Trends, rhythms, and aberrations in global climate 65 Ma to present
2
2001
... 新生代以来,全球气候至少经历了始新世-渐新世之交、中中新世和上新世末期的三次台阶式降温[1 -2 ] .上新世-更新世转型是上新世的温暖气候向更新世冰期-间冰期气候转换的重要时段,同时也是现今地质-地貌格局、气候、生态系统和极地冰盖最终稳定形成的重要演化期[1 ,3 -8 ] .晚上新世末北半球冰盖快速扩张的驱动机制一直是讨论的焦点:有研究认为巴拿马海峡的关闭或晚上新世以来大气CO2 浓度减少控制着北极冰盖发育,更有学者提出构造隆升对北极冰盖演化和全球变冷起到驱动作用,指出青藏高原隆升对全球气候变化的重要性[3 ,8 ] .青藏高原强烈隆升使得高原及周边地区沉积巨厚砾岩[8 -9 ] ,内陆干旱化加剧,在我国黄土高原地区堆积典型的第四纪风成黄土-古土壤序列[10 -11 ] . ...
... [1 ,3 -8 ].晚上新世末北半球冰盖快速扩张的驱动机制一直是讨论的焦点:有研究认为巴拿马海峡的关闭或晚上新世以来大气CO2 浓度减少控制着北极冰盖发育,更有学者提出构造隆升对北极冰盖演化和全球变冷起到驱动作用,指出青藏高原隆升对全球气候变化的重要性[3 ,8 ] .青藏高原强烈隆升使得高原及周边地区沉积巨厚砾岩[8 -9 ] ,内陆干旱化加剧,在我国黄土高原地区堆积典型的第四纪风成黄土-古土壤序列[10 -11 ] . ...
Cenozoic climate changes: a review based on time series analysis of marine benthic δ 18 O records
1
2014
... 新生代以来,全球气候至少经历了始新世-渐新世之交、中中新世和上新世末期的三次台阶式降温[1 -2 ] .上新世-更新世转型是上新世的温暖气候向更新世冰期-间冰期气候转换的重要时段,同时也是现今地质-地貌格局、气候、生态系统和极地冰盖最终稳定形成的重要演化期[1 ,3 -8 ] .晚上新世末北半球冰盖快速扩张的驱动机制一直是讨论的焦点:有研究认为巴拿马海峡的关闭或晚上新世以来大气CO2 浓度减少控制着北极冰盖发育,更有学者提出构造隆升对北极冰盖演化和全球变冷起到驱动作用,指出青藏高原隆升对全球气候变化的重要性[3 ,8 ] .青藏高原强烈隆升使得高原及周边地区沉积巨厚砾岩[8 -9 ] ,内陆干旱化加剧,在我国黄土高原地区堆积典型的第四纪风成黄土-古土壤序列[10 -11 ] . ...
The initiation of Northern Hemisphere glaciation
2
1994
... 新生代以来,全球气候至少经历了始新世-渐新世之交、中中新世和上新世末期的三次台阶式降温[1 -2 ] .上新世-更新世转型是上新世的温暖气候向更新世冰期-间冰期气候转换的重要时段,同时也是现今地质-地貌格局、气候、生态系统和极地冰盖最终稳定形成的重要演化期[1 ,3 -8 ] .晚上新世末北半球冰盖快速扩张的驱动机制一直是讨论的焦点:有研究认为巴拿马海峡的关闭或晚上新世以来大气CO2 浓度减少控制着北极冰盖发育,更有学者提出构造隆升对北极冰盖演化和全球变冷起到驱动作用,指出青藏高原隆升对全球气候变化的重要性[3 ,8 ] .青藏高原强烈隆升使得高原及周边地区沉积巨厚砾岩[8 -9 ] ,内陆干旱化加剧,在我国黄土高原地区堆积典型的第四纪风成黄土-古土壤序列[10 -11 ] . ...
... [3 ,8 ].青藏高原强烈隆升使得高原及周边地区沉积巨厚砾岩[8 -9 ] ,内陆干旱化加剧,在我国黄土高原地区堆积典型的第四纪风成黄土-古土壤序列[10 -11 ] . ...
Evolution of Asian monsoons and phased uplift of the Himalaya-Tibetan plateau since Late Miocene times
1
2001
... 目前关于高原隆升过程、地貌演化、季风演化和亚洲内陆干旱化等方面均取得了重要研究成果[5 ,8 ] ,但关于上新世-更新世转型期的C3 /C4 植被演化历史仍存在不同观点.如来自北太平洋钻孔单体碳同位素证据表明,中国北方C4 植被在12 Ma左右已达最高水平,之后持续减少并在上新世-更新世转型期降至较低水平[12 ] ;这也得到中国南海钻孔正构烷烃δ 13 C结果支持[13 ] .然而,同纬度日本海钻孔黑碳δ 13 C揭示C4 植被在5.3 Ma才开始显著扩张,并在上新世-更新世转型期明显增加[14 ] .同样,华北平原G3钻孔有机碳δ 13 C记录也支持这样的观点:C4 植被从4 Ma开始扩张,并在上新世-更新世转型期达到最高水平[15 ] .除大尺度空间差异外,黄土高原东西部地区C4 植被演化也存在差异.黄土高原中部的闫峪剖面有机碳和碳酸盐同位素结果显示:早更新世时期C4 植被仍处于较低水平,直至1.6 Ma和0.43 Ma才逐步扩张[16 ] .而西峰剖面有机碳同位素结果则显示,红黏土向黄土沉积转变时δ 13 C突然偏正,指示了上新世-更新世转型期C4 植被扩张[17 ] .而青藏高原东北缘临夏盆地哺乳动物牙釉δ 13 C结果表明:2~3 Ma之前发育纯C3 植被,直至上新世-更新世转型期C4 植被才开始出现[18 -19 ] .由此可见,上新世-更新世转型期中国北方C4 植被演化仍存较大争议.另外,前述长尺度C3 /C4 植被演化研究侧重于C4 植被扩张历史,分辨率问题使得关键时期的演化细节缺失,需要更多证据来进一步澄清前述科学问题. ...
Extraordinarily thick-boned fish linked to the aridification of the Qaidam Basin (northern Tibetan Plateau)
0
2008
Out of Tibet: Pliocene woolly rhino suggests high-plateau origin of ice age megaherbivores
0
2011
Late Miocene-Quaternary rapid stepwise uplift of the NE Tibetan Plateau and its effects on climatic and environmental changes
4
2014
... 新生代以来,全球气候至少经历了始新世-渐新世之交、中中新世和上新世末期的三次台阶式降温[1 -2 ] .上新世-更新世转型是上新世的温暖气候向更新世冰期-间冰期气候转换的重要时段,同时也是现今地质-地貌格局、气候、生态系统和极地冰盖最终稳定形成的重要演化期[1 ,3 -8 ] .晚上新世末北半球冰盖快速扩张的驱动机制一直是讨论的焦点:有研究认为巴拿马海峡的关闭或晚上新世以来大气CO2 浓度减少控制着北极冰盖发育,更有学者提出构造隆升对北极冰盖演化和全球变冷起到驱动作用,指出青藏高原隆升对全球气候变化的重要性[3 ,8 ] .青藏高原强烈隆升使得高原及周边地区沉积巨厚砾岩[8 -9 ] ,内陆干旱化加剧,在我国黄土高原地区堆积典型的第四纪风成黄土-古土壤序列[10 -11 ] . ...
... ,8 ].青藏高原强烈隆升使得高原及周边地区沉积巨厚砾岩[8 -9 ] ,内陆干旱化加剧,在我国黄土高原地区堆积典型的第四纪风成黄土-古土壤序列[10 -11 ] . ...
... [8 -9 ],内陆干旱化加剧,在我国黄土高原地区堆积典型的第四纪风成黄土-古土壤序列[10 -11 ] . ...
... 目前关于高原隆升过程、地貌演化、季风演化和亚洲内陆干旱化等方面均取得了重要研究成果[5 ,8 ] ,但关于上新世-更新世转型期的C3 /C4 植被演化历史仍存在不同观点.如来自北太平洋钻孔单体碳同位素证据表明,中国北方C4 植被在12 Ma左右已达最高水平,之后持续减少并在上新世-更新世转型期降至较低水平[12 ] ;这也得到中国南海钻孔正构烷烃δ 13 C结果支持[13 ] .然而,同纬度日本海钻孔黑碳δ 13 C揭示C4 植被在5.3 Ma才开始显著扩张,并在上新世-更新世转型期明显增加[14 ] .同样,华北平原G3钻孔有机碳δ 13 C记录也支持这样的观点:C4 植被从4 Ma开始扩张,并在上新世-更新世转型期达到最高水平[15 ] .除大尺度空间差异外,黄土高原东西部地区C4 植被演化也存在差异.黄土高原中部的闫峪剖面有机碳和碳酸盐同位素结果显示:早更新世时期C4 植被仍处于较低水平,直至1.6 Ma和0.43 Ma才逐步扩张[16 ] .而西峰剖面有机碳同位素结果则显示,红黏土向黄土沉积转变时δ 13 C突然偏正,指示了上新世-更新世转型期C4 植被扩张[17 ] .而青藏高原东北缘临夏盆地哺乳动物牙釉δ 13 C结果表明:2~3 Ma之前发育纯C3 植被,直至上新世-更新世转型期C4 植被才开始出现[18 -19 ] .由此可见,上新世-更新世转型期中国北方C4 植被演化仍存较大争议.另外,前述长尺度C3 /C4 植被演化研究侧重于C4 植被扩张历史,分辨率问题使得关键时期的演化细节缺失,需要更多证据来进一步澄清前述科学问题. ...
Late Pliocene establishment of exorheic drainage in the northeastern Tibetan Plateau as evidenced by the Wuquan Formation in the Lanzhou Basin
2
2018
... 新生代以来,全球气候至少经历了始新世-渐新世之交、中中新世和上新世末期的三次台阶式降温[1 -2 ] .上新世-更新世转型是上新世的温暖气候向更新世冰期-间冰期气候转换的重要时段,同时也是现今地质-地貌格局、气候、生态系统和极地冰盖最终稳定形成的重要演化期[1 ,3 -8 ] .晚上新世末北半球冰盖快速扩张的驱动机制一直是讨论的焦点:有研究认为巴拿马海峡的关闭或晚上新世以来大气CO2 浓度减少控制着北极冰盖发育,更有学者提出构造隆升对北极冰盖演化和全球变冷起到驱动作用,指出青藏高原隆升对全球气候变化的重要性[3 ,8 ] .青藏高原强烈隆升使得高原及周边地区沉积巨厚砾岩[8 -9 ] ,内陆干旱化加剧,在我国黄土高原地区堆积典型的第四纪风成黄土-古土壤序列[10 -11 ] . ...
... 青藏高原东北缘作为高原向东北方向生长扩展的前缘部位,晚新生代以来构造活动显著;同时,地理位置独特,对区域气候和环境变化较为敏感.因此,该区是探讨青藏高原隆升过程和相关生态环境变化的理想地区[20 -21 ] .兰州盆地作为陇中盆地的次级子盆地,沉积序列较为完整,近年来围绕上新统-更新统五泉砾岩和黄土地层学方面开展了大量工作,研究揭示上新世以来兰州地区经历了强烈的构造活动,同时也在皋兰山地区堆积了连续的上新世红黏土-第四纪黄土序列[9 ,11 ] ,为探讨晚上新世-第四纪的构造隆升和古环境演变提供了理想材料.为系统厘清高原东北缘上新世-更新世转型期的古生态演化问题,同时也为弥补西部黄土高原地区上新世-更新世转型期长链正构烷烃碳同位素研究的不足,本文以皋兰山钻孔上新世-更新世转型期的红黏土-黄土岩芯(深度265.7~230.0 m)为研究对象(图1 ),在已有黄土高精度磁性地层学基础上,详细开展了正构烷烃分布、总有机碳和长链正构烷烃碳同位素的有机地球化学分析,恢复了兰州地区上新世-更新世转型期(3.0~2.2 Ma)的C3 /C4 植被演化历史,以揭示高原东北缘兰州地区上新世-更新世过渡时期的古生态环境演变规律,进而尝试探讨该区晚上新世-早第四纪生态转型的可能原因. ...
Stepwise expansion of desert environment across northern China in the past 3.5 Ma and implications for monsoon evolution
1
2005
... 新生代以来,全球气候至少经历了始新世-渐新世之交、中中新世和上新世末期的三次台阶式降温[1 -2 ] .上新世-更新世转型是上新世的温暖气候向更新世冰期-间冰期气候转换的重要时段,同时也是现今地质-地貌格局、气候、生态系统和极地冰盖最终稳定形成的重要演化期[1 ,3 -8 ] .晚上新世末北半球冰盖快速扩张的驱动机制一直是讨论的焦点:有研究认为巴拿马海峡的关闭或晚上新世以来大气CO2 浓度减少控制着北极冰盖发育,更有学者提出构造隆升对北极冰盖演化和全球变冷起到驱动作用,指出青藏高原隆升对全球气候变化的重要性[3 ,8 ] .青藏高原强烈隆升使得高原及周边地区沉积巨厚砾岩[8 -9 ] ,内陆干旱化加剧,在我国黄土高原地区堆积典型的第四纪风成黄土-古土壤序列[10 -11 ] . ...
Magnetostratigraphy and palaeoclimatic significance of the Late Pliocene red clay‐Quaternary loess sequence in the Lanzhou Basin, western Chinese Loess Plateau
7
2020
... 新生代以来,全球气候至少经历了始新世-渐新世之交、中中新世和上新世末期的三次台阶式降温[1 -2 ] .上新世-更新世转型是上新世的温暖气候向更新世冰期-间冰期气候转换的重要时段,同时也是现今地质-地貌格局、气候、生态系统和极地冰盖最终稳定形成的重要演化期[1 ,3 -8 ] .晚上新世末北半球冰盖快速扩张的驱动机制一直是讨论的焦点:有研究认为巴拿马海峡的关闭或晚上新世以来大气CO2 浓度减少控制着北极冰盖发育,更有学者提出构造隆升对北极冰盖演化和全球变冷起到驱动作用,指出青藏高原隆升对全球气候变化的重要性[3 ,8 ] .青藏高原强烈隆升使得高原及周边地区沉积巨厚砾岩[8 -9 ] ,内陆干旱化加剧,在我国黄土高原地区堆积典型的第四纪风成黄土-古土壤序列[10 -11 ] . ...
... 青藏高原东北缘作为高原向东北方向生长扩展的前缘部位,晚新生代以来构造活动显著;同时,地理位置独特,对区域气候和环境变化较为敏感.因此,该区是探讨青藏高原隆升过程和相关生态环境变化的理想地区[20 -21 ] .兰州盆地作为陇中盆地的次级子盆地,沉积序列较为完整,近年来围绕上新统-更新统五泉砾岩和黄土地层学方面开展了大量工作,研究揭示上新世以来兰州地区经历了强烈的构造活动,同时也在皋兰山地区堆积了连续的上新世红黏土-第四纪黄土序列[9 ,11 ] ,为探讨晚上新世-第四纪的构造隆升和古环境演变提供了理想材料.为系统厘清高原东北缘上新世-更新世转型期的古生态演化问题,同时也为弥补西部黄土高原地区上新世-更新世转型期长链正构烷烃碳同位素研究的不足,本文以皋兰山钻孔上新世-更新世转型期的红黏土-黄土岩芯(深度265.7~230.0 m)为研究对象(图1 ),在已有黄土高精度磁性地层学基础上,详细开展了正构烷烃分布、总有机碳和长链正构烷烃碳同位素的有机地球化学分析,恢复了兰州地区上新世-更新世转型期(3.0~2.2 Ma)的C3 /C4 植被演化历史,以揭示高原东北缘兰州地区上新世-更新世过渡时期的古生态环境演变规律,进而尝试探讨该区晚上新世-早第四纪生态转型的可能原因. ...
... 晚新生代青藏高原的隆升使兰州盆地新生代红层发生变形,其上覆盖厚度超过百余米的五泉砾岩[23 ] ;与此同时,在地形较为平坦的台地(皋兰山民族村)开始堆积风成红黏土-黄土序列[11 ] .兰州地区临近粉尘源区,且北支西风急流在高原北部绕流产生了兰州小高压[24 ] .因此,本区发育较厚的第四纪黄土序列,为区域古环境演化研究提供了高分辨率的理想材料. ...
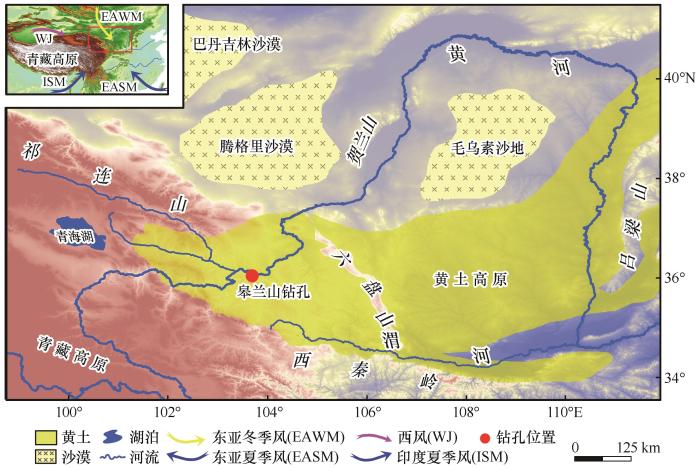
... 皋兰山钻孔(36°01′ N,103°51′ E,海拔2 120 m,图1 )位于兰州盆地南缘皋兰山民族村附近,共获取了黄土-红黏土岩芯样品265.7 m(取芯率高于95%),主要为棕黄色粉砂质黄土和褐红色古土壤互层.本文首先依据皋兰山钻孔高精度磁性地层年代选取绝对控制点[11 ] ,再结合黄土高原黄土-古土壤地层年代框架[25 ] ,利用线性内差法获得皋兰山钻孔底部的深度-年代模型.为探讨兰州盆地上新世-更新世转型期(3.0~2.2 Ma)C3 /C4 植被演化历史,本文选取265.7~230.0 m红黏土-黄土岩芯为研究对象,挑取70块原生样品进行生物标志化合物实验(间距0.5 m). ...
... 如前所述,典型黄土层L32和L33的多数样品正构烷烃呈双鼓包分布(图2 ),表明曾遭受明显的微生物降解[45 -46 ] .而黄土地层和粒度等证据也表明:L32和L33时期,源区和沉积区气候寒冷干燥,均不利于正构烷烃降解;黄土层粒径整体较粗,显示冬季风较为强盛[11 ] .同时,黄土堆积区气候干旱,不利于植被发育.由此推测,降解产物UCM主要为源区母岩输入贡献.事实上,TOC和δ 13 Corg 指标最呈规律性变化的层位正是正构烷烃表现出外源输入特征的L32和L33黄土层(图3 ).这直接支持皋兰山底部黄土层有机碳受外源输入影响的看法.此外,该时段磁化率也表现出随粒径变粗而增大趋势[47 ] .这一证据亦支持源区磁性矿物输入增多的观点.因此,皋兰山样品的TOC高值和δ 13 Corg 的偏负很可能是外源输入导致的假象. ...
...
图5 为端元法校正之后的结果,据此可将兰州地区皋兰山上新世-更新世转型期(3.0~2.2 Ma) C
3 /C
4 植被演化划分为两个阶段:
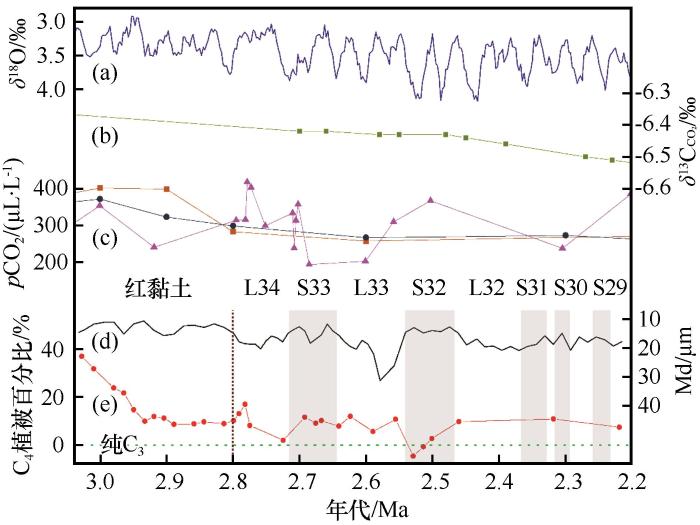
图5 皋兰山C<sub>4</sub>植被含量与深海氧同位素(a)、大气<span class="formulaText"><inline-formula><math id="M6"><msup><mrow><mi>δ</mi></mrow><mrow><mn mathvariant="normal">13</mn></mrow></msup><msub><mrow><mi mathvariant="normal">C</mi></mrow><mrow><mi mathvariant="normal">C</mi><msub><mrow><mi mathvariant="normal">O</mi></mrow><mrow><mn mathvariant="normal">2</mn></mrow></msub></mrow></msub></math></span></inline-formula></span> (b)、<i>p</i>CO<sub>2</sub> (c)和中值粒径(d)综合对比图 Comparisons between deep ocean <i>δ</i><sup>18</sup>O(a) from Lisieki and Raymo<sup>[<xref ref-type="bibr" rid="R56">56</xref>]</sup>, atmosphere <span class="formulaText"><inline-formula><math id="M7"><msup><mrow><mi>δ</mi></mrow><mrow><mn mathvariant="normal">13</mn></mrow></msup><msub><mrow><mi mathvariant="normal">C</mi></mrow><mrow><mi mathvariant="normal">C</mi><msub><mrow><mi mathvariant="normal">O</mi></mrow><mrow><mn mathvariant="normal">2</mn></mrow></msub></mrow></msub></math></span></inline-formula></span> (b) from Tipple et al.<sup>[<xref ref-type="bibr" rid="R51">51</xref>]</sup>, atmosphere <i>p</i>CO<sub>2</sub> (c) from Beerling and Royer<sup>[<xref ref-type="bibr" rid="R52">52</xref>]</sup> & Bartoli et al.<sup>[<xref ref-type="bibr" rid="R54">54</xref>]</sup>, median diameter (d) from Guo et al.<sup>[<xref ref-type="bibr" rid="R11">11</xref>]</sup>, and C<sub>4</sub> content (e). Atmosphere <i>p</i>CO<sub>2</sub> based on <i>δ</i><sup>11</sup>B, phytoplankton<sup>[<xref ref-type="bibr" rid="R52">52</xref>]</sup>, and <i>δ</i><sup>11</sup>B<sup>[<xref ref-type="bibr" rid="R55">55</xref>]</sup> are outlined by the orange, black, and pink plots and line, respectively Fig.5 ![]()
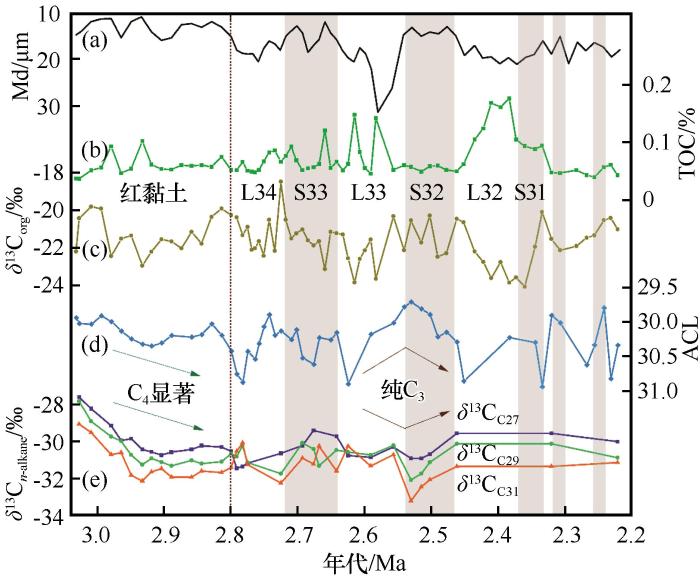
第一阶段:3.0~2.9 Ma时段,主体为红黏土沉积,C4 植被相对比例最高.上新世大暖期结束时(3.0 Ma),δ 13 Cn -alkane 整体较为偏正[图(3)和(5)],远高于C3 植被δ 13 Cn -alkane 分布范围(-39‰~-32‰).ACL值较低,指示气候整体温暖,但ACL逐渐增大,支持气候逐渐变冷.与此同时,ACL与δ 13 Cn -alkane 呈反相位变化,即气候越冷干,δ 13 Cn -alkane 值逐渐偏负,与黄土高原东部的C4 模式一致.据此推断:此时气候条件仍相对温暖湿润,适宜C4 植被发育,δ 13 Cn -alkane 由C4 植被相对丰度变化主导.定量重建结果表明,此时段C4 植被比例最高可达40%(图5 ).2.9~2.8 Ma,ACL值逐渐降低,磁化率较之前明显增加[47 ] ,表明此时降水较之前有所增多,相对充沛.而兰州位于季风影响的边缘区域,雨热同期主要集中在生长季(夏季).因此,强降水也意味着季节性加强,更适宜C4 生长.但此时C4 植被仍稳定在低水平,说明降水不是C4 植被减少的主要因素.同时,此时段p CO2 略微降低[52 ,55 ] (图5 ),也更适宜C4 发育.排除前述因子后,C4 植被生长的限制因素极可能为温度.研究表明,上新世温暖期(3.3~3.0 Ma)结束后,氧同位素揭示的全球冰量显著扩张[56 ] ,温度进一步下降.这可能逐渐突破了C4 植被生长阈值,进而导致兰州皋兰山C4 植被快速减少. ...
... 2.8 Ma左右,沉积相从红黏土转变为黄土,北半球冰盖开始扩张,ACL和δ 13 Cn -alkane 波动明显,黄土中值粒径增加指示冬季风加强[11 ] ,气候转冷干.值得注意的是,δ 13 CC27 与δ 13 CC31 发生了截然不同的变化(图3 ).这是C3 模式和C4 模式都难以解释的.据此本文认为,可能是因为皋兰山C3 植被与C4 植被的主峰碳数不同,而较为贫乏的C4 植被不足以掩盖C3 植被δ 13 Cn -alkane 对气候响应的信号,从而导致了这种分异.δ 13 CC27 与δ 13 CC31 反向变化一直持续到2.6 Ma前后,标志着此时段气候较之前更为恶化.与第一阶段类似的是:S33阶段高磁化率[47 ] 的丰沛降水也未能显著影响C4 含量,表明降水量亦不是C4 发育的控制因子.因此,此时期C4 植被减少的主要因素仍是温度. ...
Aeolian n -alkane isotopic evidence from North Pacific for a Late Miocene decline of C4 plant in the arid Asian interior
1
2012
... 目前关于高原隆升过程、地貌演化、季风演化和亚洲内陆干旱化等方面均取得了重要研究成果[5 ,8 ] ,但关于上新世-更新世转型期的C3 /C4 植被演化历史仍存在不同观点.如来自北太平洋钻孔单体碳同位素证据表明,中国北方C4 植被在12 Ma左右已达最高水平,之后持续减少并在上新世-更新世转型期降至较低水平[12 ] ;这也得到中国南海钻孔正构烷烃δ 13 C结果支持[13 ] .然而,同纬度日本海钻孔黑碳δ 13 C揭示C4 植被在5.3 Ma才开始显著扩张,并在上新世-更新世转型期明显增加[14 ] .同样,华北平原G3钻孔有机碳δ 13 C记录也支持这样的观点:C4 植被从4 Ma开始扩张,并在上新世-更新世转型期达到最高水平[15 ] .除大尺度空间差异外,黄土高原东西部地区C4 植被演化也存在差异.黄土高原中部的闫峪剖面有机碳和碳酸盐同位素结果显示:早更新世时期C4 植被仍处于较低水平,直至1.6 Ma和0.43 Ma才逐步扩张[16 ] .而西峰剖面有机碳同位素结果则显示,红黏土向黄土沉积转变时δ 13 C突然偏正,指示了上新世-更新世转型期C4 植被扩张[17 ] .而青藏高原东北缘临夏盆地哺乳动物牙釉δ 13 C结果表明:2~3 Ma之前发育纯C3 植被,直至上新世-更新世转型期C4 植被才开始出现[18 -19 ] .由此可见,上新世-更新世转型期中国北方C4 植被演化仍存较大争议.另外,前述长尺度C3 /C4 植被演化研究侧重于C4 植被扩张历史,分辨率问题使得关键时期的演化细节缺失,需要更多证据来进一步澄清前述科学问题. ...
New sedimentary evidence reveals a unique history of C4 biomass in continental East Asia since the early Miocene
1
2017
... 目前关于高原隆升过程、地貌演化、季风演化和亚洲内陆干旱化等方面均取得了重要研究成果[5 ,8 ] ,但关于上新世-更新世转型期的C3 /C4 植被演化历史仍存在不同观点.如来自北太平洋钻孔单体碳同位素证据表明,中国北方C4 植被在12 Ma左右已达最高水平,之后持续减少并在上新世-更新世转型期降至较低水平[12 ] ;这也得到中国南海钻孔正构烷烃δ 13 C结果支持[13 ] .然而,同纬度日本海钻孔黑碳δ 13 C揭示C4 植被在5.3 Ma才开始显著扩张,并在上新世-更新世转型期明显增加[14 ] .同样,华北平原G3钻孔有机碳δ 13 C记录也支持这样的观点:C4 植被从4 Ma开始扩张,并在上新世-更新世转型期达到最高水平[15 ] .除大尺度空间差异外,黄土高原东西部地区C4 植被演化也存在差异.黄土高原中部的闫峪剖面有机碳和碳酸盐同位素结果显示:早更新世时期C4 植被仍处于较低水平,直至1.6 Ma和0.43 Ma才逐步扩张[16 ] .而西峰剖面有机碳同位素结果则显示,红黏土向黄土沉积转变时δ 13 C突然偏正,指示了上新世-更新世转型期C4 植被扩张[17 ] .而青藏高原东北缘临夏盆地哺乳动物牙釉δ 13 C结果表明:2~3 Ma之前发育纯C3 植被,直至上新世-更新世转型期C4 植被才开始出现[18 -19 ] .由此可见,上新世-更新世转型期中国北方C4 植被演化仍存较大争议.另外,前述长尺度C3 /C4 植被演化研究侧重于C4 植被扩张历史,分辨率问题使得关键时期的演化细节缺失,需要更多证据来进一步澄清前述科学问题. ...
Increased seasonality and aridity drove the C4 plant expansion in Central Asia since the Miocene-Pliocene boundary
3
2018
... 目前关于高原隆升过程、地貌演化、季风演化和亚洲内陆干旱化等方面均取得了重要研究成果[5 ,8 ] ,但关于上新世-更新世转型期的C3 /C4 植被演化历史仍存在不同观点.如来自北太平洋钻孔单体碳同位素证据表明,中国北方C4 植被在12 Ma左右已达最高水平,之后持续减少并在上新世-更新世转型期降至较低水平[12 ] ;这也得到中国南海钻孔正构烷烃δ 13 C结果支持[13 ] .然而,同纬度日本海钻孔黑碳δ 13 C揭示C4 植被在5.3 Ma才开始显著扩张,并在上新世-更新世转型期明显增加[14 ] .同样,华北平原G3钻孔有机碳δ 13 C记录也支持这样的观点:C4 植被从4 Ma开始扩张,并在上新世-更新世转型期达到最高水平[15 ] .除大尺度空间差异外,黄土高原东西部地区C4 植被演化也存在差异.黄土高原中部的闫峪剖面有机碳和碳酸盐同位素结果显示:早更新世时期C4 植被仍处于较低水平,直至1.6 Ma和0.43 Ma才逐步扩张[16 ] .而西峰剖面有机碳同位素结果则显示,红黏土向黄土沉积转变时δ 13 C突然偏正,指示了上新世-更新世转型期C4 植被扩张[17 ] .而青藏高原东北缘临夏盆地哺乳动物牙釉δ 13 C结果表明:2~3 Ma之前发育纯C3 植被,直至上新世-更新世转型期C4 植被才开始出现[18 -19 ] .由此可见,上新世-更新世转型期中国北方C4 植被演化仍存较大争议.另外,前述长尺度C3 /C4 植被演化研究侧重于C4 植被扩张历史,分辨率问题使得关键时期的演化细节缺失,需要更多证据来进一步澄清前述科学问题. ...
... C4 植被的出现可以追溯到32~30 Ma,其大幅扩张一般认为发生在晚中新世[57 ] .来自巴基斯坦Siwalik沉积有机质和无机碳[58 ] 、北美哺乳动物牙釉化石[59 ] 、南美东部钻孔无机碳和动物化石[60 ] 和东非钻孔中动物化石[61 ] 等研究结果均支持这一说法:晚中新世7~8 Ma左右全球发生C4 植被的显著扩张.因此,C4 植被扩张是由大气CO2 浓度降低驱动的全球生态事件[62 ] .21世纪以来,越来越多的记录表明,全球C4 植被扩张的时间和范围并不一致:非洲东北部正构烷烃δ 13 C表明,C4 植被在11 Ma业已达到较高水平[50 ] ;新疆塔里木盆地钻孔有机碳δ 13 C显示,C4 植被在5.3 Ma显著增加[63 ] .这也得到日本海钻孔正构烷烃δ 13 C和内蒙古哺乳动物牙釉化石δ 13 C结果的支持[14 ,64 ] .近来,Lu等[15 ] 汇总全球记录后提出,早上新世存在一次全球性C4 扩张事件.然而,澳大利亚近海钻孔正构烷烃δ 13 C表明,C4 植被直至3.5 Ma才开始扩张[65 ] .同时,太平洋近赤道地区海洋钻孔长链脂肪酸δ 13 C[66 ] 和黄土高原西部临夏盆地牙釉δ 13 C[18 -19 ] 指示,C4 植被直至早更新世后才显著增加.此外,黄土高原东部灵台、蓝田剖面无机碳δ 13 C指示,C4 植被经历了2.9~2.7 Ma、1.3~0.9 Ma、0.6 Ma三次反复扩张[67 ] .基于全球各地C4 植被演化差异,学者们将控制C4 植被演化的因素归结于干旱程度[14 ,63 ,68 -69 ] 、暖季温度和降水[67 ,70 -71 ] 和野火[72 -73 ] 等区域环境因素.由此可见,关于晚中新世以来C4 植被演化的时间和机制等的认识仍未统一. ...
... [14 ,63 ,68 -69 ]、暖季温度和降水[67 ,70 -71 ] 和野火[72 -73 ] 等区域环境因素.由此可见,关于晚中新世以来C4 植被演化的时间和机制等的认识仍未统一. ...
The Early Pliocene global expansion of C4 grasslands: A new organic carbon-isotopic dataset from the north China plain
2
2020
... 目前关于高原隆升过程、地貌演化、季风演化和亚洲内陆干旱化等方面均取得了重要研究成果[5 ,8 ] ,但关于上新世-更新世转型期的C3 /C4 植被演化历史仍存在不同观点.如来自北太平洋钻孔单体碳同位素证据表明,中国北方C4 植被在12 Ma左右已达最高水平,之后持续减少并在上新世-更新世转型期降至较低水平[12 ] ;这也得到中国南海钻孔正构烷烃δ 13 C结果支持[13 ] .然而,同纬度日本海钻孔黑碳δ 13 C揭示C4 植被在5.3 Ma才开始显著扩张,并在上新世-更新世转型期明显增加[14 ] .同样,华北平原G3钻孔有机碳δ 13 C记录也支持这样的观点:C4 植被从4 Ma开始扩张,并在上新世-更新世转型期达到最高水平[15 ] .除大尺度空间差异外,黄土高原东西部地区C4 植被演化也存在差异.黄土高原中部的闫峪剖面有机碳和碳酸盐同位素结果显示:早更新世时期C4 植被仍处于较低水平,直至1.6 Ma和0.43 Ma才逐步扩张[16 ] .而西峰剖面有机碳同位素结果则显示,红黏土向黄土沉积转变时δ 13 C突然偏正,指示了上新世-更新世转型期C4 植被扩张[17 ] .而青藏高原东北缘临夏盆地哺乳动物牙釉δ 13 C结果表明:2~3 Ma之前发育纯C3 植被,直至上新世-更新世转型期C4 植被才开始出现[18 -19 ] .由此可见,上新世-更新世转型期中国北方C4 植被演化仍存较大争议.另外,前述长尺度C3 /C4 植被演化研究侧重于C4 植被扩张历史,分辨率问题使得关键时期的演化细节缺失,需要更多证据来进一步澄清前述科学问题. ...
... C4 植被的出现可以追溯到32~30 Ma,其大幅扩张一般认为发生在晚中新世[57 ] .来自巴基斯坦Siwalik沉积有机质和无机碳[58 ] 、北美哺乳动物牙釉化石[59 ] 、南美东部钻孔无机碳和动物化石[60 ] 和东非钻孔中动物化石[61 ] 等研究结果均支持这一说法:晚中新世7~8 Ma左右全球发生C4 植被的显著扩张.因此,C4 植被扩张是由大气CO2 浓度降低驱动的全球生态事件[62 ] .21世纪以来,越来越多的记录表明,全球C4 植被扩张的时间和范围并不一致:非洲东北部正构烷烃δ 13 C表明,C4 植被在11 Ma业已达到较高水平[50 ] ;新疆塔里木盆地钻孔有机碳δ 13 C显示,C4 植被在5.3 Ma显著增加[63 ] .这也得到日本海钻孔正构烷烃δ 13 C和内蒙古哺乳动物牙釉化石δ 13 C结果的支持[14 ,64 ] .近来,Lu等[15 ] 汇总全球记录后提出,早上新世存在一次全球性C4 扩张事件.然而,澳大利亚近海钻孔正构烷烃δ 13 C表明,C4 植被直至3.5 Ma才开始扩张[65 ] .同时,太平洋近赤道地区海洋钻孔长链脂肪酸δ 13 C[66 ] 和黄土高原西部临夏盆地牙釉δ 13 C[18 -19 ] 指示,C4 植被直至早更新世后才显著增加.此外,黄土高原东部灵台、蓝田剖面无机碳δ 13 C指示,C4 植被经历了2.9~2.7 Ma、1.3~0.9 Ma、0.6 Ma三次反复扩张[67 ] .基于全球各地C4 植被演化差异,学者们将控制C4 植被演化的因素归结于干旱程度[14 ,63 ,68 -69 ] 、暖季温度和降水[67 ,70 -71 ] 和野火[72 -73 ] 等区域环境因素.由此可见,关于晚中新世以来C4 植被演化的时间和机制等的认识仍未统一. ...
Stepwise expansions of C4 biomass and enhanced seasonal precipitation and regional aridity during the Quaternary on the southern Chinese Loess Plateau
1
2012
... 目前关于高原隆升过程、地貌演化、季风演化和亚洲内陆干旱化等方面均取得了重要研究成果[5 ,8 ] ,但关于上新世-更新世转型期的C3 /C4 植被演化历史仍存在不同观点.如来自北太平洋钻孔单体碳同位素证据表明,中国北方C4 植被在12 Ma左右已达最高水平,之后持续减少并在上新世-更新世转型期降至较低水平[12 ] ;这也得到中国南海钻孔正构烷烃δ 13 C结果支持[13 ] .然而,同纬度日本海钻孔黑碳δ 13 C揭示C4 植被在5.3 Ma才开始显著扩张,并在上新世-更新世转型期明显增加[14 ] .同样,华北平原G3钻孔有机碳δ 13 C记录也支持这样的观点:C4 植被从4 Ma开始扩张,并在上新世-更新世转型期达到最高水平[15 ] .除大尺度空间差异外,黄土高原东西部地区C4 植被演化也存在差异.黄土高原中部的闫峪剖面有机碳和碳酸盐同位素结果显示:早更新世时期C4 植被仍处于较低水平,直至1.6 Ma和0.43 Ma才逐步扩张[16 ] .而西峰剖面有机碳同位素结果则显示,红黏土向黄土沉积转变时δ 13 C突然偏正,指示了上新世-更新世转型期C4 植被扩张[17 ] .而青藏高原东北缘临夏盆地哺乳动物牙釉δ 13 C结果表明:2~3 Ma之前发育纯C3 植被,直至上新世-更新世转型期C4 植被才开始出现[18 -19 ] .由此可见,上新世-更新世转型期中国北方C4 植被演化仍存较大争议.另外,前述长尺度C3 /C4 植被演化研究侧重于C4 植被扩张历史,分辨率问题使得关键时期的演化细节缺失,需要更多证据来进一步澄清前述科学问题. ...
西峰红黏土/黄土剖面有机碳同位素研究的初步结果
1
2012
... 目前关于高原隆升过程、地貌演化、季风演化和亚洲内陆干旱化等方面均取得了重要研究成果[5 ,8 ] ,但关于上新世-更新世转型期的C3 /C4 植被演化历史仍存在不同观点.如来自北太平洋钻孔单体碳同位素证据表明,中国北方C4 植被在12 Ma左右已达最高水平,之后持续减少并在上新世-更新世转型期降至较低水平[12 ] ;这也得到中国南海钻孔正构烷烃δ 13 C结果支持[13 ] .然而,同纬度日本海钻孔黑碳δ 13 C揭示C4 植被在5.3 Ma才开始显著扩张,并在上新世-更新世转型期明显增加[14 ] .同样,华北平原G3钻孔有机碳δ 13 C记录也支持这样的观点:C4 植被从4 Ma开始扩张,并在上新世-更新世转型期达到最高水平[15 ] .除大尺度空间差异外,黄土高原东西部地区C4 植被演化也存在差异.黄土高原中部的闫峪剖面有机碳和碳酸盐同位素结果显示:早更新世时期C4 植被仍处于较低水平,直至1.6 Ma和0.43 Ma才逐步扩张[16 ] .而西峰剖面有机碳同位素结果则显示,红黏土向黄土沉积转变时δ 13 C突然偏正,指示了上新世-更新世转型期C4 植被扩张[17 ] .而青藏高原东北缘临夏盆地哺乳动物牙釉δ 13 C结果表明:2~3 Ma之前发育纯C3 植被,直至上新世-更新世转型期C4 植被才开始出现[18 -19 ] .由此可见,上新世-更新世转型期中国北方C4 植被演化仍存较大争议.另外,前述长尺度C3 /C4 植被演化研究侧重于C4 植被扩张历史,分辨率问题使得关键时期的演化细节缺失,需要更多证据来进一步澄清前述科学问题. ...
西峰红黏土/黄土剖面有机碳同位素研究的初步结果
1
2012
... 目前关于高原隆升过程、地貌演化、季风演化和亚洲内陆干旱化等方面均取得了重要研究成果[5 ,8 ] ,但关于上新世-更新世转型期的C3 /C4 植被演化历史仍存在不同观点.如来自北太平洋钻孔单体碳同位素证据表明,中国北方C4 植被在12 Ma左右已达最高水平,之后持续减少并在上新世-更新世转型期降至较低水平[12 ] ;这也得到中国南海钻孔正构烷烃δ 13 C结果支持[13 ] .然而,同纬度日本海钻孔黑碳δ 13 C揭示C4 植被在5.3 Ma才开始显著扩张,并在上新世-更新世转型期明显增加[14 ] .同样,华北平原G3钻孔有机碳δ 13 C记录也支持这样的观点:C4 植被从4 Ma开始扩张,并在上新世-更新世转型期达到最高水平[15 ] .除大尺度空间差异外,黄土高原东西部地区C4 植被演化也存在差异.黄土高原中部的闫峪剖面有机碳和碳酸盐同位素结果显示:早更新世时期C4 植被仍处于较低水平,直至1.6 Ma和0.43 Ma才逐步扩张[16 ] .而西峰剖面有机碳同位素结果则显示,红黏土向黄土沉积转变时δ 13 C突然偏正,指示了上新世-更新世转型期C4 植被扩张[17 ] .而青藏高原东北缘临夏盆地哺乳动物牙釉δ 13 C结果表明:2~3 Ma之前发育纯C3 植被,直至上新世-更新世转型期C4 植被才开始出现[18 -19 ] .由此可见,上新世-更新世转型期中国北方C4 植被演化仍存较大争议.另外,前述长尺度C3 /C4 植被演化研究侧重于C4 植被扩张历史,分辨率问题使得关键时期的演化细节缺失,需要更多证据来进一步澄清前述科学问题. ...
A 25 m.y. isotopic record of paleodiet and environmental change from fossil mammals and paleosols from the NE margin of the Tibetan Plateau
3
2005
... 目前关于高原隆升过程、地貌演化、季风演化和亚洲内陆干旱化等方面均取得了重要研究成果[5 ,8 ] ,但关于上新世-更新世转型期的C3 /C4 植被演化历史仍存在不同观点.如来自北太平洋钻孔单体碳同位素证据表明,中国北方C4 植被在12 Ma左右已达最高水平,之后持续减少并在上新世-更新世转型期降至较低水平[12 ] ;这也得到中国南海钻孔正构烷烃δ 13 C结果支持[13 ] .然而,同纬度日本海钻孔黑碳δ 13 C揭示C4 植被在5.3 Ma才开始显著扩张,并在上新世-更新世转型期明显增加[14 ] .同样,华北平原G3钻孔有机碳δ 13 C记录也支持这样的观点:C4 植被从4 Ma开始扩张,并在上新世-更新世转型期达到最高水平[15 ] .除大尺度空间差异外,黄土高原东西部地区C4 植被演化也存在差异.黄土高原中部的闫峪剖面有机碳和碳酸盐同位素结果显示:早更新世时期C4 植被仍处于较低水平,直至1.6 Ma和0.43 Ma才逐步扩张[16 ] .而西峰剖面有机碳同位素结果则显示,红黏土向黄土沉积转变时δ 13 C突然偏正,指示了上新世-更新世转型期C4 植被扩张[17 ] .而青藏高原东北缘临夏盆地哺乳动物牙釉δ 13 C结果表明:2~3 Ma之前发育纯C3 植被,直至上新世-更新世转型期C4 植被才开始出现[18 -19 ] .由此可见,上新世-更新世转型期中国北方C4 植被演化仍存较大争议.另外,前述长尺度C3 /C4 植被演化研究侧重于C4 植被扩张历史,分辨率问题使得关键时期的演化细节缺失,需要更多证据来进一步澄清前述科学问题. ...
... C4 植被的出现可以追溯到32~30 Ma,其大幅扩张一般认为发生在晚中新世[57 ] .来自巴基斯坦Siwalik沉积有机质和无机碳[58 ] 、北美哺乳动物牙釉化石[59 ] 、南美东部钻孔无机碳和动物化石[60 ] 和东非钻孔中动物化石[61 ] 等研究结果均支持这一说法:晚中新世7~8 Ma左右全球发生C4 植被的显著扩张.因此,C4 植被扩张是由大气CO2 浓度降低驱动的全球生态事件[62 ] .21世纪以来,越来越多的记录表明,全球C4 植被扩张的时间和范围并不一致:非洲东北部正构烷烃δ 13 C表明,C4 植被在11 Ma业已达到较高水平[50 ] ;新疆塔里木盆地钻孔有机碳δ 13 C显示,C4 植被在5.3 Ma显著增加[63 ] .这也得到日本海钻孔正构烷烃δ 13 C和内蒙古哺乳动物牙釉化石δ 13 C结果的支持[14 ,64 ] .近来,Lu等[15 ] 汇总全球记录后提出,早上新世存在一次全球性C4 扩张事件.然而,澳大利亚近海钻孔正构烷烃δ 13 C表明,C4 植被直至3.5 Ma才开始扩张[65 ] .同时,太平洋近赤道地区海洋钻孔长链脂肪酸δ 13 C[66 ] 和黄土高原西部临夏盆地牙釉δ 13 C[18 -19 ] 指示,C4 植被直至早更新世后才显著增加.此外,黄土高原东部灵台、蓝田剖面无机碳δ 13 C指示,C4 植被经历了2.9~2.7 Ma、1.3~0.9 Ma、0.6 Ma三次反复扩张[67 ] .基于全球各地C4 植被演化差异,学者们将控制C4 植被演化的因素归结于干旱程度[14 ,63 ,68 -69 ] 、暖季温度和降水[67 ,70 -71 ] 和野火[72 -73 ] 等区域环境因素.由此可见,关于晚中新世以来C4 植被演化的时间和机制等的认识仍未统一. ...
... 截止目前,黄土高原西部地区关于C4 植被演化的记录较少,以临夏盆地哺乳动物化石δ 13 C记录为代表的研究,揭示了中新世以来C4 植被演化的整体框架[18 -19 ] .但这一记录分辨率较低,对C4 扩张事件的细节不能准确刻画.而皋兰山单体碳同位素刚好弥补此时段的不足.本文结果表明,3.0 Ma之前兰州盆地C4 植被占比40%,表明至少3.0 Ma之前发生过C4 扩张,但持续多长等问题仍需更多证据验证;3.0~2.9 Ma时期,兰州地区发生了一次快速的C4 植被减少事件;之后,整体以C3 植被为主.因此,本文研究结果亦不支持早第四纪C4 植被扩张的观点. ...
Strengthening of the East Asian summer monsoon revealed by a shift in seasonal patterns in diet and climate after 2~3 Ma in northwest China
3
2010
... 目前关于高原隆升过程、地貌演化、季风演化和亚洲内陆干旱化等方面均取得了重要研究成果[5 ,8 ] ,但关于上新世-更新世转型期的C3 /C4 植被演化历史仍存在不同观点.如来自北太平洋钻孔单体碳同位素证据表明,中国北方C4 植被在12 Ma左右已达最高水平,之后持续减少并在上新世-更新世转型期降至较低水平[12 ] ;这也得到中国南海钻孔正构烷烃δ 13 C结果支持[13 ] .然而,同纬度日本海钻孔黑碳δ 13 C揭示C4 植被在5.3 Ma才开始显著扩张,并在上新世-更新世转型期明显增加[14 ] .同样,华北平原G3钻孔有机碳δ 13 C记录也支持这样的观点:C4 植被从4 Ma开始扩张,并在上新世-更新世转型期达到最高水平[15 ] .除大尺度空间差异外,黄土高原东西部地区C4 植被演化也存在差异.黄土高原中部的闫峪剖面有机碳和碳酸盐同位素结果显示:早更新世时期C4 植被仍处于较低水平,直至1.6 Ma和0.43 Ma才逐步扩张[16 ] .而西峰剖面有机碳同位素结果则显示,红黏土向黄土沉积转变时δ 13 C突然偏正,指示了上新世-更新世转型期C4 植被扩张[17 ] .而青藏高原东北缘临夏盆地哺乳动物牙釉δ 13 C结果表明:2~3 Ma之前发育纯C3 植被,直至上新世-更新世转型期C4 植被才开始出现[18 -19 ] .由此可见,上新世-更新世转型期中国北方C4 植被演化仍存较大争议.另外,前述长尺度C3 /C4 植被演化研究侧重于C4 植被扩张历史,分辨率问题使得关键时期的演化细节缺失,需要更多证据来进一步澄清前述科学问题. ...
... C4 植被的出现可以追溯到32~30 Ma,其大幅扩张一般认为发生在晚中新世[57 ] .来自巴基斯坦Siwalik沉积有机质和无机碳[58 ] 、北美哺乳动物牙釉化石[59 ] 、南美东部钻孔无机碳和动物化石[60 ] 和东非钻孔中动物化石[61 ] 等研究结果均支持这一说法:晚中新世7~8 Ma左右全球发生C4 植被的显著扩张.因此,C4 植被扩张是由大气CO2 浓度降低驱动的全球生态事件[62 ] .21世纪以来,越来越多的记录表明,全球C4 植被扩张的时间和范围并不一致:非洲东北部正构烷烃δ 13 C表明,C4 植被在11 Ma业已达到较高水平[50 ] ;新疆塔里木盆地钻孔有机碳δ 13 C显示,C4 植被在5.3 Ma显著增加[63 ] .这也得到日本海钻孔正构烷烃δ 13 C和内蒙古哺乳动物牙釉化石δ 13 C结果的支持[14 ,64 ] .近来,Lu等[15 ] 汇总全球记录后提出,早上新世存在一次全球性C4 扩张事件.然而,澳大利亚近海钻孔正构烷烃δ 13 C表明,C4 植被直至3.5 Ma才开始扩张[65 ] .同时,太平洋近赤道地区海洋钻孔长链脂肪酸δ 13 C[66 ] 和黄土高原西部临夏盆地牙釉δ 13 C[18 -19 ] 指示,C4 植被直至早更新世后才显著增加.此外,黄土高原东部灵台、蓝田剖面无机碳δ 13 C指示,C4 植被经历了2.9~2.7 Ma、1.3~0.9 Ma、0.6 Ma三次反复扩张[67 ] .基于全球各地C4 植被演化差异,学者们将控制C4 植被演化的因素归结于干旱程度[14 ,63 ,68 -69 ] 、暖季温度和降水[67 ,70 -71 ] 和野火[72 -73 ] 等区域环境因素.由此可见,关于晚中新世以来C4 植被演化的时间和机制等的认识仍未统一. ...
... 截止目前,黄土高原西部地区关于C4 植被演化的记录较少,以临夏盆地哺乳动物化石δ 13 C记录为代表的研究,揭示了中新世以来C4 植被演化的整体框架[18 -19 ] .但这一记录分辨率较低,对C4 扩张事件的细节不能准确刻画.而皋兰山单体碳同位素刚好弥补此时段的不足.本文结果表明,3.0 Ma之前兰州盆地C4 植被占比40%,表明至少3.0 Ma之前发生过C4 扩张,但持续多长等问题仍需更多证据验证;3.0~2.9 Ma时期,兰州地区发生了一次快速的C4 植被减少事件;之后,整体以C3 植被为主.因此,本文研究结果亦不支持早第四纪C4 植被扩张的观点. ...
What controlled Mid-Late Miocene long-term aridification in Central Asia?Global cooling or Tibetan Plateau uplift: A review
1
2012
... 青藏高原东北缘作为高原向东北方向生长扩展的前缘部位,晚新生代以来构造活动显著;同时,地理位置独特,对区域气候和环境变化较为敏感.因此,该区是探讨青藏高原隆升过程和相关生态环境变化的理想地区[20 -21 ] .兰州盆地作为陇中盆地的次级子盆地,沉积序列较为完整,近年来围绕上新统-更新统五泉砾岩和黄土地层学方面开展了大量工作,研究揭示上新世以来兰州地区经历了强烈的构造活动,同时也在皋兰山地区堆积了连续的上新世红黏土-第四纪黄土序列[9 ,11 ] ,为探讨晚上新世-第四纪的构造隆升和古环境演变提供了理想材料.为系统厘清高原东北缘上新世-更新世转型期的古生态演化问题,同时也为弥补西部黄土高原地区上新世-更新世转型期长链正构烷烃碳同位素研究的不足,本文以皋兰山钻孔上新世-更新世转型期的红黏土-黄土岩芯(深度265.7~230.0 m)为研究对象(图1 ),在已有黄土高精度磁性地层学基础上,详细开展了正构烷烃分布、总有机碳和长链正构烷烃碳同位素的有机地球化学分析,恢复了兰州地区上新世-更新世转型期(3.0~2.2 Ma)的C3 /C4 植被演化历史,以揭示高原东北缘兰州地区上新世-更新世过渡时期的古生态环境演变规律,进而尝试探讨该区晚上新世-早第四纪生态转型的可能原因. ...
Orographic controls on climate and paleoclimate of Asia: Thermal and mechanical roles for the Tibetan Plateau
1
2010
... 青藏高原东北缘作为高原向东北方向生长扩展的前缘部位,晚新生代以来构造活动显著;同时,地理位置独特,对区域气候和环境变化较为敏感.因此,该区是探讨青藏高原隆升过程和相关生态环境变化的理想地区[20 -21 ] .兰州盆地作为陇中盆地的次级子盆地,沉积序列较为完整,近年来围绕上新统-更新统五泉砾岩和黄土地层学方面开展了大量工作,研究揭示上新世以来兰州地区经历了强烈的构造活动,同时也在皋兰山地区堆积了连续的上新世红黏土-第四纪黄土序列[9 ,11 ] ,为探讨晚上新世-第四纪的构造隆升和古环境演变提供了理想材料.为系统厘清高原东北缘上新世-更新世转型期的古生态演化问题,同时也为弥补西部黄土高原地区上新世-更新世转型期长链正构烷烃碳同位素研究的不足,本文以皋兰山钻孔上新世-更新世转型期的红黏土-黄土岩芯(深度265.7~230.0 m)为研究对象(图1 ),在已有黄土高精度磁性地层学基础上,详细开展了正构烷烃分布、总有机碳和长链正构烷烃碳同位素的有机地球化学分析,恢复了兰州地区上新世-更新世转型期(3.0~2.2 Ma)的C3 /C4 植被演化历史,以揭示高原东北缘兰州地区上新世-更新世过渡时期的古生态环境演变规律,进而尝试探讨该区晚上新世-早第四纪生态转型的可能原因. ...
兰州西津黄土-古土壤序列中黏土矿物特征
1
2018
... 兰州盆地作为青藏高原东北缘陇中盆地的次级子盆地,同时也位于黄土高原向青藏高原过渡的关键区域,从自然地理角度讲,兰州地处青藏高寒区、西北干旱区和东部季风区的交汇地带,亦是“季风三角”的枢纽部位所在(图1 ),对环境变化较为敏感,是进行古环境研究的理想区域.兰州现属温带半干旱气候,发育稀疏的灌木和草本植物;年均温10 ℃左右,年均降水约310 mm,主要集中在夏季的6—9月[22 ] . ...
兰州西津黄土-古土壤序列中黏土矿物特征
1
2018
... 兰州盆地作为青藏高原东北缘陇中盆地的次级子盆地,同时也位于黄土高原向青藏高原过渡的关键区域,从自然地理角度讲,兰州地处青藏高寒区、西北干旱区和东部季风区的交汇地带,亦是“季风三角”的枢纽部位所在(图1 ),对环境变化较为敏感,是进行古环境研究的理想区域.兰州现属温带半干旱气候,发育稀疏的灌木和草本植物;年均温10 ℃左右,年均降水约310 mm,主要集中在夏季的6—9月[22 ] . ...
Pedogenic components of Xijin loess from the western Chinese Loess Plateau with implications for the Quaternary climate change
3
2019
... 青藏高原东北缘作为高原向东北方向生长扩展的前缘部位,晚新生代以来构造活动显著;同时,地理位置独特,对区域气候和环境变化较为敏感.因此,该区是探讨青藏高原隆升过程和相关生态环境变化的理想地区
[20 -21 ] .兰州盆地作为陇中盆地的次级子盆地,沉积序列较为完整,近年来围绕上新统-更新统五泉砾岩和黄土地层学方面开展了大量工作,研究揭示上新世以来兰州地区经历了强烈的构造活动,同时也在皋兰山地区堆积了连续的上新世红黏土-第四纪黄土序列
[9 ,11 ] ,为探讨晚上新世-第四纪的构造隆升和古环境演变提供了理想材料.为系统厘清高原东北缘上新世-更新世转型期的古生态演化问题,同时也为弥补西部黄土高原地区上新世-更新世转型期长链正构烷烃碳同位素研究的不足,本文以皋兰山钻孔上新世-更新世转型期的红黏土-黄土岩芯(深度265.7~230.0 m)为研究对象(
图1 ),在已有黄土高精度磁性地层学基础上,详细开展了正构烷烃分布、总有机碳和长链正构烷烃碳同位素的有机地球化学分析,恢复了兰州地区上新世-更新世转型期(3.0~2.2 Ma)的C
3 /C
4 植被演化历史,以揭示高原东北缘兰州地区上新世-更新世过渡时期的古生态环境演变规律,进而尝试探讨该区晚上新世-早第四纪生态转型的可能原因.
图1 西部黄土高原皋兰山钻孔位置和季风示意图(改自Guo等<sup>[<xref ref-type="bibr" rid="R23">23</xref>]</sup>) Location of the Gaolanshan drill core on the Western Chinese Loess Plateau and patterns of associated monsoonal activities (Modified from Guo et al<sup>[<xref ref-type="bibr" rid="R23">23</xref>]</sup>) Fig.1 ![]()
1 研究区概况 兰州盆地作为青藏高原东北缘陇中盆地的次级子盆地,同时也位于黄土高原向青藏高原过渡的关键区域,从自然地理角度讲,兰州地处青藏高寒区、西北干旱区和东部季风区的交汇地带,亦是“季风三角”的枢纽部位所在(图1 ),对环境变化较为敏感,是进行古环境研究的理想区域.兰州现属温带半干旱气候,发育稀疏的灌木和草本植物;年均温10 ℃左右,年均降水约310 mm,主要集中在夏季的6—9月[22 ] . ...
... [
23 ])
Fig.1 ![]()
1 研究区概况 兰州盆地作为青藏高原东北缘陇中盆地的次级子盆地,同时也位于黄土高原向青藏高原过渡的关键区域,从自然地理角度讲,兰州地处青藏高寒区、西北干旱区和东部季风区的交汇地带,亦是“季风三角”的枢纽部位所在(图1 ),对环境变化较为敏感,是进行古环境研究的理想区域.兰州现属温带半干旱气候,发育稀疏的灌木和草本植物;年均温10 ℃左右,年均降水约310 mm,主要集中在夏季的6—9月[22 ] . ...
... 晚新生代青藏高原的隆升使兰州盆地新生代红层发生变形,其上覆盖厚度超过百余米的五泉砾岩[23 ] ;与此同时,在地形较为平坦的台地(皋兰山民族村)开始堆积风成红黏土-黄土序列[11 ] .兰州地区临近粉尘源区,且北支西风急流在高原北部绕流产生了兰州小高压[24 ] .因此,本区发育较厚的第四纪黄土序列,为区域古环境演化研究提供了高分辨率的理想材料. ...
冬季我国高原东侧切变线形成的分析
1
1963
... 晚新生代青藏高原的隆升使兰州盆地新生代红层发生变形,其上覆盖厚度超过百余米的五泉砾岩[23 ] ;与此同时,在地形较为平坦的台地(皋兰山民族村)开始堆积风成红黏土-黄土序列[11 ] .兰州地区临近粉尘源区,且北支西风急流在高原北部绕流产生了兰州小高压[24 ] .因此,本区发育较厚的第四纪黄土序列,为区域古环境演化研究提供了高分辨率的理想材料. ...
冬季我国高原东侧切变线形成的分析
1
1963
... 晚新生代青藏高原的隆升使兰州盆地新生代红层发生变形,其上覆盖厚度超过百余米的五泉砾岩[23 ] ;与此同时,在地形较为平坦的台地(皋兰山民族村)开始堆积风成红黏土-黄土序列[11 ] .兰州地区临近粉尘源区,且北支西风急流在高原北部绕流产生了兰州小高压[24 ] .因此,本区发育较厚的第四纪黄土序列,为区域古环境演化研究提供了高分辨率的理想材料. ...
Stacked 2.6-Ma grain size record from the Chinese loess based on five sections and correlation with the deep-sea δ 18 O record
1
2002
... 皋兰山钻孔(36°01′ N,103°51′ E,海拔2 120 m,图1 )位于兰州盆地南缘皋兰山民族村附近,共获取了黄土-红黏土岩芯样品265.7 m(取芯率高于95%),主要为棕黄色粉砂质黄土和褐红色古土壤互层.本文首先依据皋兰山钻孔高精度磁性地层年代选取绝对控制点[11 ] ,再结合黄土高原黄土-古土壤地层年代框架[25 ] ,利用线性内差法获得皋兰山钻孔底部的深度-年代模型.为探讨兰州盆地上新世-更新世转型期(3.0~2.2 Ma)C3 /C4 植被演化历史,本文选取265.7~230.0 m红黏土-黄土岩芯为研究对象,挑取70块原生样品进行生物标志化合物实验(间距0.5 m). ...
Purification of BaSO4 precipitate contaminated with organic matter for oxygen isotope measurements (δ 18 O and Δ 17 O)
1
2016
... 称取皋兰山岩芯样品约5 g,加过量盐酸之后再静置24 h,接着放入80 ℃水浴锅中加热,再加入过量盐酸并反应12 h,目的是充分反应,去除所有无机碳.然后将样品用蒸馏水洗至中性,烘干后用锡箔纸包好待测.整个实验测试分析在中国科学院广州地球化学研究所所完成,仪器类型为PYROcube型元素分析仪和Iso Prime 100型同位素质谱仪,测试方法同Xie等[26 ] . ...
青藏高原东北部湖泊沉积物末次冰消期以来正构烷烃分子分布特征及其碳、氢同位素的古环境意义
1
2011
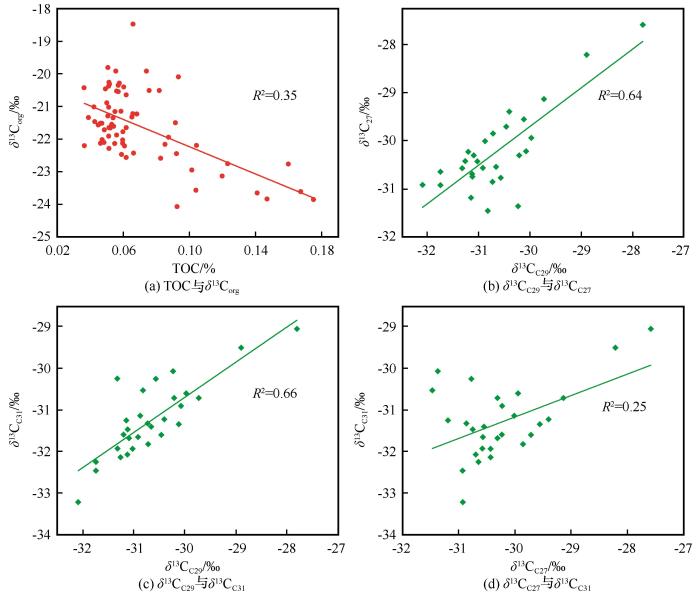
... 此外,皋兰山红黏土-黄土单体碳同位素值随碳链长度增加而偏负(图3 ),这与和其他地区的研究结果相似[27 -31 ] .具体而言,δ 13 CC27 数值在 -31.46‰~-27.59‰之间波动(均值为-30.21‰);δ 13 CC29 数值主要在-32.09‰~-27.80‰之间波动(均值为-30.65‰);δ 13 CC31 数值主要在-33.22‰~ -29.05‰之间波动(均值为-31.43‰).如图4 所示,δ 13 CC29 与δ 13 CC27 或δ 13 CC31 均呈现明显的正相关关系,而δ 13 CC27 与δ 13 CC31 线性关系较差,主要由于L33、S33和L34层位样品(图中左上部分)结果较为离散所致. ...
青藏高原东北部湖泊沉积物末次冰消期以来正构烷烃分子分布特征及其碳、氢同位素的古环境意义
1
2011
... 此外,皋兰山红黏土-黄土单体碳同位素值随碳链长度增加而偏负(图3 ),这与和其他地区的研究结果相似[27 -31 ] .具体而言,δ 13 CC27 数值在 -31.46‰~-27.59‰之间波动(均值为-30.21‰);δ 13 CC29 数值主要在-32.09‰~-27.80‰之间波动(均值为-30.65‰);δ 13 CC31 数值主要在-33.22‰~ -29.05‰之间波动(均值为-31.43‰).如图4 所示,δ 13 CC29 与δ 13 CC27 或δ 13 CC31 均呈现明显的正相关关系,而δ 13 CC27 与δ 13 CC31 线性关系较差,主要由于L33、S33和L34层位样品(图中左上部分)结果较为离散所致. ...
山西公海过去15 ka以来的湖泊沉积物长链正构烷烃单体碳/氢同位素及其古气候意义
0
2014
山西公海过去15 ka以来的湖泊沉积物长链正构烷烃单体碳/氢同位素及其古气候意义
0
2014
Sources of sedimentary lipids deduced from stable carbon-isotope analyses of individual compounds
0
1991
Stable carbon isotopic correlation of individual biolipids in aquatic organisms and a lake bottom sediment
0
1994
Molecular-isotopic stratigraphy of long-chain n -alkanes in Lake Baikal Holocene and glacial age sediments
1
2000
... 此外,皋兰山红黏土-黄土单体碳同位素值随碳链长度增加而偏负(图3 ),这与和其他地区的研究结果相似[27 -31 ] .具体而言,δ 13 CC27 数值在 -31.46‰~-27.59‰之间波动(均值为-30.21‰);δ 13 CC29 数值主要在-32.09‰~-27.80‰之间波动(均值为-30.65‰);δ 13 CC31 数值主要在-33.22‰~ -29.05‰之间波动(均值为-31.43‰).如图4 所示,δ 13 CC29 与δ 13 CC27 或δ 13 CC31 均呈现明显的正相关关系,而δ 13 CC27 与δ 13 CC31 线性关系较差,主要由于L33、S33和L34层位样品(图中左上部分)结果较为离散所致. ...
Development of Asian monsoon revealed by marked ecological shift during the latest Miocene in northern Pakistan
1
1989
... 一般认为,土壤有机质碳同位素的信号主要受控于原生植被的影响[32 ] ,指示了C3 /C4 植被演化过程.但与此同时,研究也发现微生物活动和外源输入等因素均会干扰碳同位素信号[33 -35 ] .因此,有机质的来源分析是进行古环境重建的基础和前提.风成黄土沉积的有机质可能来自当时的植被、成岩降解作用和外源输入.研究发现,我国西北地区表土的总有机碳同位素较上覆植被正偏约2.2‰,而外源输入带来的误差则更小,因此土壤有机碳同位素仍能较好指示C3 /C4 植被变化[34 ,36 ] .然而,在黄土高原东部,有机碳同位素和总有机质含量呈正相关;黄土层的δ 13 Corg 偏负,古土壤层的δ 13 Corg 偏正,指示了C3 /C4 植被相对含量的变化[37 -38 ] .但黄土高原西北部、新疆伊犁盆地和西欧莱茵河谷的黄土证据则显示:有机碳同位素和总有机碳负相关,黄土层偏正,古土壤层偏负,指示了纯C3 植被对气候变化的响应[39 -41 ] .这可能是因为这些地区不满足C4 植被的生长温度或降水阈值[42 -43 ] .此外,Liu等[44 ] 通过对西部黄土高原兰州和西宁末次冰期以来黄土有机碳同位素研究后发现,源区输入的外源信号可能掩盖了本地原生植被的信号,继而提出有机碳同位素难以准确反映西部黄土高原的古生态演变历史. ...
末次间冰期以来黄土高原的草原植被景观: 来自分子化石的证据
1
2002
... 一般认为,土壤有机质碳同位素的信号主要受控于原生植被的影响[32 ] ,指示了C3 /C4 植被演化过程.但与此同时,研究也发现微生物活动和外源输入等因素均会干扰碳同位素信号[33 -35 ] .因此,有机质的来源分析是进行古环境重建的基础和前提.风成黄土沉积的有机质可能来自当时的植被、成岩降解作用和外源输入.研究发现,我国西北地区表土的总有机碳同位素较上覆植被正偏约2.2‰,而外源输入带来的误差则更小,因此土壤有机碳同位素仍能较好指示C3 /C4 植被变化[34 ,36 ] .然而,在黄土高原东部,有机碳同位素和总有机质含量呈正相关;黄土层的δ 13 Corg 偏负,古土壤层的δ 13 Corg 偏正,指示了C3 /C4 植被相对含量的变化[37 -38 ] .但黄土高原西北部、新疆伊犁盆地和西欧莱茵河谷的黄土证据则显示:有机碳同位素和总有机碳负相关,黄土层偏正,古土壤层偏负,指示了纯C3 植被对气候变化的响应[39 -41 ] .这可能是因为这些地区不满足C4 植被的生长温度或降水阈值[42 -43 ] .此外,Liu等[44 ] 通过对西部黄土高原兰州和西宁末次冰期以来黄土有机碳同位素研究后发现,源区输入的外源信号可能掩盖了本地原生植被的信号,继而提出有机碳同位素难以准确反映西部黄土高原的古生态演变历史. ...
末次间冰期以来黄土高原的草原植被景观: 来自分子化石的证据
1
2002
... 一般认为,土壤有机质碳同位素的信号主要受控于原生植被的影响[32 ] ,指示了C3 /C4 植被演化过程.但与此同时,研究也发现微生物活动和外源输入等因素均会干扰碳同位素信号[33 -35 ] .因此,有机质的来源分析是进行古环境重建的基础和前提.风成黄土沉积的有机质可能来自当时的植被、成岩降解作用和外源输入.研究发现,我国西北地区表土的总有机碳同位素较上覆植被正偏约2.2‰,而外源输入带来的误差则更小,因此土壤有机碳同位素仍能较好指示C3 /C4 植被变化[34 ,36 ] .然而,在黄土高原东部,有机碳同位素和总有机质含量呈正相关;黄土层的δ 13 Corg 偏负,古土壤层的δ 13 Corg 偏正,指示了C3 /C4 植被相对含量的变化[37 -38 ] .但黄土高原西北部、新疆伊犁盆地和西欧莱茵河谷的黄土证据则显示:有机碳同位素和总有机碳负相关,黄土层偏正,古土壤层偏负,指示了纯C3 植被对气候变化的响应[39 -41 ] .这可能是因为这些地区不满足C4 植被的生长温度或降水阈值[42 -43 ] .此外,Liu等[44 ] 通过对西部黄土高原兰州和西宁末次冰期以来黄土有机碳同位素研究后发现,源区输入的外源信号可能掩盖了本地原生植被的信号,继而提出有机碳同位素难以准确反映西部黄土高原的古生态演变历史. ...
黄土高原西部地区黄土地层有机质主要来源分析
1
2015
... 一般认为,土壤有机质碳同位素的信号主要受控于原生植被的影响[32 ] ,指示了C3 /C4 植被演化过程.但与此同时,研究也发现微生物活动和外源输入等因素均会干扰碳同位素信号[33 -35 ] .因此,有机质的来源分析是进行古环境重建的基础和前提.风成黄土沉积的有机质可能来自当时的植被、成岩降解作用和外源输入.研究发现,我国西北地区表土的总有机碳同位素较上覆植被正偏约2.2‰,而外源输入带来的误差则更小,因此土壤有机碳同位素仍能较好指示C3 /C4 植被变化[34 ,36 ] .然而,在黄土高原东部,有机碳同位素和总有机质含量呈正相关;黄土层的δ 13 Corg 偏负,古土壤层的δ 13 Corg 偏正,指示了C3 /C4 植被相对含量的变化[37 -38 ] .但黄土高原西北部、新疆伊犁盆地和西欧莱茵河谷的黄土证据则显示:有机碳同位素和总有机碳负相关,黄土层偏正,古土壤层偏负,指示了纯C3 植被对气候变化的响应[39 -41 ] .这可能是因为这些地区不满足C4 植被的生长温度或降水阈值[42 -43 ] .此外,Liu等[44 ] 通过对西部黄土高原兰州和西宁末次冰期以来黄土有机碳同位素研究后发现,源区输入的外源信号可能掩盖了本地原生植被的信号,继而提出有机碳同位素难以准确反映西部黄土高原的古生态演变历史. ...
黄土高原西部地区黄土地层有机质主要来源分析
1
2015
... 一般认为,土壤有机质碳同位素的信号主要受控于原生植被的影响[32 ] ,指示了C3 /C4 植被演化过程.但与此同时,研究也发现微生物活动和外源输入等因素均会干扰碳同位素信号[33 -35 ] .因此,有机质的来源分析是进行古环境重建的基础和前提.风成黄土沉积的有机质可能来自当时的植被、成岩降解作用和外源输入.研究发现,我国西北地区表土的总有机碳同位素较上覆植被正偏约2.2‰,而外源输入带来的误差则更小,因此土壤有机碳同位素仍能较好指示C3 /C4 植被变化[34 ,36 ] .然而,在黄土高原东部,有机碳同位素和总有机质含量呈正相关;黄土层的δ 13 Corg 偏负,古土壤层的δ 13 Corg 偏正,指示了C3 /C4 植被相对含量的变化[37 -38 ] .但黄土高原西北部、新疆伊犁盆地和西欧莱茵河谷的黄土证据则显示:有机碳同位素和总有机碳负相关,黄土层偏正,古土壤层偏负,指示了纯C3 植被对气候变化的响应[39 -41 ] .这可能是因为这些地区不满足C4 植被的生长温度或降水阈值[42 -43 ] .此外,Liu等[44 ] 通过对西部黄土高原兰州和西宁末次冰期以来黄土有机碳同位素研究后发现,源区输入的外源信号可能掩盖了本地原生植被的信号,继而提出有机碳同位素难以准确反映西部黄土高原的古生态演变历史. ...
Carbon and nitrogen dynamics along the decay continuum: plant litter to soil organic matter
1
1989
... 一般认为,土壤有机质碳同位素的信号主要受控于原生植被的影响[32 ] ,指示了C3 /C4 植被演化过程.但与此同时,研究也发现微生物活动和外源输入等因素均会干扰碳同位素信号[33 -35 ] .因此,有机质的来源分析是进行古环境重建的基础和前提.风成黄土沉积的有机质可能来自当时的植被、成岩降解作用和外源输入.研究发现,我国西北地区表土的总有机碳同位素较上覆植被正偏约2.2‰,而外源输入带来的误差则更小,因此土壤有机碳同位素仍能较好指示C3 /C4 植被变化[34 ,36 ] .然而,在黄土高原东部,有机碳同位素和总有机质含量呈正相关;黄土层的δ 13 Corg 偏负,古土壤层的δ 13 Corg 偏正,指示了C3 /C4 植被相对含量的变化[37 -38 ] .但黄土高原西北部、新疆伊犁盆地和西欧莱茵河谷的黄土证据则显示:有机碳同位素和总有机碳负相关,黄土层偏正,古土壤层偏负,指示了纯C3 植被对气候变化的响应[39 -41 ] .这可能是因为这些地区不满足C4 植被的生长温度或降水阈值[42 -43 ] .此外,Liu等[44 ] 通过对西部黄土高原兰州和西宁末次冰期以来黄土有机碳同位素研究后发现,源区输入的外源信号可能掩盖了本地原生植被的信号,继而提出有机碳同位素难以准确反映西部黄土高原的古生态演变历史. ...
稳定碳同位素在第四纪古环境研究中的应用
1
2003
... 一般认为,土壤有机质碳同位素的信号主要受控于原生植被的影响[32 ] ,指示了C3 /C4 植被演化过程.但与此同时,研究也发现微生物活动和外源输入等因素均会干扰碳同位素信号[33 -35 ] .因此,有机质的来源分析是进行古环境重建的基础和前提.风成黄土沉积的有机质可能来自当时的植被、成岩降解作用和外源输入.研究发现,我国西北地区表土的总有机碳同位素较上覆植被正偏约2.2‰,而外源输入带来的误差则更小,因此土壤有机碳同位素仍能较好指示C3 /C4 植被变化[34 ,36 ] .然而,在黄土高原东部,有机碳同位素和总有机质含量呈正相关;黄土层的δ 13 Corg 偏负,古土壤层的δ 13 Corg 偏正,指示了C3 /C4 植被相对含量的变化[37 -38 ] .但黄土高原西北部、新疆伊犁盆地和西欧莱茵河谷的黄土证据则显示:有机碳同位素和总有机碳负相关,黄土层偏正,古土壤层偏负,指示了纯C3 植被对气候变化的响应[39 -41 ] .这可能是因为这些地区不满足C4 植被的生长温度或降水阈值[42 -43 ] .此外,Liu等[44 ] 通过对西部黄土高原兰州和西宁末次冰期以来黄土有机碳同位素研究后发现,源区输入的外源信号可能掩盖了本地原生植被的信号,继而提出有机碳同位素难以准确反映西部黄土高原的古生态演变历史. ...
稳定碳同位素在第四纪古环境研究中的应用
1
2003
... 一般认为,土壤有机质碳同位素的信号主要受控于原生植被的影响[32 ] ,指示了C3 /C4 植被演化过程.但与此同时,研究也发现微生物活动和外源输入等因素均会干扰碳同位素信号[33 -35 ] .因此,有机质的来源分析是进行古环境重建的基础和前提.风成黄土沉积的有机质可能来自当时的植被、成岩降解作用和外源输入.研究发现,我国西北地区表土的总有机碳同位素较上覆植被正偏约2.2‰,而外源输入带来的误差则更小,因此土壤有机碳同位素仍能较好指示C3 /C4 植被变化[34 ,36 ] .然而,在黄土高原东部,有机碳同位素和总有机质含量呈正相关;黄土层的δ 13 Corg 偏负,古土壤层的δ 13 Corg 偏正,指示了C3 /C4 植被相对含量的变化[37 -38 ] .但黄土高原西北部、新疆伊犁盆地和西欧莱茵河谷的黄土证据则显示:有机碳同位素和总有机碳负相关,黄土层偏正,古土壤层偏负,指示了纯C3 植被对气候变化的响应[39 -41 ] .这可能是因为这些地区不满足C4 植被的生长温度或降水阈值[42 -43 ] .此外,Liu等[44 ] 通过对西部黄土高原兰州和西宁末次冰期以来黄土有机碳同位素研究后发现,源区输入的外源信号可能掩盖了本地原生植被的信号,继而提出有机碳同位素难以准确反映西部黄土高原的古生态演变历史. ...
Lower temperature as the main cause of C4 plant declines during the glacial periods on the Chinese Loess Plateau
1
2003
... 一般认为,土壤有机质碳同位素的信号主要受控于原生植被的影响[32 ] ,指示了C3 /C4 植被演化过程.但与此同时,研究也发现微生物活动和外源输入等因素均会干扰碳同位素信号[33 -35 ] .因此,有机质的来源分析是进行古环境重建的基础和前提.风成黄土沉积的有机质可能来自当时的植被、成岩降解作用和外源输入.研究发现,我国西北地区表土的总有机碳同位素较上覆植被正偏约2.2‰,而外源输入带来的误差则更小,因此土壤有机碳同位素仍能较好指示C3 /C4 植被变化[34 ,36 ] .然而,在黄土高原东部,有机碳同位素和总有机质含量呈正相关;黄土层的δ 13 Corg 偏负,古土壤层的δ 13 Corg 偏正,指示了C3 /C4 植被相对含量的变化[37 -38 ] .但黄土高原西北部、新疆伊犁盆地和西欧莱茵河谷的黄土证据则显示:有机碳同位素和总有机碳负相关,黄土层偏正,古土壤层偏负,指示了纯C3 植被对气候变化的响应[39 -41 ] .这可能是因为这些地区不满足C4 植被的生长温度或降水阈值[42 -43 ] .此外,Liu等[44 ] 通过对西部黄土高原兰州和西宁末次冰期以来黄土有机碳同位素研究后发现,源区输入的外源信号可能掩盖了本地原生植被的信号,继而提出有机碳同位素难以准确反映西部黄土高原的古生态演变历史. ...
Summer monsoon intensity controls C4 /C3 plant abundance during the last 35 ka in the Chinese Loess Plateau: Carbon isotope evidence from bulk organic matter and individual leaf waxes
1
2005
... 一般认为,土壤有机质碳同位素的信号主要受控于原生植被的影响[32 ] ,指示了C3 /C4 植被演化过程.但与此同时,研究也发现微生物活动和外源输入等因素均会干扰碳同位素信号[33 -35 ] .因此,有机质的来源分析是进行古环境重建的基础和前提.风成黄土沉积的有机质可能来自当时的植被、成岩降解作用和外源输入.研究发现,我国西北地区表土的总有机碳同位素较上覆植被正偏约2.2‰,而外源输入带来的误差则更小,因此土壤有机碳同位素仍能较好指示C3 /C4 植被变化[34 ,36 ] .然而,在黄土高原东部,有机碳同位素和总有机质含量呈正相关;黄土层的δ 13 Corg 偏负,古土壤层的δ 13 Corg 偏正,指示了C3 /C4 植被相对含量的变化[37 -38 ] .但黄土高原西北部、新疆伊犁盆地和西欧莱茵河谷的黄土证据则显示:有机碳同位素和总有机碳负相关,黄土层偏正,古土壤层偏负,指示了纯C3 植被对气候变化的响应[39 -41 ] .这可能是因为这些地区不满足C4 植被的生长温度或降水阈值[42 -43 ] .此外,Liu等[44 ] 通过对西部黄土高原兰州和西宁末次冰期以来黄土有机碳同位素研究后发现,源区输入的外源信号可能掩盖了本地原生植被的信号,继而提出有机碳同位素难以准确反映西部黄土高原的古生态演变历史. ...
黄土高原西部地区末次冰期和全新世有机碳同位素变化与C3 /C4 植被类型转换研究
1
2005
... 一般认为,土壤有机质碳同位素的信号主要受控于原生植被的影响[32 ] ,指示了C3 /C4 植被演化过程.但与此同时,研究也发现微生物活动和外源输入等因素均会干扰碳同位素信号[33 -35 ] .因此,有机质的来源分析是进行古环境重建的基础和前提.风成黄土沉积的有机质可能来自当时的植被、成岩降解作用和外源输入.研究发现,我国西北地区表土的总有机碳同位素较上覆植被正偏约2.2‰,而外源输入带来的误差则更小,因此土壤有机碳同位素仍能较好指示C3 /C4 植被变化[34 ,36 ] .然而,在黄土高原东部,有机碳同位素和总有机质含量呈正相关;黄土层的δ 13 Corg 偏负,古土壤层的δ 13 Corg 偏正,指示了C3 /C4 植被相对含量的变化[37 -38 ] .但黄土高原西北部、新疆伊犁盆地和西欧莱茵河谷的黄土证据则显示:有机碳同位素和总有机碳负相关,黄土层偏正,古土壤层偏负,指示了纯C3 植被对气候变化的响应[39 -41 ] .这可能是因为这些地区不满足C4 植被的生长温度或降水阈值[42 -43 ] .此外,Liu等[44 ] 通过对西部黄土高原兰州和西宁末次冰期以来黄土有机碳同位素研究后发现,源区输入的外源信号可能掩盖了本地原生植被的信号,继而提出有机碳同位素难以准确反映西部黄土高原的古生态演变历史. ...
黄土高原西部地区末次冰期和全新世有机碳同位素变化与C3 /C4 植被类型转换研究
1
2005
... 一般认为,土壤有机质碳同位素的信号主要受控于原生植被的影响[32 ] ,指示了C3 /C4 植被演化过程.但与此同时,研究也发现微生物活动和外源输入等因素均会干扰碳同位素信号[33 -35 ] .因此,有机质的来源分析是进行古环境重建的基础和前提.风成黄土沉积的有机质可能来自当时的植被、成岩降解作用和外源输入.研究发现,我国西北地区表土的总有机碳同位素较上覆植被正偏约2.2‰,而外源输入带来的误差则更小,因此土壤有机碳同位素仍能较好指示C3 /C4 植被变化[34 ,36 ] .然而,在黄土高原东部,有机碳同位素和总有机质含量呈正相关;黄土层的δ 13 Corg 偏负,古土壤层的δ 13 Corg 偏正,指示了C3 /C4 植被相对含量的变化[37 -38 ] .但黄土高原西北部、新疆伊犁盆地和西欧莱茵河谷的黄土证据则显示:有机碳同位素和总有机碳负相关,黄土层偏正,古土壤层偏负,指示了纯C3 植被对气候变化的响应[39 -41 ] .这可能是因为这些地区不满足C4 植被的生长温度或降水阈值[42 -43 ] .此外,Liu等[44 ] 通过对西部黄土高原兰州和西宁末次冰期以来黄土有机碳同位素研究后发现,源区输入的外源信号可能掩盖了本地原生植被的信号,继而提出有机碳同位素难以准确反映西部黄土高原的古生态演变历史. ...
陇西黄土高原和新疆伊犁盆地黄土有机碳同位素的变化及其古环境意义
0
2013
陇西黄土高原和新疆伊犁盆地黄土有机碳同位素的变化及其古环境意义
0
2013
δ 13 C variations of loess organic matter as a record of the vegetation response to climatic changes during the Weichselian
1
1998
... 一般认为,土壤有机质碳同位素的信号主要受控于原生植被的影响[32 ] ,指示了C3 /C4 植被演化过程.但与此同时,研究也发现微生物活动和外源输入等因素均会干扰碳同位素信号[33 -35 ] .因此,有机质的来源分析是进行古环境重建的基础和前提.风成黄土沉积的有机质可能来自当时的植被、成岩降解作用和外源输入.研究发现,我国西北地区表土的总有机碳同位素较上覆植被正偏约2.2‰,而外源输入带来的误差则更小,因此土壤有机碳同位素仍能较好指示C3 /C4 植被变化[34 ,36 ] .然而,在黄土高原东部,有机碳同位素和总有机质含量呈正相关;黄土层的δ 13 Corg 偏负,古土壤层的δ 13 Corg 偏正,指示了C3 /C4 植被相对含量的变化[37 -38 ] .但黄土高原西北部、新疆伊犁盆地和西欧莱茵河谷的黄土证据则显示:有机碳同位素和总有机碳负相关,黄土层偏正,古土壤层偏负,指示了纯C3 植被对气候变化的响应[39 -41 ] .这可能是因为这些地区不满足C4 植被的生长温度或降水阈值[42 -43 ] .此外,Liu等[44 ] 通过对西部黄土高原兰州和西宁末次冰期以来黄土有机碳同位素研究后发现,源区输入的外源信号可能掩盖了本地原生植被的信号,继而提出有机碳同位素难以准确反映西部黄土高原的古生态演变历史. ...
The precipitation “threshold value” on C4 /C3 abundance of the Loess Plateau, China
1
2015
... 一般认为,土壤有机质碳同位素的信号主要受控于原生植被的影响[32 ] ,指示了C3 /C4 植被演化过程.但与此同时,研究也发现微生物活动和外源输入等因素均会干扰碳同位素信号[33 -35 ] .因此,有机质的来源分析是进行古环境重建的基础和前提.风成黄土沉积的有机质可能来自当时的植被、成岩降解作用和外源输入.研究发现,我国西北地区表土的总有机碳同位素较上覆植被正偏约2.2‰,而外源输入带来的误差则更小,因此土壤有机碳同位素仍能较好指示C3 /C4 植被变化[34 ,36 ] .然而,在黄土高原东部,有机碳同位素和总有机质含量呈正相关;黄土层的δ 13 Corg 偏负,古土壤层的δ 13 Corg 偏正,指示了C3 /C4 植被相对含量的变化[37 -38 ] .但黄土高原西北部、新疆伊犁盆地和西欧莱茵河谷的黄土证据则显示:有机碳同位素和总有机碳负相关,黄土层偏正,古土壤层偏负,指示了纯C3 植被对气候变化的响应[39 -41 ] .这可能是因为这些地区不满足C4 植被的生长温度或降水阈值[42 -43 ] .此外,Liu等[44 ] 通过对西部黄土高原兰州和西宁末次冰期以来黄土有机碳同位素研究后发现,源区输入的外源信号可能掩盖了本地原生植被的信号,继而提出有机碳同位素难以准确反映西部黄土高原的古生态演变历史. ...
Variation and paleoclimatic significance of organic carbon isotopes of Ili loess in arid Central Asia
1
2013
... 一般认为,土壤有机质碳同位素的信号主要受控于原生植被的影响[32 ] ,指示了C3 /C4 植被演化过程.但与此同时,研究也发现微生物活动和外源输入等因素均会干扰碳同位素信号[33 -35 ] .因此,有机质的来源分析是进行古环境重建的基础和前提.风成黄土沉积的有机质可能来自当时的植被、成岩降解作用和外源输入.研究发现,我国西北地区表土的总有机碳同位素较上覆植被正偏约2.2‰,而外源输入带来的误差则更小,因此土壤有机碳同位素仍能较好指示C3 /C4 植被变化[34 ,36 ] .然而,在黄土高原东部,有机碳同位素和总有机质含量呈正相关;黄土层的δ 13 Corg 偏负,古土壤层的δ 13 Corg 偏正,指示了C3 /C4 植被相对含量的变化[37 -38 ] .但黄土高原西北部、新疆伊犁盆地和西欧莱茵河谷的黄土证据则显示:有机碳同位素和总有机碳负相关,黄土层偏正,古土壤层偏负,指示了纯C3 植被对气候变化的响应[39 -41 ] .这可能是因为这些地区不满足C4 植被的生长温度或降水阈值[42 -43 ] .此外,Liu等[44 ] 通过对西部黄土高原兰州和西宁末次冰期以来黄土有机碳同位素研究后发现,源区输入的外源信号可能掩盖了本地原生植被的信号,继而提出有机碳同位素难以准确反映西部黄土高原的古生态演变历史. ...
Contribution of inherent organic carbon to the bulk δ 13 C signal in loess deposits from the arid western Chinese Loess Plateau
1
2007
... 一般认为,土壤有机质碳同位素的信号主要受控于原生植被的影响[32 ] ,指示了C3 /C4 植被演化过程.但与此同时,研究也发现微生物活动和外源输入等因素均会干扰碳同位素信号[33 -35 ] .因此,有机质的来源分析是进行古环境重建的基础和前提.风成黄土沉积的有机质可能来自当时的植被、成岩降解作用和外源输入.研究发现,我国西北地区表土的总有机碳同位素较上覆植被正偏约2.2‰,而外源输入带来的误差则更小,因此土壤有机碳同位素仍能较好指示C3 /C4 植被变化[34 ,36 ] .然而,在黄土高原东部,有机碳同位素和总有机质含量呈正相关;黄土层的δ 13 Corg 偏负,古土壤层的δ 13 Corg 偏正,指示了C3 /C4 植被相对含量的变化[37 -38 ] .但黄土高原西北部、新疆伊犁盆地和西欧莱茵河谷的黄土证据则显示:有机碳同位素和总有机碳负相关,黄土层偏正,古土壤层偏负,指示了纯C3 植被对气候变化的响应[39 -41 ] .这可能是因为这些地区不满足C4 植被的生长温度或降水阈值[42 -43 ] .此外,Liu等[44 ] 通过对西部黄土高原兰州和西宁末次冰期以来黄土有机碳同位素研究后发现,源区输入的外源信号可能掩盖了本地原生植被的信号,继而提出有机碳同位素难以准确反映西部黄土高原的古生态演变历史. ...
Characterization of unresolved complex mixtures of hydrocarbons in petroleum
1
1990
... 如前所述,典型黄土层L32和L33的多数样品正构烷烃呈双鼓包分布(图2 ),表明曾遭受明显的微生物降解[45 -46 ] .而黄土地层和粒度等证据也表明:L32和L33时期,源区和沉积区气候寒冷干燥,均不利于正构烷烃降解;黄土层粒径整体较粗,显示冬季风较为强盛[11 ] .同时,黄土堆积区气候干旱,不利于植被发育.由此推测,降解产物UCM主要为源区母岩输入贡献.事实上,TOC和δ 13 Corg 指标最呈规律性变化的层位正是正构烷烃表现出外源输入特征的L32和L33黄土层(图3 ).这直接支持皋兰山底部黄土层有机碳受外源输入影响的看法.此外,该时段磁化率也表现出随粒径变粗而增大趋势[47 ] .这一证据亦支持源区磁性矿物输入增多的观点.因此,皋兰山样品的TOC高值和δ 13 Corg 的偏负很可能是外源输入导致的假象. ...
泌阳凹陷生物降解油"基线鼓包"成因及化合物组成
1
2014
... 如前所述,典型黄土层L32和L33的多数样品正构烷烃呈双鼓包分布(图2 ),表明曾遭受明显的微生物降解[45 -46 ] .而黄土地层和粒度等证据也表明:L32和L33时期,源区和沉积区气候寒冷干燥,均不利于正构烷烃降解;黄土层粒径整体较粗,显示冬季风较为强盛[11 ] .同时,黄土堆积区气候干旱,不利于植被发育.由此推测,降解产物UCM主要为源区母岩输入贡献.事实上,TOC和δ 13 Corg 指标最呈规律性变化的层位正是正构烷烃表现出外源输入特征的L32和L33黄土层(图3 ).这直接支持皋兰山底部黄土层有机碳受外源输入影响的看法.此外,该时段磁化率也表现出随粒径变粗而增大趋势[47 ] .这一证据亦支持源区磁性矿物输入增多的观点.因此,皋兰山样品的TOC高值和δ 13 Corg 的偏负很可能是外源输入导致的假象. ...
泌阳凹陷生物降解油"基线鼓包"成因及化合物组成
1
2014
... 如前所述,典型黄土层L32和L33的多数样品正构烷烃呈双鼓包分布(图2 ),表明曾遭受明显的微生物降解[45 -46 ] .而黄土地层和粒度等证据也表明:L32和L33时期,源区和沉积区气候寒冷干燥,均不利于正构烷烃降解;黄土层粒径整体较粗,显示冬季风较为强盛[11 ] .同时,黄土堆积区气候干旱,不利于植被发育.由此推测,降解产物UCM主要为源区母岩输入贡献.事实上,TOC和δ 13 Corg 指标最呈规律性变化的层位正是正构烷烃表现出外源输入特征的L32和L33黄土层(图3 ).这直接支持皋兰山底部黄土层有机碳受外源输入影响的看法.此外,该时段磁化率也表现出随粒径变粗而增大趋势[47 ] .这一证据亦支持源区磁性矿物输入增多的观点.因此,皋兰山样品的TOC高值和δ 13 Corg 的偏负很可能是外源输入导致的假象. ...
兰州皋兰山黄土地层划分及其记录的气候演变
3
2017
... 如前所述,典型黄土层L32和L33的多数样品正构烷烃呈双鼓包分布(图2 ),表明曾遭受明显的微生物降解[45 -46 ] .而黄土地层和粒度等证据也表明:L32和L33时期,源区和沉积区气候寒冷干燥,均不利于正构烷烃降解;黄土层粒径整体较粗,显示冬季风较为强盛[11 ] .同时,黄土堆积区气候干旱,不利于植被发育.由此推测,降解产物UCM主要为源区母岩输入贡献.事实上,TOC和δ 13 Corg 指标最呈规律性变化的层位正是正构烷烃表现出外源输入特征的L32和L33黄土层(图3 ).这直接支持皋兰山底部黄土层有机碳受外源输入影响的看法.此外,该时段磁化率也表现出随粒径变粗而增大趋势[47 ] .这一证据亦支持源区磁性矿物输入增多的观点.因此,皋兰山样品的TOC高值和δ 13 Corg 的偏负很可能是外源输入导致的假象. ...
... 第一阶段:3.0~2.9 Ma时段,主体为红黏土沉积,C4 植被相对比例最高.上新世大暖期结束时(3.0 Ma),δ 13 Cn -alkane 整体较为偏正[图(3)和(5)],远高于C3 植被δ 13 Cn -alkane 分布范围(-39‰~-32‰).ACL值较低,指示气候整体温暖,但ACL逐渐增大,支持气候逐渐变冷.与此同时,ACL与δ 13 Cn -alkane 呈反相位变化,即气候越冷干,δ 13 Cn -alkane 值逐渐偏负,与黄土高原东部的C4 模式一致.据此推断:此时气候条件仍相对温暖湿润,适宜C4 植被发育,δ 13 Cn -alkane 由C4 植被相对丰度变化主导.定量重建结果表明,此时段C4 植被比例最高可达40%(图5 ).2.9~2.8 Ma,ACL值逐渐降低,磁化率较之前明显增加[47 ] ,表明此时降水较之前有所增多,相对充沛.而兰州位于季风影响的边缘区域,雨热同期主要集中在生长季(夏季).因此,强降水也意味着季节性加强,更适宜C4 生长.但此时C4 植被仍稳定在低水平,说明降水不是C4 植被减少的主要因素.同时,此时段p CO2 略微降低[52 ,55 ] (图5 ),也更适宜C4 发育.排除前述因子后,C4 植被生长的限制因素极可能为温度.研究表明,上新世温暖期(3.3~3.0 Ma)结束后,氧同位素揭示的全球冰量显著扩张[56 ] ,温度进一步下降.这可能逐渐突破了C4 植被生长阈值,进而导致兰州皋兰山C4 植被快速减少. ...
... 2.8 Ma左右,沉积相从红黏土转变为黄土,北半球冰盖开始扩张,ACL和δ 13 Cn -alkane 波动明显,黄土中值粒径增加指示冬季风加强[11 ] ,气候转冷干.值得注意的是,δ 13 CC27 与δ 13 CC31 发生了截然不同的变化(图3 ).这是C3 模式和C4 模式都难以解释的.据此本文认为,可能是因为皋兰山C3 植被与C4 植被的主峰碳数不同,而较为贫乏的C4 植被不足以掩盖C3 植被δ 13 Cn -alkane 对气候响应的信号,从而导致了这种分异.δ 13 CC27 与δ 13 CC31 反向变化一直持续到2.6 Ma前后,标志着此时段气候较之前更为恶化.与第一阶段类似的是:S33阶段高磁化率[47 ] 的丰沛降水也未能显著影响C4 含量,表明降水量亦不是C4 发育的控制因子.因此,此时期C4 植被减少的主要因素仍是温度. ...
兰州皋兰山黄土地层划分及其记录的气候演变
3
2017
... 如前所述,典型黄土层L32和L33的多数样品正构烷烃呈双鼓包分布(图2 ),表明曾遭受明显的微生物降解[45 -46 ] .而黄土地层和粒度等证据也表明:L32和L33时期,源区和沉积区气候寒冷干燥,均不利于正构烷烃降解;黄土层粒径整体较粗,显示冬季风较为强盛[11 ] .同时,黄土堆积区气候干旱,不利于植被发育.由此推测,降解产物UCM主要为源区母岩输入贡献.事实上,TOC和δ 13 Corg 指标最呈规律性变化的层位正是正构烷烃表现出外源输入特征的L32和L33黄土层(图3 ).这直接支持皋兰山底部黄土层有机碳受外源输入影响的看法.此外,该时段磁化率也表现出随粒径变粗而增大趋势[47 ] .这一证据亦支持源区磁性矿物输入增多的观点.因此,皋兰山样品的TOC高值和δ 13 Corg 的偏负很可能是外源输入导致的假象. ...
... 第一阶段:3.0~2.9 Ma时段,主体为红黏土沉积,C4 植被相对比例最高.上新世大暖期结束时(3.0 Ma),δ 13 Cn -alkane 整体较为偏正[图(3)和(5)],远高于C3 植被δ 13 Cn -alkane 分布范围(-39‰~-32‰).ACL值较低,指示气候整体温暖,但ACL逐渐增大,支持气候逐渐变冷.与此同时,ACL与δ 13 Cn -alkane 呈反相位变化,即气候越冷干,δ 13 Cn -alkane 值逐渐偏负,与黄土高原东部的C4 模式一致.据此推断:此时气候条件仍相对温暖湿润,适宜C4 植被发育,δ 13 Cn -alkane 由C4 植被相对丰度变化主导.定量重建结果表明,此时段C4 植被比例最高可达40%(图5 ).2.9~2.8 Ma,ACL值逐渐降低,磁化率较之前明显增加[47 ] ,表明此时降水较之前有所增多,相对充沛.而兰州位于季风影响的边缘区域,雨热同期主要集中在生长季(夏季).因此,强降水也意味着季节性加强,更适宜C4 生长.但此时C4 植被仍稳定在低水平,说明降水不是C4 植被减少的主要因素.同时,此时段p CO2 略微降低[52 ,55 ] (图5 ),也更适宜C4 发育.排除前述因子后,C4 植被生长的限制因素极可能为温度.研究表明,上新世温暖期(3.3~3.0 Ma)结束后,氧同位素揭示的全球冰量显著扩张[56 ] ,温度进一步下降.这可能逐渐突破了C4 植被生长阈值,进而导致兰州皋兰山C4 植被快速减少. ...
... 2.8 Ma左右,沉积相从红黏土转变为黄土,北半球冰盖开始扩张,ACL和δ 13 Cn -alkane 波动明显,黄土中值粒径增加指示冬季风加强[11 ] ,气候转冷干.值得注意的是,δ 13 CC27 与δ 13 CC31 发生了截然不同的变化(图3 ).这是C3 模式和C4 模式都难以解释的.据此本文认为,可能是因为皋兰山C3 植被与C4 植被的主峰碳数不同,而较为贫乏的C4 植被不足以掩盖C3 植被δ 13 Cn -alkane 对气候响应的信号,从而导致了这种分异.δ 13 CC27 与δ 13 CC31 反向变化一直持续到2.6 Ma前后,标志着此时段气候较之前更为恶化.与第一阶段类似的是:S33阶段高磁化率[47 ] 的丰沛降水也未能显著影响C4 含量,表明降水量亦不是C4 发育的控制因子.因此,此时期C4 植被减少的主要因素仍是温度. ...
Source identification of oil spills based on the isotopic composition of individual components in weathered oil samples
1
1997
... 更重要的是,皋兰山红黏土-黄土总有机碳同位素δ 13 Corg 和长链正构烷烃单体碳同位素δ 13 Cn -alkane 变化不一致(图3 ).长链正构烷烃直接来源于高等陆生植被,沉积后性质比较稳定,并且其碳同位素值在风化、生物降解作用下无明显变化[48 -49 ] .因此,δ 13 Cn -alkane 相较于δ 13 Corg 具有来源明确、性质稳定的优点,被广泛应用于C3 /C4 植被重建工作[50 ] .在红黏土、L34、S33和L33地层转变处,δ 13 Corg 均表现出明显变化;但这些变化却未被δ 13 Cn -alkane 所记录.同时,相较于S32古土壤层中δ 13 Cn -alkane 的正偏,δ 13 Corg 则表现为无趋势的波动.上述矛盾充分说明,δ 13 Corg 受到外源输入因素的影响. ...
Absence of stable carbon isotope fractionation of saturated and polycyclic aromatic hydrocarbons during aerobic bacterial biodegradation
1
2002
... 更重要的是,皋兰山红黏土-黄土总有机碳同位素δ 13 Corg 和长链正构烷烃单体碳同位素δ 13 Cn -alkane 变化不一致(图3 ).长链正构烷烃直接来源于高等陆生植被,沉积后性质比较稳定,并且其碳同位素值在风化、生物降解作用下无明显变化[48 -49 ] .因此,δ 13 Cn -alkane 相较于δ 13 Corg 具有来源明确、性质稳定的优点,被广泛应用于C3 /C4 植被重建工作[50 ] .在红黏土、L34、S33和L33地层转变处,δ 13 Corg 均表现出明显变化;但这些变化却未被δ 13 Cn -alkane 所记录.同时,相较于S32古土壤层中δ 13 Cn -alkane 的正偏,δ 13 Corg 则表现为无趋势的波动.上述矛盾充分说明,δ 13 Corg 受到外源输入因素的影响. ...
Northeast African vegetation change over 12 my
2
2013
... 更重要的是,皋兰山红黏土-黄土总有机碳同位素δ 13 Corg 和长链正构烷烃单体碳同位素δ 13 Cn -alkane 变化不一致(图3 ).长链正构烷烃直接来源于高等陆生植被,沉积后性质比较稳定,并且其碳同位素值在风化、生物降解作用下无明显变化[48 -49 ] .因此,δ 13 Cn -alkane 相较于δ 13 Corg 具有来源明确、性质稳定的优点,被广泛应用于C3 /C4 植被重建工作[50 ] .在红黏土、L34、S33和L33地层转变处,δ 13 Corg 均表现出明显变化;但这些变化却未被δ 13 Cn -alkane 所记录.同时,相较于S32古土壤层中δ 13 Cn -alkane 的正偏,δ 13 Corg 则表现为无趋势的波动.上述矛盾充分说明,δ 13 Corg 受到外源输入因素的影响. ...
... C4 植被的出现可以追溯到32~30 Ma,其大幅扩张一般认为发生在晚中新世[57 ] .来自巴基斯坦Siwalik沉积有机质和无机碳[58 ] 、北美哺乳动物牙釉化石[59 ] 、南美东部钻孔无机碳和动物化石[60 ] 和东非钻孔中动物化石[61 ] 等研究结果均支持这一说法:晚中新世7~8 Ma左右全球发生C4 植被的显著扩张.因此,C4 植被扩张是由大气CO2 浓度降低驱动的全球生态事件[62 ] .21世纪以来,越来越多的记录表明,全球C4 植被扩张的时间和范围并不一致:非洲东北部正构烷烃δ 13 C表明,C4 植被在11 Ma业已达到较高水平[50 ] ;新疆塔里木盆地钻孔有机碳δ 13 C显示,C4 植被在5.3 Ma显著增加[63 ] .这也得到日本海钻孔正构烷烃δ 13 C和内蒙古哺乳动物牙釉化石δ 13 C结果的支持[14 ,64 ] .近来,Lu等[15 ] 汇总全球记录后提出,早上新世存在一次全球性C4 扩张事件.然而,澳大利亚近海钻孔正构烷烃δ 13 C表明,C4 植被直至3.5 Ma才开始扩张[65 ] .同时,太平洋近赤道地区海洋钻孔长链脂肪酸δ 13 C[66 ] 和黄土高原西部临夏盆地牙釉δ 13 C[18 -19 ] 指示,C4 植被直至早更新世后才显著增加.此外,黄土高原东部灵台、蓝田剖面无机碳δ 13 C指示,C4 植被经历了2.9~2.7 Ma、1.3~0.9 Ma、0.6 Ma三次反复扩张[67 ] .基于全球各地C4 植被演化差异,学者们将控制C4 植被演化的因素归结于干旱程度[14 ,63 ,68 -69 ] 、暖季温度和降水[67 ,70 -71 ] 和野火[72 -73 ] 等区域环境因素.由此可见,关于晚中新世以来C4 植被演化的时间和机制等的认识仍未统一. ...
Carbon isotope ratio of Cenozoic CO2 : A comparative evaluation of available geochemical proxies
2
2010
... 为准确恢复和定量重建皋兰山C3 /C4 植被演变历史,本文利用端元法首先校正了大气δ 13 C C O 2 2 浓度变化带来的可能影响.假设植物的碳分馏能力固定不变,大气δ 13 C C O 2 δ 13 C的变化.因此,在进行古气候重建工作时应予以校正.本文利用底栖有孔虫建立的δ 13 C C O 2 [51 ] ,通过公式(1) 进行校正.同时,考虑到C3 植被δ 13 C水平(即C3 端元值)亦受大气CO2 浓度变化影响,本文利用浮游植物化石重建的大气CO2 浓度[52 ] 对C3 植被端元值进行校正.相关计算方法参照Schubert和Jahren[53 ] .ε C3 端元和ε C4 端元初始值引自[54 ] .校正后利用公式(2) 和公式(3) 计算出C4 植被所占比例. ...
...
图5 为端元法校正之后的结果,据此可将兰州地区皋兰山上新世-更新世转型期(3.0~2.2 Ma) C
3 /C
4 植被演化划分为两个阶段:
图5 皋兰山C<sub>4</sub>植被含量与深海氧同位素(a)、大气<span class="formulaText"><inline-formula><math id="M6"><msup><mrow><mi>δ</mi></mrow><mrow><mn mathvariant="normal">13</mn></mrow></msup><msub><mrow><mi mathvariant="normal">C</mi></mrow><mrow><mi mathvariant="normal">C</mi><msub><mrow><mi mathvariant="normal">O</mi></mrow><mrow><mn mathvariant="normal">2</mn></mrow></msub></mrow></msub></math></span></inline-formula></span> (b)、<i>p</i>CO<sub>2</sub> (c)和中值粒径(d)综合对比图 Comparisons between deep ocean <i>δ</i><sup>18</sup>O(a) from Lisieki and Raymo<sup>[<xref ref-type="bibr" rid="R56">56</xref>]</sup>, atmosphere <span class="formulaText"><inline-formula><math id="M7"><msup><mrow><mi>δ</mi></mrow><mrow><mn mathvariant="normal">13</mn></mrow></msup><msub><mrow><mi mathvariant="normal">C</mi></mrow><mrow><mi mathvariant="normal">C</mi><msub><mrow><mi mathvariant="normal">O</mi></mrow><mrow><mn mathvariant="normal">2</mn></mrow></msub></mrow></msub></math></span></inline-formula></span> (b) from Tipple et al.<sup>[<xref ref-type="bibr" rid="R51">51</xref>]</sup>, atmosphere <i>p</i>CO<sub>2</sub> (c) from Beerling and Royer<sup>[<xref ref-type="bibr" rid="R52">52</xref>]</sup> & Bartoli et al.<sup>[<xref ref-type="bibr" rid="R54">54</xref>]</sup>, median diameter (d) from Guo et al.<sup>[<xref ref-type="bibr" rid="R11">11</xref>]</sup>, and C<sub>4</sub> content (e). Atmosphere <i>p</i>CO<sub>2</sub> based on <i>δ</i><sup>11</sup>B, phytoplankton<sup>[<xref ref-type="bibr" rid="R52">52</xref>]</sup>, and <i>δ</i><sup>11</sup>B<sup>[<xref ref-type="bibr" rid="R55">55</xref>]</sup> are outlined by the orange, black, and pink plots and line, respectively Fig.5 ![]()
第一阶段:3.0~2.9 Ma时段,主体为红黏土沉积,C4 植被相对比例最高.上新世大暖期结束时(3.0 Ma),δ 13 Cn -alkane 整体较为偏正[图(3)和(5)],远高于C3 植被δ 13 Cn -alkane 分布范围(-39‰~-32‰).ACL值较低,指示气候整体温暖,但ACL逐渐增大,支持气候逐渐变冷.与此同时,ACL与δ 13 Cn -alkane 呈反相位变化,即气候越冷干,δ 13 Cn -alkane 值逐渐偏负,与黄土高原东部的C4 模式一致.据此推断:此时气候条件仍相对温暖湿润,适宜C4 植被发育,δ 13 Cn -alkane 由C4 植被相对丰度变化主导.定量重建结果表明,此时段C4 植被比例最高可达40%(图5 ).2.9~2.8 Ma,ACL值逐渐降低,磁化率较之前明显增加[47 ] ,表明此时降水较之前有所增多,相对充沛.而兰州位于季风影响的边缘区域,雨热同期主要集中在生长季(夏季).因此,强降水也意味着季节性加强,更适宜C4 生长.但此时C4 植被仍稳定在低水平,说明降水不是C4 植被减少的主要因素.同时,此时段p CO2 略微降低[52 ,55 ] (图5 ),也更适宜C4 发育.排除前述因子后,C4 植被生长的限制因素极可能为温度.研究表明,上新世温暖期(3.3~3.0 Ma)结束后,氧同位素揭示的全球冰量显著扩张[56 ] ,温度进一步下降.这可能逐渐突破了C4 植被生长阈值,进而导致兰州皋兰山C4 植被快速减少. ...
Convergent Cenozoic CO2 history
4
2011
... 为准确恢复和定量重建皋兰山C3 /C4 植被演变历史,本文利用端元法首先校正了大气δ 13 C C O 2 2 浓度变化带来的可能影响.假设植物的碳分馏能力固定不变,大气δ 13 C C O 2 δ 13 C的变化.因此,在进行古气候重建工作时应予以校正.本文利用底栖有孔虫建立的δ 13 C C O 2 [51 ] ,通过公式(1) 进行校正.同时,考虑到C3 植被δ 13 C水平(即C3 端元值)亦受大气CO2 浓度变化影响,本文利用浮游植物化石重建的大气CO2 浓度[52 ] 对C3 植被端元值进行校正.相关计算方法参照Schubert和Jahren[53 ] .ε C3 端元和ε C4 端元初始值引自[54 ] .校正后利用公式(2) 和公式(3) 计算出C4 植被所占比例. ...
...
图5 为端元法校正之后的结果,据此可将兰州地区皋兰山上新世-更新世转型期(3.0~2.2 Ma) C
3 /C
4 植被演化划分为两个阶段:
图5 皋兰山C<sub>4</sub>植被含量与深海氧同位素(a)、大气<span class="formulaText"><inline-formula><math id="M6"><msup><mrow><mi>δ</mi></mrow><mrow><mn mathvariant="normal">13</mn></mrow></msup><msub><mrow><mi mathvariant="normal">C</mi></mrow><mrow><mi mathvariant="normal">C</mi><msub><mrow><mi mathvariant="normal">O</mi></mrow><mrow><mn mathvariant="normal">2</mn></mrow></msub></mrow></msub></math></span></inline-formula></span> (b)、<i>p</i>CO<sub>2</sub> (c)和中值粒径(d)综合对比图 Comparisons between deep ocean <i>δ</i><sup>18</sup>O(a) from Lisieki and Raymo<sup>[<xref ref-type="bibr" rid="R56">56</xref>]</sup>, atmosphere <span class="formulaText"><inline-formula><math id="M7"><msup><mrow><mi>δ</mi></mrow><mrow><mn mathvariant="normal">13</mn></mrow></msup><msub><mrow><mi mathvariant="normal">C</mi></mrow><mrow><mi mathvariant="normal">C</mi><msub><mrow><mi mathvariant="normal">O</mi></mrow><mrow><mn mathvariant="normal">2</mn></mrow></msub></mrow></msub></math></span></inline-formula></span> (b) from Tipple et al.<sup>[<xref ref-type="bibr" rid="R51">51</xref>]</sup>, atmosphere <i>p</i>CO<sub>2</sub> (c) from Beerling and Royer<sup>[<xref ref-type="bibr" rid="R52">52</xref>]</sup> & Bartoli et al.<sup>[<xref ref-type="bibr" rid="R54">54</xref>]</sup>, median diameter (d) from Guo et al.<sup>[<xref ref-type="bibr" rid="R11">11</xref>]</sup>, and C<sub>4</sub> content (e). Atmosphere <i>p</i>CO<sub>2</sub> based on <i>δ</i><sup>11</sup>B, phytoplankton<sup>[<xref ref-type="bibr" rid="R52">52</xref>]</sup>, and <i>δ</i><sup>11</sup>B<sup>[<xref ref-type="bibr" rid="R55">55</xref>]</sup> are outlined by the orange, black, and pink plots and line, respectively Fig.5 ![]()
第一阶段:3.0~2.9 Ma时段,主体为红黏土沉积,C4 植被相对比例最高.上新世大暖期结束时(3.0 Ma),δ 13 Cn -alkane 整体较为偏正[图(3)和(5)],远高于C3 植被δ 13 Cn -alkane 分布范围(-39‰~-32‰).ACL值较低,指示气候整体温暖,但ACL逐渐增大,支持气候逐渐变冷.与此同时,ACL与δ 13 Cn -alkane 呈反相位变化,即气候越冷干,δ 13 Cn -alkane 值逐渐偏负,与黄土高原东部的C4 模式一致.据此推断:此时气候条件仍相对温暖湿润,适宜C4 植被发育,δ 13 Cn -alkane 由C4 植被相对丰度变化主导.定量重建结果表明,此时段C4 植被比例最高可达40%(图5 ).2.9~2.8 Ma,ACL值逐渐降低,磁化率较之前明显增加[47 ] ,表明此时降水较之前有所增多,相对充沛.而兰州位于季风影响的边缘区域,雨热同期主要集中在生长季(夏季).因此,强降水也意味着季节性加强,更适宜C4 生长.但此时C4 植被仍稳定在低水平,说明降水不是C4 植被减少的主要因素.同时,此时段p CO2 略微降低[52 ,55 ] (图5 ),也更适宜C4 发育.排除前述因子后,C4 植被生长的限制因素极可能为温度.研究表明,上新世温暖期(3.3~3.0 Ma)结束后,氧同位素揭示的全球冰量显著扩张[56 ] ,温度进一步下降.这可能逐渐突破了C4 植被生长阈值,进而导致兰州皋兰山C4 植被快速减少. ...
... [
52 ], and
δ 11 B
[55 ] are outlined by the orange, black, and pink plots and line, respectively
Fig.5 ![]()
第一阶段:3.0~2.9 Ma时段,主体为红黏土沉积,C4 植被相对比例最高.上新世大暖期结束时(3.0 Ma),δ 13 Cn -alkane 整体较为偏正[图(3)和(5)],远高于C3 植被δ 13 Cn -alkane 分布范围(-39‰~-32‰).ACL值较低,指示气候整体温暖,但ACL逐渐增大,支持气候逐渐变冷.与此同时,ACL与δ 13 Cn -alkane 呈反相位变化,即气候越冷干,δ 13 Cn -alkane 值逐渐偏负,与黄土高原东部的C4 模式一致.据此推断:此时气候条件仍相对温暖湿润,适宜C4 植被发育,δ 13 Cn -alkane 由C4 植被相对丰度变化主导.定量重建结果表明,此时段C4 植被比例最高可达40%(图5 ).2.9~2.8 Ma,ACL值逐渐降低,磁化率较之前明显增加[47 ] ,表明此时降水较之前有所增多,相对充沛.而兰州位于季风影响的边缘区域,雨热同期主要集中在生长季(夏季).因此,强降水也意味着季节性加强,更适宜C4 生长.但此时C4 植被仍稳定在低水平,说明降水不是C4 植被减少的主要因素.同时,此时段p CO2 略微降低[52 ,55 ] (图5 ),也更适宜C4 发育.排除前述因子后,C4 植被生长的限制因素极可能为温度.研究表明,上新世温暖期(3.3~3.0 Ma)结束后,氧同位素揭示的全球冰量显著扩张[56 ] ,温度进一步下降.这可能逐渐突破了C4 植被生长阈值,进而导致兰州皋兰山C4 植被快速减少. ...
... 第一阶段:3.0~2.9 Ma时段,主体为红黏土沉积,C4 植被相对比例最高.上新世大暖期结束时(3.0 Ma),δ 13 Cn -alkane 整体较为偏正[图(3)和(5)],远高于C3 植被δ 13 Cn -alkane 分布范围(-39‰~-32‰).ACL值较低,指示气候整体温暖,但ACL逐渐增大,支持气候逐渐变冷.与此同时,ACL与δ 13 Cn -alkane 呈反相位变化,即气候越冷干,δ 13 Cn -alkane 值逐渐偏负,与黄土高原东部的C4 模式一致.据此推断:此时气候条件仍相对温暖湿润,适宜C4 植被发育,δ 13 Cn -alkane 由C4 植被相对丰度变化主导.定量重建结果表明,此时段C4 植被比例最高可达40%(图5 ).2.9~2.8 Ma,ACL值逐渐降低,磁化率较之前明显增加[47 ] ,表明此时降水较之前有所增多,相对充沛.而兰州位于季风影响的边缘区域,雨热同期主要集中在生长季(夏季).因此,强降水也意味着季节性加强,更适宜C4 生长.但此时C4 植被仍稳定在低水平,说明降水不是C4 植被减少的主要因素.同时,此时段p CO2 略微降低[52 ,55 ] (图5 ),也更适宜C4 发育.排除前述因子后,C4 植被生长的限制因素极可能为温度.研究表明,上新世温暖期(3.3~3.0 Ma)结束后,氧同位素揭示的全球冰量显著扩张[56 ] ,温度进一步下降.这可能逐渐突破了C4 植被生长阈值,进而导致兰州皋兰山C4 植被快速减少. ...
Global increase in plant carbon isotope fractionation following the last glacial maximum caused by increase in atmospheric p CO2
1
2015
... 为准确恢复和定量重建皋兰山C3 /C4 植被演变历史,本文利用端元法首先校正了大气δ 13 C C O 2 2 浓度变化带来的可能影响.假设植物的碳分馏能力固定不变,大气δ 13 C C O 2 δ 13 C的变化.因此,在进行古气候重建工作时应予以校正.本文利用底栖有孔虫建立的δ 13 C C O 2 [51 ] ,通过公式(1) 进行校正.同时,考虑到C3 植被δ 13 C水平(即C3 端元值)亦受大气CO2 浓度变化影响,本文利用浮游植物化石重建的大气CO2 浓度[52 ] 对C3 植被端元值进行校正.相关计算方法参照Schubert和Jahren[53 ] .ε C3 端元和ε C4 端元初始值引自[54 ] .校正后利用公式(2) 和公式(3) 计算出C4 植被所占比例. ...
A 35 Myr North American leaf-wax compound-specific carbon and hydrogen isotope record: Implications for C4 grasslands and hydrologic cycle dynamics
2
2010
... 为准确恢复和定量重建皋兰山C3 /C4 植被演变历史,本文利用端元法首先校正了大气δ 13 C C O 2 2 浓度变化带来的可能影响.假设植物的碳分馏能力固定不变,大气δ 13 C C O 2 δ 13 C的变化.因此,在进行古气候重建工作时应予以校正.本文利用底栖有孔虫建立的δ 13 C C O 2 [51 ] ,通过公式(1) 进行校正.同时,考虑到C3 植被δ 13 C水平(即C3 端元值)亦受大气CO2 浓度变化影响,本文利用浮游植物化石重建的大气CO2 浓度[52 ] 对C3 植被端元值进行校正.相关计算方法参照Schubert和Jahren[53 ] .ε C3 端元和ε C4 端元初始值引自[54 ] .校正后利用公式(2) 和公式(3) 计算出C4 植被所占比例. ...
...
图5 为端元法校正之后的结果,据此可将兰州地区皋兰山上新世-更新世转型期(3.0~2.2 Ma) C
3 /C
4 植被演化划分为两个阶段:
图5 皋兰山C<sub>4</sub>植被含量与深海氧同位素(a)、大气<span class="formulaText"><inline-formula><math id="M6"><msup><mrow><mi>δ</mi></mrow><mrow><mn mathvariant="normal">13</mn></mrow></msup><msub><mrow><mi mathvariant="normal">C</mi></mrow><mrow><mi mathvariant="normal">C</mi><msub><mrow><mi mathvariant="normal">O</mi></mrow><mrow><mn mathvariant="normal">2</mn></mrow></msub></mrow></msub></math></span></inline-formula></span> (b)、<i>p</i>CO<sub>2</sub> (c)和中值粒径(d)综合对比图 Comparisons between deep ocean <i>δ</i><sup>18</sup>O(a) from Lisieki and Raymo<sup>[<xref ref-type="bibr" rid="R56">56</xref>]</sup>, atmosphere <span class="formulaText"><inline-formula><math id="M7"><msup><mrow><mi>δ</mi></mrow><mrow><mn mathvariant="normal">13</mn></mrow></msup><msub><mrow><mi mathvariant="normal">C</mi></mrow><mrow><mi mathvariant="normal">C</mi><msub><mrow><mi mathvariant="normal">O</mi></mrow><mrow><mn mathvariant="normal">2</mn></mrow></msub></mrow></msub></math></span></inline-formula></span> (b) from Tipple et al.<sup>[<xref ref-type="bibr" rid="R51">51</xref>]</sup>, atmosphere <i>p</i>CO<sub>2</sub> (c) from Beerling and Royer<sup>[<xref ref-type="bibr" rid="R52">52</xref>]</sup> & Bartoli et al.<sup>[<xref ref-type="bibr" rid="R54">54</xref>]</sup>, median diameter (d) from Guo et al.<sup>[<xref ref-type="bibr" rid="R11">11</xref>]</sup>, and C<sub>4</sub> content (e). Atmosphere <i>p</i>CO<sub>2</sub> based on <i>δ</i><sup>11</sup>B, phytoplankton<sup>[<xref ref-type="bibr" rid="R52">52</xref>]</sup>, and <i>δ</i><sup>11</sup>B<sup>[<xref ref-type="bibr" rid="R55">55</xref>]</sup> are outlined by the orange, black, and pink plots and line, respectively Fig.5 ![]()
第一阶段:3.0~2.9 Ma时段,主体为红黏土沉积,C4 植被相对比例最高.上新世大暖期结束时(3.0 Ma),δ 13 Cn -alkane 整体较为偏正[图(3)和(5)],远高于C3 植被δ 13 Cn -alkane 分布范围(-39‰~-32‰).ACL值较低,指示气候整体温暖,但ACL逐渐增大,支持气候逐渐变冷.与此同时,ACL与δ 13 Cn -alkane 呈反相位变化,即气候越冷干,δ 13 Cn -alkane 值逐渐偏负,与黄土高原东部的C4 模式一致.据此推断:此时气候条件仍相对温暖湿润,适宜C4 植被发育,δ 13 Cn -alkane 由C4 植被相对丰度变化主导.定量重建结果表明,此时段C4 植被比例最高可达40%(图5 ).2.9~2.8 Ma,ACL值逐渐降低,磁化率较之前明显增加[47 ] ,表明此时降水较之前有所增多,相对充沛.而兰州位于季风影响的边缘区域,雨热同期主要集中在生长季(夏季).因此,强降水也意味着季节性加强,更适宜C4 生长.但此时C4 植被仍稳定在低水平,说明降水不是C4 植被减少的主要因素.同时,此时段p CO2 略微降低[52 ,55 ] (图5 ),也更适宜C4 发育.排除前述因子后,C4 植被生长的限制因素极可能为温度.研究表明,上新世温暖期(3.3~3.0 Ma)结束后,氧同位素揭示的全球冰量显著扩张[56 ] ,温度进一步下降.这可能逐渐突破了C4 植被生长阈值,进而导致兰州皋兰山C4 植被快速减少. ...
Atmospheric CO2 decline during the Pliocene intensification of Northern Hemisphere glaciations
2
2011
...
图5 为端元法校正之后的结果,据此可将兰州地区皋兰山上新世-更新世转型期(3.0~2.2 Ma) C
3 /C
4 植被演化划分为两个阶段:
图5 皋兰山C<sub>4</sub>植被含量与深海氧同位素(a)、大气<span class="formulaText"><inline-formula><math id="M6"><msup><mrow><mi>δ</mi></mrow><mrow><mn mathvariant="normal">13</mn></mrow></msup><msub><mrow><mi mathvariant="normal">C</mi></mrow><mrow><mi mathvariant="normal">C</mi><msub><mrow><mi mathvariant="normal">O</mi></mrow><mrow><mn mathvariant="normal">2</mn></mrow></msub></mrow></msub></math></span></inline-formula></span> (b)、<i>p</i>CO<sub>2</sub> (c)和中值粒径(d)综合对比图 Comparisons between deep ocean <i>δ</i><sup>18</sup>O(a) from Lisieki and Raymo<sup>[<xref ref-type="bibr" rid="R56">56</xref>]</sup>, atmosphere <span class="formulaText"><inline-formula><math id="M7"><msup><mrow><mi>δ</mi></mrow><mrow><mn mathvariant="normal">13</mn></mrow></msup><msub><mrow><mi mathvariant="normal">C</mi></mrow><mrow><mi mathvariant="normal">C</mi><msub><mrow><mi mathvariant="normal">O</mi></mrow><mrow><mn mathvariant="normal">2</mn></mrow></msub></mrow></msub></math></span></inline-formula></span> (b) from Tipple et al.<sup>[<xref ref-type="bibr" rid="R51">51</xref>]</sup>, atmosphere <i>p</i>CO<sub>2</sub> (c) from Beerling and Royer<sup>[<xref ref-type="bibr" rid="R52">52</xref>]</sup> & Bartoli et al.<sup>[<xref ref-type="bibr" rid="R54">54</xref>]</sup>, median diameter (d) from Guo et al.<sup>[<xref ref-type="bibr" rid="R11">11</xref>]</sup>, and C<sub>4</sub> content (e). Atmosphere <i>p</i>CO<sub>2</sub> based on <i>δ</i><sup>11</sup>B, phytoplankton<sup>[<xref ref-type="bibr" rid="R52">52</xref>]</sup>, and <i>δ</i><sup>11</sup>B<sup>[<xref ref-type="bibr" rid="R55">55</xref>]</sup> are outlined by the orange, black, and pink plots and line, respectively Fig.5 ![]()
第一阶段:3.0~2.9 Ma时段,主体为红黏土沉积,C4 植被相对比例最高.上新世大暖期结束时(3.0 Ma),δ 13 Cn -alkane 整体较为偏正[图(3)和(5)],远高于C3 植被δ 13 Cn -alkane 分布范围(-39‰~-32‰).ACL值较低,指示气候整体温暖,但ACL逐渐增大,支持气候逐渐变冷.与此同时,ACL与δ 13 Cn -alkane 呈反相位变化,即气候越冷干,δ 13 Cn -alkane 值逐渐偏负,与黄土高原东部的C4 模式一致.据此推断:此时气候条件仍相对温暖湿润,适宜C4 植被发育,δ 13 Cn -alkane 由C4 植被相对丰度变化主导.定量重建结果表明,此时段C4 植被比例最高可达40%(图5 ).2.9~2.8 Ma,ACL值逐渐降低,磁化率较之前明显增加[47 ] ,表明此时降水较之前有所增多,相对充沛.而兰州位于季风影响的边缘区域,雨热同期主要集中在生长季(夏季).因此,强降水也意味着季节性加强,更适宜C4 生长.但此时C4 植被仍稳定在低水平,说明降水不是C4 植被减少的主要因素.同时,此时段p CO2 略微降低[52 ,55 ] (图5 ),也更适宜C4 发育.排除前述因子后,C4 植被生长的限制因素极可能为温度.研究表明,上新世温暖期(3.3~3.0 Ma)结束后,氧同位素揭示的全球冰量显著扩张[56 ] ,温度进一步下降.这可能逐渐突破了C4 植被生长阈值,进而导致兰州皋兰山C4 植被快速减少. ...
... 第一阶段:3.0~2.9 Ma时段,主体为红黏土沉积,C4 植被相对比例最高.上新世大暖期结束时(3.0 Ma),δ 13 Cn -alkane 整体较为偏正[图(3)和(5)],远高于C3 植被δ 13 Cn -alkane 分布范围(-39‰~-32‰).ACL值较低,指示气候整体温暖,但ACL逐渐增大,支持气候逐渐变冷.与此同时,ACL与δ 13 Cn -alkane 呈反相位变化,即气候越冷干,δ 13 Cn -alkane 值逐渐偏负,与黄土高原东部的C4 模式一致.据此推断:此时气候条件仍相对温暖湿润,适宜C4 植被发育,δ 13 Cn -alkane 由C4 植被相对丰度变化主导.定量重建结果表明,此时段C4 植被比例最高可达40%(图5 ).2.9~2.8 Ma,ACL值逐渐降低,磁化率较之前明显增加[47 ] ,表明此时降水较之前有所增多,相对充沛.而兰州位于季风影响的边缘区域,雨热同期主要集中在生长季(夏季).因此,强降水也意味着季节性加强,更适宜C4 生长.但此时C4 植被仍稳定在低水平,说明降水不是C4 植被减少的主要因素.同时,此时段p CO2 略微降低[52 ,55 ] (图5 ),也更适宜C4 发育.排除前述因子后,C4 植被生长的限制因素极可能为温度.研究表明,上新世温暖期(3.3~3.0 Ma)结束后,氧同位素揭示的全球冰量显著扩张[56 ] ,温度进一步下降.这可能逐渐突破了C4 植被生长阈值,进而导致兰州皋兰山C4 植被快速减少. ...
A Pliocene-Pleistocene stack of 57 globally distributed benthic δ 18 O records
2
2005
...
图5 为端元法校正之后的结果,据此可将兰州地区皋兰山上新世-更新世转型期(3.0~2.2 Ma) C
3 /C
4 植被演化划分为两个阶段:
图5 皋兰山C<sub>4</sub>植被含量与深海氧同位素(a)、大气<span class="formulaText"><inline-formula><math id="M6"><msup><mrow><mi>δ</mi></mrow><mrow><mn mathvariant="normal">13</mn></mrow></msup><msub><mrow><mi mathvariant="normal">C</mi></mrow><mrow><mi mathvariant="normal">C</mi><msub><mrow><mi mathvariant="normal">O</mi></mrow><mrow><mn mathvariant="normal">2</mn></mrow></msub></mrow></msub></math></span></inline-formula></span> (b)、<i>p</i>CO<sub>2</sub> (c)和中值粒径(d)综合对比图 Comparisons between deep ocean <i>δ</i><sup>18</sup>O(a) from Lisieki and Raymo<sup>[<xref ref-type="bibr" rid="R56">56</xref>]</sup>, atmosphere <span class="formulaText"><inline-formula><math id="M7"><msup><mrow><mi>δ</mi></mrow><mrow><mn mathvariant="normal">13</mn></mrow></msup><msub><mrow><mi mathvariant="normal">C</mi></mrow><mrow><mi mathvariant="normal">C</mi><msub><mrow><mi mathvariant="normal">O</mi></mrow><mrow><mn mathvariant="normal">2</mn></mrow></msub></mrow></msub></math></span></inline-formula></span> (b) from Tipple et al.<sup>[<xref ref-type="bibr" rid="R51">51</xref>]</sup>, atmosphere <i>p</i>CO<sub>2</sub> (c) from Beerling and Royer<sup>[<xref ref-type="bibr" rid="R52">52</xref>]</sup> & Bartoli et al.<sup>[<xref ref-type="bibr" rid="R54">54</xref>]</sup>, median diameter (d) from Guo et al.<sup>[<xref ref-type="bibr" rid="R11">11</xref>]</sup>, and C<sub>4</sub> content (e). Atmosphere <i>p</i>CO<sub>2</sub> based on <i>δ</i><sup>11</sup>B, phytoplankton<sup>[<xref ref-type="bibr" rid="R52">52</xref>]</sup>, and <i>δ</i><sup>11</sup>B<sup>[<xref ref-type="bibr" rid="R55">55</xref>]</sup> are outlined by the orange, black, and pink plots and line, respectively Fig.5 ![]()
第一阶段:3.0~2.9 Ma时段,主体为红黏土沉积,C4 植被相对比例最高.上新世大暖期结束时(3.0 Ma),δ 13 Cn -alkane 整体较为偏正[图(3)和(5)],远高于C3 植被δ 13 Cn -alkane 分布范围(-39‰~-32‰).ACL值较低,指示气候整体温暖,但ACL逐渐增大,支持气候逐渐变冷.与此同时,ACL与δ 13 Cn -alkane 呈反相位变化,即气候越冷干,δ 13 Cn -alkane 值逐渐偏负,与黄土高原东部的C4 模式一致.据此推断:此时气候条件仍相对温暖湿润,适宜C4 植被发育,δ 13 Cn -alkane 由C4 植被相对丰度变化主导.定量重建结果表明,此时段C4 植被比例最高可达40%(图5 ).2.9~2.8 Ma,ACL值逐渐降低,磁化率较之前明显增加[47 ] ,表明此时降水较之前有所增多,相对充沛.而兰州位于季风影响的边缘区域,雨热同期主要集中在生长季(夏季).因此,强降水也意味着季节性加强,更适宜C4 生长.但此时C4 植被仍稳定在低水平,说明降水不是C4 植被减少的主要因素.同时,此时段p CO2 略微降低[52 ,55 ] (图5 ),也更适宜C4 发育.排除前述因子后,C4 植被生长的限制因素极可能为温度.研究表明,上新世温暖期(3.3~3.0 Ma)结束后,氧同位素揭示的全球冰量显著扩张[56 ] ,温度进一步下降.这可能逐渐突破了C4 植被生长阈值,进而导致兰州皋兰山C4 植被快速减少. ...
... 第一阶段:3.0~2.9 Ma时段,主体为红黏土沉积,C4 植被相对比例最高.上新世大暖期结束时(3.0 Ma),δ 13 Cn -alkane 整体较为偏正[图(3)和(5)],远高于C3 植被δ 13 Cn -alkane 分布范围(-39‰~-32‰).ACL值较低,指示气候整体温暖,但ACL逐渐增大,支持气候逐渐变冷.与此同时,ACL与δ 13 Cn -alkane 呈反相位变化,即气候越冷干,δ 13 Cn -alkane 值逐渐偏负,与黄土高原东部的C4 模式一致.据此推断:此时气候条件仍相对温暖湿润,适宜C4 植被发育,δ 13 Cn -alkane 由C4 植被相对丰度变化主导.定量重建结果表明,此时段C4 植被比例最高可达40%(图5 ).2.9~2.8 Ma,ACL值逐渐降低,磁化率较之前明显增加[47 ] ,表明此时降水较之前有所增多,相对充沛.而兰州位于季风影响的边缘区域,雨热同期主要集中在生长季(夏季).因此,强降水也意味着季节性加强,更适宜C4 生长.但此时C4 植被仍稳定在低水平,说明降水不是C4 植被减少的主要因素.同时,此时段p CO2 略微降低[52 ,55 ] (图5 ),也更适宜C4 发育.排除前述因子后,C4 植被生长的限制因素极可能为温度.研究表明,上新世温暖期(3.3~3.0 Ma)结束后,氧同位素揭示的全球冰量显著扩张[56 ] ,温度进一步下降.这可能逐渐突破了C4 植被生长阈值,进而导致兰州皋兰山C4 植被快速减少. ...
The Origins of C4 grasslands: Integrating evolutionary and ecosystem science
1
2010
... C4 植被的出现可以追溯到32~30 Ma,其大幅扩张一般认为发生在晚中新世[57 ] .来自巴基斯坦Siwalik沉积有机质和无机碳[58 ] 、北美哺乳动物牙釉化石[59 ] 、南美东部钻孔无机碳和动物化石[60 ] 和东非钻孔中动物化石[61 ] 等研究结果均支持这一说法:晚中新世7~8 Ma左右全球发生C4 植被的显著扩张.因此,C4 植被扩张是由大气CO2 浓度降低驱动的全球生态事件[62 ] .21世纪以来,越来越多的记录表明,全球C4 植被扩张的时间和范围并不一致:非洲东北部正构烷烃δ 13 C表明,C4 植被在11 Ma业已达到较高水平[50 ] ;新疆塔里木盆地钻孔有机碳δ 13 C显示,C4 植被在5.3 Ma显著增加[63 ] .这也得到日本海钻孔正构烷烃δ 13 C和内蒙古哺乳动物牙釉化石δ 13 C结果的支持[14 ,64 ] .近来,Lu等[15 ] 汇总全球记录后提出,早上新世存在一次全球性C4 扩张事件.然而,澳大利亚近海钻孔正构烷烃δ 13 C表明,C4 植被直至3.5 Ma才开始扩张[65 ] .同时,太平洋近赤道地区海洋钻孔长链脂肪酸δ 13 C[66 ] 和黄土高原西部临夏盆地牙釉δ 13 C[18 -19 ] 指示,C4 植被直至早更新世后才显著增加.此外,黄土高原东部灵台、蓝田剖面无机碳δ 13 C指示,C4 植被经历了2.9~2.7 Ma、1.3~0.9 Ma、0.6 Ma三次反复扩张[67 ] .基于全球各地C4 植被演化差异,学者们将控制C4 植被演化的因素归结于干旱程度[14 ,63 ,68 -69 ] 、暖季温度和降水[67 ,70 -71 ] 和野火[72 -73 ] 等区域环境因素.由此可见,关于晚中新世以来C4 植被演化的时间和机制等的认识仍未统一. ...
Development of Asian monsoon revealed by marked ecological shift during the latest Miocene in
1
1989
... C4 植被的出现可以追溯到32~30 Ma,其大幅扩张一般认为发生在晚中新世[57 ] .来自巴基斯坦Siwalik沉积有机质和无机碳[58 ] 、北美哺乳动物牙釉化石[59 ] 、南美东部钻孔无机碳和动物化石[60 ] 和东非钻孔中动物化石[61 ] 等研究结果均支持这一说法:晚中新世7~8 Ma左右全球发生C4 植被的显著扩张.因此,C4 植被扩张是由大气CO2 浓度降低驱动的全球生态事件[62 ] .21世纪以来,越来越多的记录表明,全球C4 植被扩张的时间和范围并不一致:非洲东北部正构烷烃δ 13 C表明,C4 植被在11 Ma业已达到较高水平[50 ] ;新疆塔里木盆地钻孔有机碳δ 13 C显示,C4 植被在5.3 Ma显著增加[63 ] .这也得到日本海钻孔正构烷烃δ 13 C和内蒙古哺乳动物牙釉化石δ 13 C结果的支持[14 ,64 ] .近来,Lu等[15 ] 汇总全球记录后提出,早上新世存在一次全球性C4 扩张事件.然而,澳大利亚近海钻孔正构烷烃δ 13 C表明,C4 植被直至3.5 Ma才开始扩张[65 ] .同时,太平洋近赤道地区海洋钻孔长链脂肪酸δ 13 C[66 ] 和黄土高原西部临夏盆地牙釉δ 13 C[18 -19 ] 指示,C4 植被直至早更新世后才显著增加.此外,黄土高原东部灵台、蓝田剖面无机碳δ 13 C指示,C4 植被经历了2.9~2.7 Ma、1.3~0.9 Ma、0.6 Ma三次反复扩张[67 ] .基于全球各地C4 植被演化差异,学者们将控制C4 植被演化的因素归结于干旱程度[14 ,63 ,68 -69 ] 、暖季温度和降水[67 ,70 -71 ] 和野火[72 -73 ] 等区域环境因素.由此可见,关于晚中新世以来C4 植被演化的时间和机制等的认识仍未统一. ...
Mammalian herbivore communities, ancient feeding ecology, and carbon isotopes: A 10 million-year sequence from the Neogene of Florida
1
1996
... C4 植被的出现可以追溯到32~30 Ma,其大幅扩张一般认为发生在晚中新世[57 ] .来自巴基斯坦Siwalik沉积有机质和无机碳[58 ] 、北美哺乳动物牙釉化石[59 ] 、南美东部钻孔无机碳和动物化石[60 ] 和东非钻孔中动物化石[61 ] 等研究结果均支持这一说法:晚中新世7~8 Ma左右全球发生C4 植被的显著扩张.因此,C4 植被扩张是由大气CO2 浓度降低驱动的全球生态事件[62 ] .21世纪以来,越来越多的记录表明,全球C4 植被扩张的时间和范围并不一致:非洲东北部正构烷烃δ 13 C表明,C4 植被在11 Ma业已达到较高水平[50 ] ;新疆塔里木盆地钻孔有机碳δ 13 C显示,C4 植被在5.3 Ma显著增加[63 ] .这也得到日本海钻孔正构烷烃δ 13 C和内蒙古哺乳动物牙釉化石δ 13 C结果的支持[14 ,64 ] .近来,Lu等[15 ] 汇总全球记录后提出,早上新世存在一次全球性C4 扩张事件.然而,澳大利亚近海钻孔正构烷烃δ 13 C表明,C4 植被直至3.5 Ma才开始扩张[65 ] .同时,太平洋近赤道地区海洋钻孔长链脂肪酸δ 13 C[66 ] 和黄土高原西部临夏盆地牙釉δ 13 C[18 -19 ] 指示,C4 植被直至早更新世后才显著增加.此外,黄土高原东部灵台、蓝田剖面无机碳δ 13 C指示,C4 植被经历了2.9~2.7 Ma、1.3~0.9 Ma、0.6 Ma三次反复扩张[67 ] .基于全球各地C4 植被演化差异,学者们将控制C4 植被演化的因素归结于干旱程度[14 ,63 ,68 -69 ] 、暖季温度和降水[67 ,70 -71 ] 和野火[72 -73 ] 等区域环境因素.由此可见,关于晚中新世以来C4 植被演化的时间和机制等的认识仍未统一. ...
The expansion of C4 grasses and global change in the late Miocene: stable isotope evidence from the Americas
1
1997
... C4 植被的出现可以追溯到32~30 Ma,其大幅扩张一般认为发生在晚中新世[57 ] .来自巴基斯坦Siwalik沉积有机质和无机碳[58 ] 、北美哺乳动物牙釉化石[59 ] 、南美东部钻孔无机碳和动物化石[60 ] 和东非钻孔中动物化石[61 ] 等研究结果均支持这一说法:晚中新世7~8 Ma左右全球发生C4 植被的显著扩张.因此,C4 植被扩张是由大气CO2 浓度降低驱动的全球生态事件[62 ] .21世纪以来,越来越多的记录表明,全球C4 植被扩张的时间和范围并不一致:非洲东北部正构烷烃δ 13 C表明,C4 植被在11 Ma业已达到较高水平[50 ] ;新疆塔里木盆地钻孔有机碳δ 13 C显示,C4 植被在5.3 Ma显著增加[63 ] .这也得到日本海钻孔正构烷烃δ 13 C和内蒙古哺乳动物牙釉化石δ 13 C结果的支持[14 ,64 ] .近来,Lu等[15 ] 汇总全球记录后提出,早上新世存在一次全球性C4 扩张事件.然而,澳大利亚近海钻孔正构烷烃δ 13 C表明,C4 植被直至3.5 Ma才开始扩张[65 ] .同时,太平洋近赤道地区海洋钻孔长链脂肪酸δ 13 C[66 ] 和黄土高原西部临夏盆地牙釉δ 13 C[18 -19 ] 指示,C4 植被直至早更新世后才显著增加.此外,黄土高原东部灵台、蓝田剖面无机碳δ 13 C指示,C4 植被经历了2.9~2.7 Ma、1.3~0.9 Ma、0.6 Ma三次反复扩张[67 ] .基于全球各地C4 植被演化差异,学者们将控制C4 植被演化的因素归结于干旱程度[14 ,63 ,68 -69 ] 、暖季温度和降水[67 ,70 -71 ] 和野火[72 -73 ] 等区域环境因素.由此可见,关于晚中新世以来C4 植被演化的时间和机制等的认识仍未统一. ...
Carbon isotopic evidence for the emergence of C4 plants in the Neogene from Pakistan and Kenya
1
1994
... C4 植被的出现可以追溯到32~30 Ma,其大幅扩张一般认为发生在晚中新世[57 ] .来自巴基斯坦Siwalik沉积有机质和无机碳[58 ] 、北美哺乳动物牙釉化石[59 ] 、南美东部钻孔无机碳和动物化石[60 ] 和东非钻孔中动物化石[61 ] 等研究结果均支持这一说法:晚中新世7~8 Ma左右全球发生C4 植被的显著扩张.因此,C4 植被扩张是由大气CO2 浓度降低驱动的全球生态事件[62 ] .21世纪以来,越来越多的记录表明,全球C4 植被扩张的时间和范围并不一致:非洲东北部正构烷烃δ 13 C表明,C4 植被在11 Ma业已达到较高水平[50 ] ;新疆塔里木盆地钻孔有机碳δ 13 C显示,C4 植被在5.3 Ma显著增加[63 ] .这也得到日本海钻孔正构烷烃δ 13 C和内蒙古哺乳动物牙釉化石δ 13 C结果的支持[14 ,64 ] .近来,Lu等[15 ] 汇总全球记录后提出,早上新世存在一次全球性C4 扩张事件.然而,澳大利亚近海钻孔正构烷烃δ 13 C表明,C4 植被直至3.5 Ma才开始扩张[65 ] .同时,太平洋近赤道地区海洋钻孔长链脂肪酸δ 13 C[66 ] 和黄土高原西部临夏盆地牙釉δ 13 C[18 -19 ] 指示,C4 植被直至早更新世后才显著增加.此外,黄土高原东部灵台、蓝田剖面无机碳δ 13 C指示,C4 植被经历了2.9~2.7 Ma、1.3~0.9 Ma、0.6 Ma三次反复扩张[67 ] .基于全球各地C4 植被演化差异,学者们将控制C4 植被演化的因素归结于干旱程度[14 ,63 ,68 -69 ] 、暖季温度和降水[67 ,70 -71 ] 和野火[72 -73 ] 等区域环境因素.由此可见,关于晚中新世以来C4 植被演化的时间和机制等的认识仍未统一. ...
Global vegetation change through the Miocene/Pliocene boundary
1
1997
... C4 植被的出现可以追溯到32~30 Ma,其大幅扩张一般认为发生在晚中新世[57 ] .来自巴基斯坦Siwalik沉积有机质和无机碳[58 ] 、北美哺乳动物牙釉化石[59 ] 、南美东部钻孔无机碳和动物化石[60 ] 和东非钻孔中动物化石[61 ] 等研究结果均支持这一说法:晚中新世7~8 Ma左右全球发生C4 植被的显著扩张.因此,C4 植被扩张是由大气CO2 浓度降低驱动的全球生态事件[62 ] .21世纪以来,越来越多的记录表明,全球C4 植被扩张的时间和范围并不一致:非洲东北部正构烷烃δ 13 C表明,C4 植被在11 Ma业已达到较高水平[50 ] ;新疆塔里木盆地钻孔有机碳δ 13 C显示,C4 植被在5.3 Ma显著增加[63 ] .这也得到日本海钻孔正构烷烃δ 13 C和内蒙古哺乳动物牙釉化石δ 13 C结果的支持[14 ,64 ] .近来,Lu等[15 ] 汇总全球记录后提出,早上新世存在一次全球性C4 扩张事件.然而,澳大利亚近海钻孔正构烷烃δ 13 C表明,C4 植被直至3.5 Ma才开始扩张[65 ] .同时,太平洋近赤道地区海洋钻孔长链脂肪酸δ 13 C[66 ] 和黄土高原西部临夏盆地牙釉δ 13 C[18 -19 ] 指示,C4 植被直至早更新世后才显著增加.此外,黄土高原东部灵台、蓝田剖面无机碳δ 13 C指示,C4 植被经历了2.9~2.7 Ma、1.3~0.9 Ma、0.6 Ma三次反复扩张[67 ] .基于全球各地C4 植被演化差异,学者们将控制C4 植被演化的因素归结于干旱程度[14 ,63 ,68 -69 ] 、暖季温度和降水[67 ,70 -71 ] 和野火[72 -73 ] 等区域环境因素.由此可见,关于晚中新世以来C4 植被演化的时间和机制等的认识仍未统一. ...
Effect of aridification on carbon isotopic variation and ecologic evolution at 5.3 Ma in the Asian interior
2
2013
... C4 植被的出现可以追溯到32~30 Ma,其大幅扩张一般认为发生在晚中新世[57 ] .来自巴基斯坦Siwalik沉积有机质和无机碳[58 ] 、北美哺乳动物牙釉化石[59 ] 、南美东部钻孔无机碳和动物化石[60 ] 和东非钻孔中动物化石[61 ] 等研究结果均支持这一说法:晚中新世7~8 Ma左右全球发生C4 植被的显著扩张.因此,C4 植被扩张是由大气CO2 浓度降低驱动的全球生态事件[62 ] .21世纪以来,越来越多的记录表明,全球C4 植被扩张的时间和范围并不一致:非洲东北部正构烷烃δ 13 C表明,C4 植被在11 Ma业已达到较高水平[50 ] ;新疆塔里木盆地钻孔有机碳δ 13 C显示,C4 植被在5.3 Ma显著增加[63 ] .这也得到日本海钻孔正构烷烃δ 13 C和内蒙古哺乳动物牙釉化石δ 13 C结果的支持[14 ,64 ] .近来,Lu等[15 ] 汇总全球记录后提出,早上新世存在一次全球性C4 扩张事件.然而,澳大利亚近海钻孔正构烷烃δ 13 C表明,C4 植被直至3.5 Ma才开始扩张[65 ] .同时,太平洋近赤道地区海洋钻孔长链脂肪酸δ 13 C[66 ] 和黄土高原西部临夏盆地牙釉δ 13 C[18 -19 ] 指示,C4 植被直至早更新世后才显著增加.此外,黄土高原东部灵台、蓝田剖面无机碳δ 13 C指示,C4 植被经历了2.9~2.7 Ma、1.3~0.9 Ma、0.6 Ma三次反复扩张[67 ] .基于全球各地C4 植被演化差异,学者们将控制C4 植被演化的因素归结于干旱程度[14 ,63 ,68 -69 ] 、暖季温度和降水[67 ,70 -71 ] 和野火[72 -73 ] 等区域环境因素.由此可见,关于晚中新世以来C4 植被演化的时间和机制等的认识仍未统一. ...
... ,63 ,68 -69 ]、暖季温度和降水[67 ,70 -71 ] 和野火[72 -73 ] 等区域环境因素.由此可见,关于晚中新世以来C4 植被演化的时间和机制等的认识仍未统一. ...
C4 expansion in the central Inner Mongolia during the latest Miocene and early Pliocene
1
2009
... C4 植被的出现可以追溯到32~30 Ma,其大幅扩张一般认为发生在晚中新世[57 ] .来自巴基斯坦Siwalik沉积有机质和无机碳[58 ] 、北美哺乳动物牙釉化石[59 ] 、南美东部钻孔无机碳和动物化石[60 ] 和东非钻孔中动物化石[61 ] 等研究结果均支持这一说法:晚中新世7~8 Ma左右全球发生C4 植被的显著扩张.因此,C4 植被扩张是由大气CO2 浓度降低驱动的全球生态事件[62 ] .21世纪以来,越来越多的记录表明,全球C4 植被扩张的时间和范围并不一致:非洲东北部正构烷烃δ 13 C表明,C4 植被在11 Ma业已达到较高水平[50 ] ;新疆塔里木盆地钻孔有机碳δ 13 C显示,C4 植被在5.3 Ma显著增加[63 ] .这也得到日本海钻孔正构烷烃δ 13 C和内蒙古哺乳动物牙釉化石δ 13 C结果的支持[14 ,64 ] .近来,Lu等[15 ] 汇总全球记录后提出,早上新世存在一次全球性C4 扩张事件.然而,澳大利亚近海钻孔正构烷烃δ 13 C表明,C4 植被直至3.5 Ma才开始扩张[65 ] .同时,太平洋近赤道地区海洋钻孔长链脂肪酸δ 13 C[66 ] 和黄土高原西部临夏盆地牙釉δ 13 C[18 -19 ] 指示,C4 植被直至早更新世后才显著增加.此外,黄土高原东部灵台、蓝田剖面无机碳δ 13 C指示,C4 植被经历了2.9~2.7 Ma、1.3~0.9 Ma、0.6 Ma三次反复扩张[67 ] .基于全球各地C4 植被演化差异,学者们将控制C4 植被演化的因素归结于干旱程度[14 ,63 ,68 -69 ] 、暖季温度和降水[67 ,70 -71 ] 和野火[72 -73 ] 等区域环境因素.由此可见,关于晚中新世以来C4 植被演化的时间和机制等的认识仍未统一. ...
Initial expansion of C4 vegetation in Australia during the Late Pliocene
1
2018
... C4 植被的出现可以追溯到32~30 Ma,其大幅扩张一般认为发生在晚中新世[57 ] .来自巴基斯坦Siwalik沉积有机质和无机碳[58 ] 、北美哺乳动物牙釉化石[59 ] 、南美东部钻孔无机碳和动物化石[60 ] 和东非钻孔中动物化石[61 ] 等研究结果均支持这一说法:晚中新世7~8 Ma左右全球发生C4 植被的显著扩张.因此,C4 植被扩张是由大气CO2 浓度降低驱动的全球生态事件[62 ] .21世纪以来,越来越多的记录表明,全球C4 植被扩张的时间和范围并不一致:非洲东北部正构烷烃δ 13 C表明,C4 植被在11 Ma业已达到较高水平[50 ] ;新疆塔里木盆地钻孔有机碳δ 13 C显示,C4 植被在5.3 Ma显著增加[63 ] .这也得到日本海钻孔正构烷烃δ 13 C和内蒙古哺乳动物牙釉化石δ 13 C结果的支持[14 ,64 ] .近来,Lu等[15 ] 汇总全球记录后提出,早上新世存在一次全球性C4 扩张事件.然而,澳大利亚近海钻孔正构烷烃δ 13 C表明,C4 植被直至3.5 Ma才开始扩张[65 ] .同时,太平洋近赤道地区海洋钻孔长链脂肪酸δ 13 C[66 ] 和黄土高原西部临夏盆地牙釉δ 13 C[18 -19 ] 指示,C4 植被直至早更新世后才显著增加.此外,黄土高原东部灵台、蓝田剖面无机碳δ 13 C指示,C4 植被经历了2.9~2.7 Ma、1.3~0.9 Ma、0.6 Ma三次反复扩张[67 ] .基于全球各地C4 植被演化差异,学者们将控制C4 植被演化的因素归结于干旱程度[14 ,63 ,68 -69 ] 、暖季温度和降水[67 ,70 -71 ] 和野火[72 -73 ] 等区域环境因素.由此可见,关于晚中新世以来C4 植被演化的时间和机制等的认识仍未统一. ...
Stable carbon isotopic variation of long chain n-alkanoic acids in the equatorial Pacific sediments over the last 40 Ma: Implications for expansion of C4 grassland in South America
1
2014
... C4 植被的出现可以追溯到32~30 Ma,其大幅扩张一般认为发生在晚中新世[57 ] .来自巴基斯坦Siwalik沉积有机质和无机碳[58 ] 、北美哺乳动物牙釉化石[59 ] 、南美东部钻孔无机碳和动物化石[60 ] 和东非钻孔中动物化石[61 ] 等研究结果均支持这一说法:晚中新世7~8 Ma左右全球发生C4 植被的显著扩张.因此,C4 植被扩张是由大气CO2 浓度降低驱动的全球生态事件[62 ] .21世纪以来,越来越多的记录表明,全球C4 植被扩张的时间和范围并不一致:非洲东北部正构烷烃δ 13 C表明,C4 植被在11 Ma业已达到较高水平[50 ] ;新疆塔里木盆地钻孔有机碳δ 13 C显示,C4 植被在5.3 Ma显著增加[63 ] .这也得到日本海钻孔正构烷烃δ 13 C和内蒙古哺乳动物牙釉化石δ 13 C结果的支持[14 ,64 ] .近来,Lu等[15 ] 汇总全球记录后提出,早上新世存在一次全球性C4 扩张事件.然而,澳大利亚近海钻孔正构烷烃δ 13 C表明,C4 植被直至3.5 Ma才开始扩张[65 ] .同时,太平洋近赤道地区海洋钻孔长链脂肪酸δ 13 C[66 ] 和黄土高原西部临夏盆地牙釉δ 13 C[18 -19 ] 指示,C4 植被直至早更新世后才显著增加.此外,黄土高原东部灵台、蓝田剖面无机碳δ 13 C指示,C4 植被经历了2.9~2.7 Ma、1.3~0.9 Ma、0.6 Ma三次反复扩张[67 ] .基于全球各地C4 植被演化差异,学者们将控制C4 植被演化的因素归结于干旱程度[14 ,63 ,68 -69 ] 、暖季温度和降水[67 ,70 -71 ] 和野火[72 -73 ] 等区域环境因素.由此可见,关于晚中新世以来C4 植被演化的时间和机制等的认识仍未统一. ...
Multiple expansions of C4 plant biomass in East Asia since 7 Ma coupled with strengthened monsoon circulation
2
2005
... C4 植被的出现可以追溯到32~30 Ma,其大幅扩张一般认为发生在晚中新世[57 ] .来自巴基斯坦Siwalik沉积有机质和无机碳[58 ] 、北美哺乳动物牙釉化石[59 ] 、南美东部钻孔无机碳和动物化石[60 ] 和东非钻孔中动物化石[61 ] 等研究结果均支持这一说法:晚中新世7~8 Ma左右全球发生C4 植被的显著扩张.因此,C4 植被扩张是由大气CO2 浓度降低驱动的全球生态事件[62 ] .21世纪以来,越来越多的记录表明,全球C4 植被扩张的时间和范围并不一致:非洲东北部正构烷烃δ 13 C表明,C4 植被在11 Ma业已达到较高水平[50 ] ;新疆塔里木盆地钻孔有机碳δ 13 C显示,C4 植被在5.3 Ma显著增加[63 ] .这也得到日本海钻孔正构烷烃δ 13 C和内蒙古哺乳动物牙釉化石δ 13 C结果的支持[14 ,64 ] .近来,Lu等[15 ] 汇总全球记录后提出,早上新世存在一次全球性C4 扩张事件.然而,澳大利亚近海钻孔正构烷烃δ 13 C表明,C4 植被直至3.5 Ma才开始扩张[65 ] .同时,太平洋近赤道地区海洋钻孔长链脂肪酸δ 13 C[66 ] 和黄土高原西部临夏盆地牙釉δ 13 C[18 -19 ] 指示,C4 植被直至早更新世后才显著增加.此外,黄土高原东部灵台、蓝田剖面无机碳δ 13 C指示,C4 植被经历了2.9~2.7 Ma、1.3~0.9 Ma、0.6 Ma三次反复扩张[67 ] .基于全球各地C4 植被演化差异,学者们将控制C4 植被演化的因素归结于干旱程度[14 ,63 ,68 -69 ] 、暖季温度和降水[67 ,70 -71 ] 和野火[72 -73 ] 等区域环境因素.由此可见,关于晚中新世以来C4 植被演化的时间和机制等的认识仍未统一. ...
... [67 ,70 -71 ]和野火[72 -73 ] 等区域环境因素.由此可见,关于晚中新世以来C4 植被演化的时间和机制等的认识仍未统一. ...
Carbon isotopic records in paleosols over the Pliocene in Northern China: implication on vegetation development and Tibetan uplift
1
2002
... C4 植被的出现可以追溯到32~30 Ma,其大幅扩张一般认为发生在晚中新世[57 ] .来自巴基斯坦Siwalik沉积有机质和无机碳[58 ] 、北美哺乳动物牙釉化石[59 ] 、南美东部钻孔无机碳和动物化石[60 ] 和东非钻孔中动物化石[61 ] 等研究结果均支持这一说法:晚中新世7~8 Ma左右全球发生C4 植被的显著扩张.因此,C4 植被扩张是由大气CO2 浓度降低驱动的全球生态事件[62 ] .21世纪以来,越来越多的记录表明,全球C4 植被扩张的时间和范围并不一致:非洲东北部正构烷烃δ 13 C表明,C4 植被在11 Ma业已达到较高水平[50 ] ;新疆塔里木盆地钻孔有机碳δ 13 C显示,C4 植被在5.3 Ma显著增加[63 ] .这也得到日本海钻孔正构烷烃δ 13 C和内蒙古哺乳动物牙釉化石δ 13 C结果的支持[14 ,64 ] .近来,Lu等[15 ] 汇总全球记录后提出,早上新世存在一次全球性C4 扩张事件.然而,澳大利亚近海钻孔正构烷烃δ 13 C表明,C4 植被直至3.5 Ma才开始扩张[65 ] .同时,太平洋近赤道地区海洋钻孔长链脂肪酸δ 13 C[66 ] 和黄土高原西部临夏盆地牙釉δ 13 C[18 -19 ] 指示,C4 植被直至早更新世后才显著增加.此外,黄土高原东部灵台、蓝田剖面无机碳δ 13 C指示,C4 植被经历了2.9~2.7 Ma、1.3~0.9 Ma、0.6 Ma三次反复扩张[67 ] .基于全球各地C4 植被演化差异,学者们将控制C4 植被演化的因素归结于干旱程度[14 ,63 ,68 -69 ] 、暖季温度和降水[67 ,70 -71 ] 和野火[72 -73 ] 等区域环境因素.由此可见,关于晚中新世以来C4 植被演化的时间和机制等的认识仍未统一. ...
Miocene to Pliocene changes in South African hydrology and vegetation in relation to the expansion of C4 plants
1
2013
... C4 植被的出现可以追溯到32~30 Ma,其大幅扩张一般认为发生在晚中新世[57 ] .来自巴基斯坦Siwalik沉积有机质和无机碳[58 ] 、北美哺乳动物牙釉化石[59 ] 、南美东部钻孔无机碳和动物化石[60 ] 和东非钻孔中动物化石[61 ] 等研究结果均支持这一说法:晚中新世7~8 Ma左右全球发生C4 植被的显著扩张.因此,C4 植被扩张是由大气CO2 浓度降低驱动的全球生态事件[62 ] .21世纪以来,越来越多的记录表明,全球C4 植被扩张的时间和范围并不一致:非洲东北部正构烷烃δ 13 C表明,C4 植被在11 Ma业已达到较高水平[50 ] ;新疆塔里木盆地钻孔有机碳δ 13 C显示,C4 植被在5.3 Ma显著增加[63 ] .这也得到日本海钻孔正构烷烃δ 13 C和内蒙古哺乳动物牙釉化石δ 13 C结果的支持[14 ,64 ] .近来,Lu等[15 ] 汇总全球记录后提出,早上新世存在一次全球性C4 扩张事件.然而,澳大利亚近海钻孔正构烷烃δ 13 C表明,C4 植被直至3.5 Ma才开始扩张[65 ] .同时,太平洋近赤道地区海洋钻孔长链脂肪酸δ 13 C[66 ] 和黄土高原西部临夏盆地牙釉δ 13 C[18 -19 ] 指示,C4 植被直至早更新世后才显著增加.此外,黄土高原东部灵台、蓝田剖面无机碳δ 13 C指示,C4 植被经历了2.9~2.7 Ma、1.3~0.9 Ma、0.6 Ma三次反复扩张[67 ] .基于全球各地C4 植被演化差异,学者们将控制C4 植被演化的因素归结于干旱程度[14 ,63 ,68 -69 ] 、暖季温度和降水[67 ,70 -71 ] 和野火[72 -73 ] 等区域环境因素.由此可见,关于晚中新世以来C4 植被演化的时间和机制等的认识仍未统一. ...
Climate change as the dominant control on glacial-interglacial variations in C3 and C4 plant abundance
1
2001
... C4 植被的出现可以追溯到32~30 Ma,其大幅扩张一般认为发生在晚中新世[57 ] .来自巴基斯坦Siwalik沉积有机质和无机碳[58 ] 、北美哺乳动物牙釉化石[59 ] 、南美东部钻孔无机碳和动物化石[60 ] 和东非钻孔中动物化石[61 ] 等研究结果均支持这一说法:晚中新世7~8 Ma左右全球发生C4 植被的显著扩张.因此,C4 植被扩张是由大气CO2 浓度降低驱动的全球生态事件[62 ] .21世纪以来,越来越多的记录表明,全球C4 植被扩张的时间和范围并不一致:非洲东北部正构烷烃δ 13 C表明,C4 植被在11 Ma业已达到较高水平[50 ] ;新疆塔里木盆地钻孔有机碳δ 13 C显示,C4 植被在5.3 Ma显著增加[63 ] .这也得到日本海钻孔正构烷烃δ 13 C和内蒙古哺乳动物牙釉化石δ 13 C结果的支持[14 ,64 ] .近来,Lu等[15 ] 汇总全球记录后提出,早上新世存在一次全球性C4 扩张事件.然而,澳大利亚近海钻孔正构烷烃δ 13 C表明,C4 植被直至3.5 Ma才开始扩张[65 ] .同时,太平洋近赤道地区海洋钻孔长链脂肪酸δ 13 C[66 ] 和黄土高原西部临夏盆地牙釉δ 13 C[18 -19 ] 指示,C4 植被直至早更新世后才显著增加.此外,黄土高原东部灵台、蓝田剖面无机碳δ 13 C指示,C4 植被经历了2.9~2.7 Ma、1.3~0.9 Ma、0.6 Ma三次反复扩张[67 ] .基于全球各地C4 植被演化差异,学者们将控制C4 植被演化的因素归结于干旱程度[14 ,63 ,68 -69 ] 、暖季温度和降水[67 ,70 -71 ] 和野火[72 -73 ] 等区域环境因素.由此可见,关于晚中新世以来C4 植被演化的时间和机制等的认识仍未统一. ...
Negative δ 18 O–δ 13 C relationship of pedogenic carbonate from northern China indicates a strong response of C3 /C4 biomass to the seasonality of Asian monsoon precipitation
1
2012
... C4 植被的出现可以追溯到32~30 Ma,其大幅扩张一般认为发生在晚中新世[57 ] .来自巴基斯坦Siwalik沉积有机质和无机碳[58 ] 、北美哺乳动物牙釉化石[59 ] 、南美东部钻孔无机碳和动物化石[60 ] 和东非钻孔中动物化石[61 ] 等研究结果均支持这一说法:晚中新世7~8 Ma左右全球发生C4 植被的显著扩张.因此,C4 植被扩张是由大气CO2 浓度降低驱动的全球生态事件[62 ] .21世纪以来,越来越多的记录表明,全球C4 植被扩张的时间和范围并不一致:非洲东北部正构烷烃δ 13 C表明,C4 植被在11 Ma业已达到较高水平[50 ] ;新疆塔里木盆地钻孔有机碳δ 13 C显示,C4 植被在5.3 Ma显著增加[63 ] .这也得到日本海钻孔正构烷烃δ 13 C和内蒙古哺乳动物牙釉化石δ 13 C结果的支持[14 ,64 ] .近来,Lu等[15 ] 汇总全球记录后提出,早上新世存在一次全球性C4 扩张事件.然而,澳大利亚近海钻孔正构烷烃δ 13 C表明,C4 植被直至3.5 Ma才开始扩张[65 ] .同时,太平洋近赤道地区海洋钻孔长链脂肪酸δ 13 C[66 ] 和黄土高原西部临夏盆地牙釉δ 13 C[18 -19 ] 指示,C4 植被直至早更新世后才显著增加.此外,黄土高原东部灵台、蓝田剖面无机碳δ 13 C指示,C4 植被经历了2.9~2.7 Ma、1.3~0.9 Ma、0.6 Ma三次反复扩张[67 ] .基于全球各地C4 植被演化差异,学者们将控制C4 植被演化的因素归结于干旱程度[14 ,63 ,68 -69 ] 、暖季温度和降水[67 ,70 -71 ] 和野火[72 -73 ] 等区域环境因素.由此可见,关于晚中新世以来C4 植被演化的时间和机制等的认识仍未统一. ...
Late Miocene atmospheric CO2 concentrations and the expansion of C4 grasses
1
1999
... C4 植被的出现可以追溯到32~30 Ma,其大幅扩张一般认为发生在晚中新世[57 ] .来自巴基斯坦Siwalik沉积有机质和无机碳[58 ] 、北美哺乳动物牙釉化石[59 ] 、南美东部钻孔无机碳和动物化石[60 ] 和东非钻孔中动物化石[61 ] 等研究结果均支持这一说法:晚中新世7~8 Ma左右全球发生C4 植被的显著扩张.因此,C4 植被扩张是由大气CO2 浓度降低驱动的全球生态事件[62 ] .21世纪以来,越来越多的记录表明,全球C4 植被扩张的时间和范围并不一致:非洲东北部正构烷烃δ 13 C表明,C4 植被在11 Ma业已达到较高水平[50 ] ;新疆塔里木盆地钻孔有机碳δ 13 C显示,C4 植被在5.3 Ma显著增加[63 ] .这也得到日本海钻孔正构烷烃δ 13 C和内蒙古哺乳动物牙釉化石δ 13 C结果的支持[14 ,64 ] .近来,Lu等[15 ] 汇总全球记录后提出,早上新世存在一次全球性C4 扩张事件.然而,澳大利亚近海钻孔正构烷烃δ 13 C表明,C4 植被直至3.5 Ma才开始扩张[65 ] .同时,太平洋近赤道地区海洋钻孔长链脂肪酸δ 13 C[66 ] 和黄土高原西部临夏盆地牙釉δ 13 C[18 -19 ] 指示,C4 植被直至早更新世后才显著增加.此外,黄土高原东部灵台、蓝田剖面无机碳δ 13 C指示,C4 植被经历了2.9~2.7 Ma、1.3~0.9 Ma、0.6 Ma三次反复扩张[67 ] .基于全球各地C4 植被演化差异,学者们将控制C4 植被演化的因素归结于干旱程度[14 ,63 ,68 -69 ] 、暖季温度和降水[67 ,70 -71 ] 和野火[72 -73 ] 等区域环境因素.由此可见,关于晚中新世以来C4 植被演化的时间和机制等的认识仍未统一. ...
The role of fire in Miocene to Pliocene C4 grassland and ecosystem evolution
1
2013
... C4 植被的出现可以追溯到32~30 Ma,其大幅扩张一般认为发生在晚中新世[57 ] .来自巴基斯坦Siwalik沉积有机质和无机碳[58 ] 、北美哺乳动物牙釉化石[59 ] 、南美东部钻孔无机碳和动物化石[60 ] 和东非钻孔中动物化石[61 ] 等研究结果均支持这一说法:晚中新世7~8 Ma左右全球发生C4 植被的显著扩张.因此,C4 植被扩张是由大气CO2 浓度降低驱动的全球生态事件[62 ] .21世纪以来,越来越多的记录表明,全球C4 植被扩张的时间和范围并不一致:非洲东北部正构烷烃δ 13 C表明,C4 植被在11 Ma业已达到较高水平[50 ] ;新疆塔里木盆地钻孔有机碳δ 13 C显示,C4 植被在5.3 Ma显著增加[63 ] .这也得到日本海钻孔正构烷烃δ 13 C和内蒙古哺乳动物牙釉化石δ 13 C结果的支持[14 ,64 ] .近来,Lu等[15 ] 汇总全球记录后提出,早上新世存在一次全球性C4 扩张事件.然而,澳大利亚近海钻孔正构烷烃δ 13 C表明,C4 植被直至3.5 Ma才开始扩张[65 ] .同时,太平洋近赤道地区海洋钻孔长链脂肪酸δ 13 C[66 ] 和黄土高原西部临夏盆地牙釉δ 13 C[18 -19 ] 指示,C4 植被直至早更新世后才显著增加.此外,黄土高原东部灵台、蓝田剖面无机碳δ 13 C指示,C4 植被经历了2.9~2.7 Ma、1.3~0.9 Ma、0.6 Ma三次反复扩张[67 ] .基于全球各地C4 植被演化差异,学者们将控制C4 植被演化的因素归结于干旱程度[14 ,63 ,68 -69 ] 、暖季温度和降水[67 ,70 -71 ] 和野火[72 -73 ] 等区域环境因素.由此可见,关于晚中新世以来C4 植被演化的时间和机制等的认识仍未统一. ...


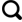



 甘公网安备 62010202000676号
甘公网安备 62010202000676号
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}