安第斯型山脉,古近纪中晚期高原中部东西向峡谷模型,新近纪早期高原中部和南部隆升,新近纪晚期高原北部生长[81 -82 ,84 -85 ,87 ,94 ,96 -97 ] ...
... Note: The tectonic history of the Tibetan Plateau shows the Andean-type mountains in the southern plateau in the early Paleogene, the east-west canyon model in the middle plateau until the late Paleogene, the uplift of the central and southern plateau in the early Neogene, and the growth of the northern plateau during the late Neogene[81 -82 ,84 -85 ,87 ,94 ,96 -97 ] ...
... 近期,演化生物学家与地质学家对青藏高原及周边高寒生物区18个被子植物类群共3 798个植物物种的遗传数据进行整合研究,发现横断山区是世界上起源最早的高寒生物区,该区域高寒植物多样性在早渐新世就已经开始积累,并分别在晚渐新世和中中新世就地演化速率快速增加(图4 )[26 ] .高寒植物在早渐新世就出现在横断山区与该区域在晚始新世-早渐新世之交就已经出现与现今海拔相似的高山环境有关[94 -95 ] .中新世就地演化速率快速增加与季风气候增强,降水增加,河流切割加剧形成生物地理隔离有关[26 ,104 ] .横断山区高寒生物演化进程与喜马拉雅山脉和青藏高原腹地的对比研究发现,横断山区是青藏高原高寒生物区物种起源和分化的摇篮,也是高寒生物的重要避难所,为青藏高原腹地和喜马拉雅山脉高寒生物多样性提供主要来源[26 ] .第四纪以来,冰期-间冰期气候变化剧烈,横断山区近南北向的高山狭谷地貌格局为遭受极端气候的北方类群和高寒生物类群的迁徙提供廊道,此外,深切狭谷与高山也可以保障山地动植物类群短时间、短距离地垂直迁徙;与此同时,横断山区高山狭谷地貌导致的地理和生境的巨大差异,提供了物种隔离障碍,成为许多动植物类群的新种分化地[26 ] .冰期时,全球变冷,冰川活动加剧,高寒生物区面积扩张,临近山地的高寒生物区之间的交流也大大增加[26 ] .该研究为高原隆升、气候变化和生物演化提供一个多学科交叉研究的范例. ...
The early Eocene rise of the Gonjo Basin, SE Tibet: from low desert to high forest
2
2020
... 最近,青藏高原古地貌重建工作取得了丰硕的成果(图3 ),但是各种指标重建结果往往存在矛盾(图4 )[25 ,79 -83 ] .同位素重建结果表明高原南部在古近纪早期就存在安第斯型的巨大山系[84 -85 ] ;古生物研究发现高原腹地从中始新世到晚渐新世都存在东西向的大峡谷,该峡谷底部海拔在2 000 m左右,两侧为高大的冈底斯山脉和羌塘山脉[82 ,86 -88 ] ,动植物如巨犀(Paraceratherium )[89 ] 、攀鲈(Eoanabas )[90 -91 ] 、臭椿(Ailanthus )[88 ] 、椿榆(Cedrelospermum )[92 ] 、兔耳果(Lagokarpos )[93 ] 等在古近纪都可以通过青藏地区峡谷与周边动植物区系交流,新近纪之前的青藏地区虽然地形复杂,但是尚未隆升成今天的高原;横断山区某些区域则在晚始新世-早渐新世之交就已经获得现今的海拔高度[94 -95 ] ;高原北部和喜马拉雅山脉则在新近纪形成现在的地貌格局[25 ,96 -97 ] .古生物研究也发现青藏高原在晚上新世就已经存在高寒生境和冰缘环境,成为第四纪冰期动物忍耐寒冷环境的“训练营”,第四纪冰期来临之后这些适应寒冷环境的生物才扩散至北半球高纬度地区[34 ,98 -99 ] .因此,高原及周边地区高寒生物类群的起源时间表明在渐新世青藏高原部分区域就已经出现高寒生态系统,晚中新世以来青藏高原地貌与气候继续发生剧烈变化,促使这些物种的祖先类群迁入或就地演化积累适应高寒环境的基因和形态,并最终演化成适应高寒生境的新物种[25 ] .高寒区物种在晚上新世的加速分化可能与高原巨大山系和水系、冰川活动等造成的地理隔离有关[26 ] .随着第四纪冰期的到来,青藏高原及周边地区的高寒生物区也开始与北半球高纬度地区存在密切联系[34 ,98 -99 ] .同时,演化生物学研究也发现在第四纪冰期期间并未形成覆盖整个青藏高原的大冰盖,高原面上仍存在很多生物避难所[26 ] . ...
... 近期,演化生物学家与地质学家对青藏高原及周边高寒生物区18个被子植物类群共3 798个植物物种的遗传数据进行整合研究,发现横断山区是世界上起源最早的高寒生物区,该区域高寒植物多样性在早渐新世就已经开始积累,并分别在晚渐新世和中中新世就地演化速率快速增加(图4 )[26 ] .高寒植物在早渐新世就出现在横断山区与该区域在晚始新世-早渐新世之交就已经出现与现今海拔相似的高山环境有关[94 -95 ] .中新世就地演化速率快速增加与季风气候增强,降水增加,河流切割加剧形成生物地理隔离有关[26 ,104 ] .横断山区高寒生物演化进程与喜马拉雅山脉和青藏高原腹地的对比研究发现,横断山区是青藏高原高寒生物区物种起源和分化的摇篮,也是高寒生物的重要避难所,为青藏高原腹地和喜马拉雅山脉高寒生物多样性提供主要来源[26 ] .第四纪以来,冰期-间冰期气候变化剧烈,横断山区近南北向的高山狭谷地貌格局为遭受极端气候的北方类群和高寒生物类群的迁徙提供廊道,此外,深切狭谷与高山也可以保障山地动植物类群短时间、短距离地垂直迁徙;与此同时,横断山区高山狭谷地貌导致的地理和生境的巨大差异,提供了物种隔离障碍,成为许多动植物类群的新种分化地[26 ] .冰期时,全球变冷,冰川活动加剧,高寒生物区面积扩张,临近山地的高寒生物区之间的交流也大大增加[26 ] .该研究为高原隆升、气候变化和生物演化提供一个多学科交叉研究的范例. ...
Late Miocene-Quaternary rapid stepwise uplift of the NE Tibetan Plateau and its effects on climatic and environmental changes
3
2014
... 最近,青藏高原古地貌重建工作取得了丰硕的成果(图3 ),但是各种指标重建结果往往存在矛盾(图4 )[25 ,79 -83 ] .同位素重建结果表明高原南部在古近纪早期就存在安第斯型的巨大山系[84 -85 ] ;古生物研究发现高原腹地从中始新世到晚渐新世都存在东西向的大峡谷,该峡谷底部海拔在2 000 m左右,两侧为高大的冈底斯山脉和羌塘山脉[82 ,86 -88 ] ,动植物如巨犀(Paraceratherium )[89 ] 、攀鲈(Eoanabas )[90 -91 ] 、臭椿(Ailanthus )[88 ] 、椿榆(Cedrelospermum )[92 ] 、兔耳果(Lagokarpos )[93 ] 等在古近纪都可以通过青藏地区峡谷与周边动植物区系交流,新近纪之前的青藏地区虽然地形复杂,但是尚未隆升成今天的高原;横断山区某些区域则在晚始新世-早渐新世之交就已经获得现今的海拔高度[94 -95 ] ;高原北部和喜马拉雅山脉则在新近纪形成现在的地貌格局[25 ,96 -97 ] .古生物研究也发现青藏高原在晚上新世就已经存在高寒生境和冰缘环境,成为第四纪冰期动物忍耐寒冷环境的“训练营”,第四纪冰期来临之后这些适应寒冷环境的生物才扩散至北半球高纬度地区[34 ,98 -99 ] .因此,高原及周边地区高寒生物类群的起源时间表明在渐新世青藏高原部分区域就已经出现高寒生态系统,晚中新世以来青藏高原地貌与气候继续发生剧烈变化,促使这些物种的祖先类群迁入或就地演化积累适应高寒环境的基因和形态,并最终演化成适应高寒生境的新物种[25 ] .高寒区物种在晚上新世的加速分化可能与高原巨大山系和水系、冰川活动等造成的地理隔离有关[26 ] .随着第四纪冰期的到来,青藏高原及周边地区的高寒生物区也开始与北半球高纬度地区存在密切联系[34 ,98 -99 ] .同时,演化生物学研究也发现在第四纪冰期期间并未形成覆盖整个青藏高原的大冰盖,高原面上仍存在很多生物避难所[26 ] . ...
... 注: 青藏高原隆升示意图从左到右依次为古近纪早期高原南部安第斯型山脉,古近纪中晚期高原中部东西向峡谷模型,新近纪早期高原中部和南部隆升,新近纪晚期高原北部生长[81 -82 ,84 -85 ,87 ,94 ,96 -97 ] ...
... Note: The tectonic history of the Tibetan Plateau shows the Andean-type mountains in the southern plateau in the early Paleogene, the east-west canyon model in the middle plateau until the late Paleogene, the uplift of the central and southern plateau in the early Neogene, and the growth of the northern plateau during the late Neogene[81 -82 ,84 -85 ,87 ,94 ,96 -97 ] ...
Constant elevation of southern Tibet over the past 15 million years
3
2003
... 最近,青藏高原古地貌重建工作取得了丰硕的成果(图3 ),但是各种指标重建结果往往存在矛盾(图4 )[25 ,79 -83 ] .同位素重建结果表明高原南部在古近纪早期就存在安第斯型的巨大山系[84 -85 ] ;古生物研究发现高原腹地从中始新世到晚渐新世都存在东西向的大峡谷,该峡谷底部海拔在2 000 m左右,两侧为高大的冈底斯山脉和羌塘山脉[82 ,86 -88 ] ,动植物如巨犀(Paraceratherium )[89 ] 、攀鲈(Eoanabas )[90 -91 ] 、臭椿(Ailanthus )[88 ] 、椿榆(Cedrelospermum )[92 ] 、兔耳果(Lagokarpos )[93 ] 等在古近纪都可以通过青藏地区峡谷与周边动植物区系交流,新近纪之前的青藏地区虽然地形复杂,但是尚未隆升成今天的高原;横断山区某些区域则在晚始新世-早渐新世之交就已经获得现今的海拔高度[94 -95 ] ;高原北部和喜马拉雅山脉则在新近纪形成现在的地貌格局[25 ,96 -97 ] .古生物研究也发现青藏高原在晚上新世就已经存在高寒生境和冰缘环境,成为第四纪冰期动物忍耐寒冷环境的“训练营”,第四纪冰期来临之后这些适应寒冷环境的生物才扩散至北半球高纬度地区[34 ,98 -99 ] .因此,高原及周边地区高寒生物类群的起源时间表明在渐新世青藏高原部分区域就已经出现高寒生态系统,晚中新世以来青藏高原地貌与气候继续发生剧烈变化,促使这些物种的祖先类群迁入或就地演化积累适应高寒环境的基因和形态,并最终演化成适应高寒生境的新物种[25 ] .高寒区物种在晚上新世的加速分化可能与高原巨大山系和水系、冰川活动等造成的地理隔离有关[26 ] .随着第四纪冰期的到来,青藏高原及周边地区的高寒生物区也开始与北半球高纬度地区存在密切联系[34 ,98 -99 ] .同时,演化生物学研究也发现在第四纪冰期期间并未形成覆盖整个青藏高原的大冰盖,高原面上仍存在很多生物避难所[26 ] . ...
... 注: 青藏高原隆升示意图从左到右依次为古近纪早期高原南部安第斯型山脉,古近纪中晚期高原中部东西向峡谷模型,新近纪早期高原中部和南部隆升,新近纪晚期高原北部生长[81 -82 ,84 -85 ,87 ,94 ,96 -97 ] ...
... Note: The tectonic history of the Tibetan Plateau shows the Andean-type mountains in the southern plateau in the early Paleogene, the east-west canyon model in the middle plateau until the late Paleogene, the uplift of the central and southern plateau in the early Neogene, and the growth of the northern plateau during the late Neogene[81 -82 ,84 -85 ,87 ,94 ,96 -97 ] ...
Out of Tibet: Pliocene woolly rhino suggests high-plateau origin of Ice Age megaherbivores
2
2011
... 最近,青藏高原古地貌重建工作取得了丰硕的成果(图3 ),但是各种指标重建结果往往存在矛盾(图4 )[25 ,79 -83 ] .同位素重建结果表明高原南部在古近纪早期就存在安第斯型的巨大山系[84 -85 ] ;古生物研究发现高原腹地从中始新世到晚渐新世都存在东西向的大峡谷,该峡谷底部海拔在2 000 m左右,两侧为高大的冈底斯山脉和羌塘山脉[82 ,86 -88 ] ,动植物如巨犀(Paraceratherium )[89 ] 、攀鲈(Eoanabas )[90 -91 ] 、臭椿(Ailanthus )[88 ] 、椿榆(Cedrelospermum )[92 ] 、兔耳果(Lagokarpos )[93 ] 等在古近纪都可以通过青藏地区峡谷与周边动植物区系交流,新近纪之前的青藏地区虽然地形复杂,但是尚未隆升成今天的高原;横断山区某些区域则在晚始新世-早渐新世之交就已经获得现今的海拔高度[94 -95 ] ;高原北部和喜马拉雅山脉则在新近纪形成现在的地貌格局[25 ,96 -97 ] .古生物研究也发现青藏高原在晚上新世就已经存在高寒生境和冰缘环境,成为第四纪冰期动物忍耐寒冷环境的“训练营”,第四纪冰期来临之后这些适应寒冷环境的生物才扩散至北半球高纬度地区[34 ,98 -99 ] .因此,高原及周边地区高寒生物类群的起源时间表明在渐新世青藏高原部分区域就已经出现高寒生态系统,晚中新世以来青藏高原地貌与气候继续发生剧烈变化,促使这些物种的祖先类群迁入或就地演化积累适应高寒环境的基因和形态,并最终演化成适应高寒生境的新物种[25 ] .高寒区物种在晚上新世的加速分化可能与高原巨大山系和水系、冰川活动等造成的地理隔离有关[26 ] .随着第四纪冰期的到来,青藏高原及周边地区的高寒生物区也开始与北半球高纬度地区存在密切联系[34 ,98 -99 ] .同时,演化生物学研究也发现在第四纪冰期期间并未形成覆盖整个青藏高原的大冰盖,高原面上仍存在很多生物避难所[26 ] . ...
... ,98 -99 ].同时,演化生物学研究也发现在第四纪冰期期间并未形成覆盖整个青藏高原的大冰盖,高原面上仍存在很多生物避难所[26 ] . ...
From ‘third pole’ to north pole: a Himalayan origin for the arctic fox
2
2014
... 最近,青藏高原古地貌重建工作取得了丰硕的成果(图3 ),但是各种指标重建结果往往存在矛盾(图4 )[25 ,79 -83 ] .同位素重建结果表明高原南部在古近纪早期就存在安第斯型的巨大山系[84 -85 ] ;古生物研究发现高原腹地从中始新世到晚渐新世都存在东西向的大峡谷,该峡谷底部海拔在2 000 m左右,两侧为高大的冈底斯山脉和羌塘山脉[82 ,86 -88 ] ,动植物如巨犀(Paraceratherium )[89 ] 、攀鲈(Eoanabas )[90 -91 ] 、臭椿(Ailanthus )[88 ] 、椿榆(Cedrelospermum )[92 ] 、兔耳果(Lagokarpos )[93 ] 等在古近纪都可以通过青藏地区峡谷与周边动植物区系交流,新近纪之前的青藏地区虽然地形复杂,但是尚未隆升成今天的高原;横断山区某些区域则在晚始新世-早渐新世之交就已经获得现今的海拔高度[94 -95 ] ;高原北部和喜马拉雅山脉则在新近纪形成现在的地貌格局[25 ,96 -97 ] .古生物研究也发现青藏高原在晚上新世就已经存在高寒生境和冰缘环境,成为第四纪冰期动物忍耐寒冷环境的“训练营”,第四纪冰期来临之后这些适应寒冷环境的生物才扩散至北半球高纬度地区[34 ,98 -99 ] .因此,高原及周边地区高寒生物类群的起源时间表明在渐新世青藏高原部分区域就已经出现高寒生态系统,晚中新世以来青藏高原地貌与气候继续发生剧烈变化,促使这些物种的祖先类群迁入或就地演化积累适应高寒环境的基因和形态,并最终演化成适应高寒生境的新物种[25 ] .高寒区物种在晚上新世的加速分化可能与高原巨大山系和水系、冰川活动等造成的地理隔离有关[26 ] .随着第四纪冰期的到来,青藏高原及周边地区的高寒生物区也开始与北半球高纬度地区存在密切联系[34 ,98 -99 ] .同时,演化生物学研究也发现在第四纪冰期期间并未形成覆盖整个青藏高原的大冰盖,高原面上仍存在很多生物避难所[26 ] . ...
... -99 ].同时,演化生物学研究也发现在第四纪冰期期间并未形成覆盖整个青藏高原的大冰盖,高原面上仍存在很多生物避难所[26 ] . ...
Paleoaltimetry reconstructions of the Tibetan Plateau: progress and contradictions
1
2015
... 最近,青藏高原古地貌重建工作取得了丰硕的成果(
图3 ),但是各种指标重建结果往往存在矛盾(
图4 )
[25 ,79 -83 ] .同位素重建结果表明高原南部在古近纪早期就存在安第斯型的巨大山系
[84 -85 ] ;古生物研究发现高原腹地从中始新世到晚渐新世都存在东西向的大峡谷,该峡谷底部海拔在2 000 m左右,两侧为高大的冈底斯山脉和羌塘山脉
[82 ,86 -88 ] ,动植物如巨犀(
Paraceratherium )
[89 ] 、攀鲈(
Eoanabas )
[90 -91 ] 、臭椿(
Ailanthus )
[88 ] 、椿榆(
Cedrelospermum )
[92 ] 、兔耳果(
Lagokarpos )
[93 ] 等在古近纪都可以通过青藏地区峡谷与周边动植物区系交流,新近纪之前的青藏地区虽然地形复杂,但是尚未隆升成今天的高原;横断山区某些区域则在晚始新世-早渐新世之交就已经获得现今的海拔高度
[94 -95 ] ;高原北部和喜马拉雅山脉则在新近纪形成现在的地貌格局
[25 ,96 -97 ] .古生物研究也发现青藏高原在晚上新世就已经存在高寒生境和冰缘环境,成为第四纪冰期动物忍耐寒冷环境的“训练营”,第四纪冰期来临之后这些适应寒冷环境的生物才扩散至北半球高纬度地区
[34 ,98 -99 ] .因此,高原及周边地区高寒生物类群的起源时间表明在渐新世青藏高原部分区域就已经出现高寒生态系统,晚中新世以来青藏高原地貌与气候继续发生剧烈变化,促使这些物种的祖先类群迁入或就地演化积累适应高寒环境的基因和形态,并最终演化成适应高寒生境的新物种
[25 ] .高寒区物种在晚上新世的加速分化可能与高原巨大山系和水系、冰川活动等造成的地理隔离有关
[26 ] .随着第四纪冰期的到来,青藏高原及周边地区的高寒生物区也开始与北半球高纬度地区存在密切联系
[34 ,98 -99 ] .同时,演化生物学研究也发现在第四纪冰期期间并未形成覆盖整个青藏高原的大冰盖,高原面上仍存在很多生物避难所
[26 ] .
图3 青藏高原新生代古高程定量重建研究点(根据文献[<xref ref-type="bibr" rid="R25">25</xref>]修改) Quantitative reconstruction paleoelevation sites on Tibetan Plateau (Revised from Reference [<xref ref-type="bibr" rid="R25">25</xref>]) Fig.3 ![]()
图4 青藏高原南北剖面古高程变化示意图(根据文献[<xref ref-type="bibr" rid="R25">25</xref>]、[<xref ref-type="bibr" rid="R34">34</xref>]、[<xref ref-type="bibr" rid="R82">82</xref>]、[<xref ref-type="bibr" rid="R87">87</xref>]、[<xref ref-type="bibr" rid="R100">100</xref>]修改) North-south paleoelevation profiles on the Tibetan Plateau (Revised from References [25],[34],[82],[87],[100]) Fig.4 ![]()
随着DNA测序技术和比较系统学方法的发展,大量涌现的青藏高寒区生物地理学研究为我们理解高原隆升过程及其气候效应提供了生物演化方面的证据[25 ] .但是,演化生物学家通常仅对单一特定类群的系统发育过程开展研究,很少进行系统的总结工作[25 ] ,对某一类群遗传数据的解释也往往与最新的青藏高原隆升和气候变化研究进展脱节,不能很好地将最新的关于高原隆升和气候变化的研究与生物类群演化联系在一起[101 ] ,同时也没有认识到青藏高原的隆升过程是阶段性的,没有对青藏高原的不同组成部分进行区分[26 ,53 ,81 ] .地质学研究大多认为高原地貌格局形成于中新世之前,但是综合近年来的生物地理学研究结果可以发现,青藏高寒生物类群的分化时间多集中在晚中新世,种内分化加速和多样性增加主要发生在晚上新世到第四纪期间(图5 ),这些不同类群在分化时间上的巧合可能反映青藏高寒区生物演化受到晚中新世以来构造和气候事件的影响,但是并未引起地质学家足够的重视[25 ] . ...
Available data point to a 4-km-high Tibetan Plateau by 40 Ma, but 100 molecular-clock papers have linked supposed recent uplift to young node ages
1
2016
... 随着DNA测序技术和比较系统学方法的发展,大量涌现的青藏高寒区生物地理学研究为我们理解高原隆升过程及其气候效应提供了生物演化方面的证据[25 ] .但是,演化生物学家通常仅对单一特定类群的系统发育过程开展研究,很少进行系统的总结工作[25 ] ,对某一类群遗传数据的解释也往往与最新的青藏高原隆升和气候变化研究进展脱节,不能很好地将最新的关于高原隆升和气候变化的研究与生物类群演化联系在一起[101 ] ,同时也没有认识到青藏高原的隆升过程是阶段性的,没有对青藏高原的不同组成部分进行区分[26 ,53 ,81 ] .地质学研究大多认为高原地貌格局形成于中新世之前,但是综合近年来的生物地理学研究结果可以发现,青藏高寒生物类群的分化时间多集中在晚中新世,种内分化加速和多样性增加主要发生在晚上新世到第四纪期间(图5 ),这些不同类群在分化时间上的巧合可能反映青藏高寒区生物演化受到晚中新世以来构造和气候事件的影响,但是并未引起地质学家足够的重视[25 ] . ...
Cenozoic climate changes: a review based on time series analysis of marine benthic δ 18 O records
1
2014
... 随着DNA测序技术和比较系统学方法的发展,大量涌现的青藏高寒区生物地理学研究为我们理解高原隆升过程及其气候效应提供了生物演化方面的证据
[25 ] .但是,演化生物学家通常仅对单一特定类群的系统发育过程开展研究,很少进行系统的总结工作
[25 ] ,对某一类群遗传数据的解释也往往与最新的青藏高原隆升和气候变化研究进展脱节,不能很好地将最新的关于高原隆升和气候变化的研究与生物类群演化联系在一起
[101 ] ,同时也没有认识到青藏高原的隆升过程是阶段性的,没有对青藏高原的不同组成部分进行区分
[26 ,53 ,81 ] .地质学研究大多认为高原地貌格局形成于中新世之前,但是综合近年来的生物地理学研究结果可以发现,青藏高寒生物类群的分化时间多集中在晚中新世,种内分化加速和多样性增加主要发生在晚上新世到第四纪期间(
图5 ),这些不同类群在分化时间上的巧合可能反映青藏高寒区生物演化受到晚中新世以来构造和气候事件的影响,但是并未引起地质学家足够的重视
[25 ] .
图5 青藏高寒区生物分化时间、新生代全球气候变化和高原隆升关系(新生代气候变化根据文献[<xref ref-type="bibr" rid="R102">102</xref>]、[103]修改) 注: 青藏高原隆升示意图从左到右依次为古近纪早期高原南部安第斯型山脉,古近纪中晚期高原中部东西向峡谷模型,新近纪早期高原中部和南部隆升,新近纪晚期高原北部生长[81 -82 ,84 -85 ,87 ,94 ,96 -97 ] ...
Trends, rhythms, and aberrations in global climate 65 Ma to present
0
2001
Rapid incision of the Mekong River in the middle Miocene linked to monsoonal precipitation
1
2018
... 近期,演化生物学家与地质学家对青藏高原及周边高寒生物区18个被子植物类群共3 798个植物物种的遗传数据进行整合研究,发现横断山区是世界上起源最早的高寒生物区,该区域高寒植物多样性在早渐新世就已经开始积累,并分别在晚渐新世和中中新世就地演化速率快速增加(图4 )[26 ] .高寒植物在早渐新世就出现在横断山区与该区域在晚始新世-早渐新世之交就已经出现与现今海拔相似的高山环境有关[94 -95 ] .中新世就地演化速率快速增加与季风气候增强,降水增加,河流切割加剧形成生物地理隔离有关[26 ,104 ] .横断山区高寒生物演化进程与喜马拉雅山脉和青藏高原腹地的对比研究发现,横断山区是青藏高原高寒生物区物种起源和分化的摇篮,也是高寒生物的重要避难所,为青藏高原腹地和喜马拉雅山脉高寒生物多样性提供主要来源[26 ] .第四纪以来,冰期-间冰期气候变化剧烈,横断山区近南北向的高山狭谷地貌格局为遭受极端气候的北方类群和高寒生物类群的迁徙提供廊道,此外,深切狭谷与高山也可以保障山地动植物类群短时间、短距离地垂直迁徙;与此同时,横断山区高山狭谷地貌导致的地理和生境的巨大差异,提供了物种隔离障碍,成为许多动植物类群的新种分化地[26 ] .冰期时,全球变冷,冰川活动加剧,高寒生物区面积扩张,临近山地的高寒生物区之间的交流也大大增加[26 ] .该研究为高原隆升、气候变化和生物演化提供一个多学科交叉研究的范例. ...


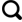


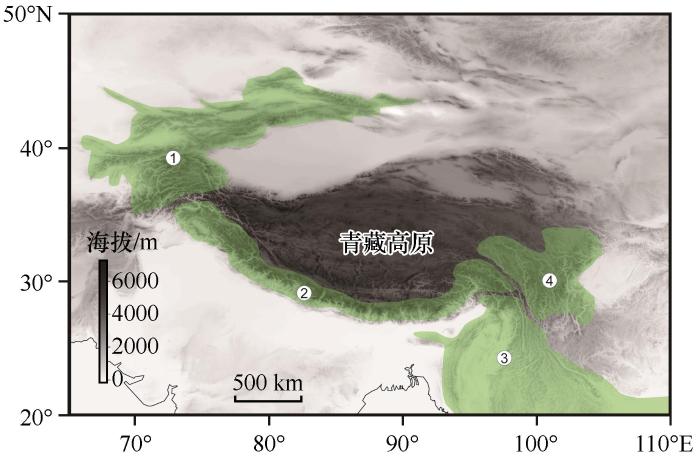
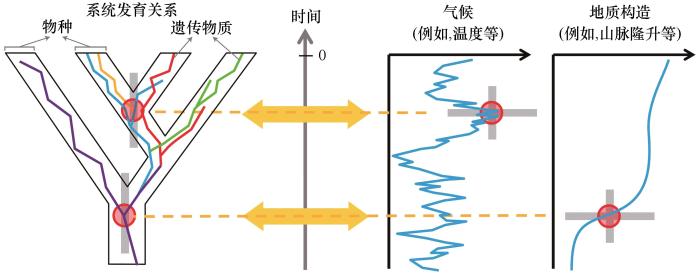
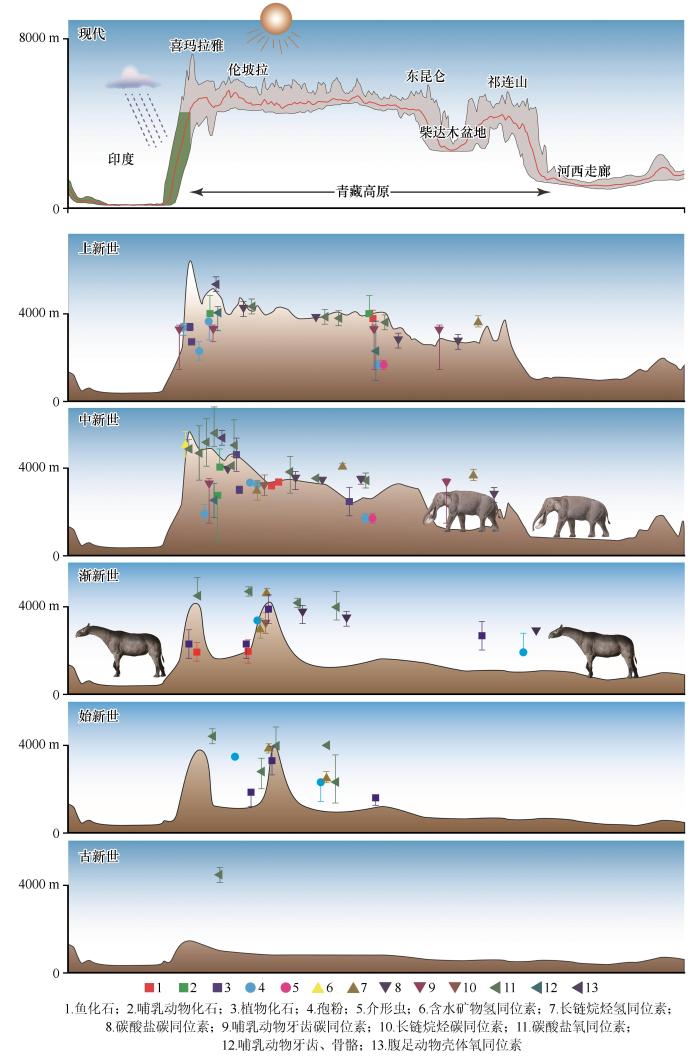
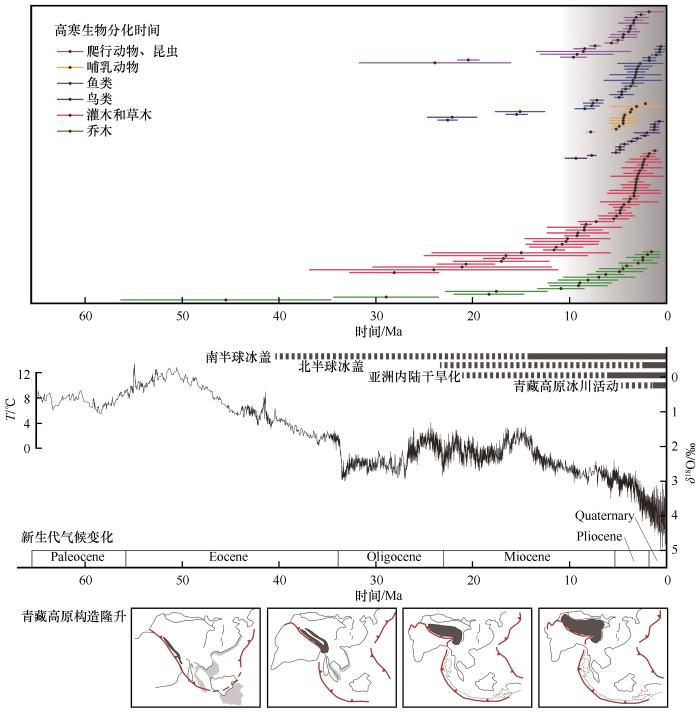

 甘公网安备 62010202000676号
甘公网安备 62010202000676号
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}