1
2016
... 国际冻土协会将多年冻土定义为温度在0 ℃或低于0 ℃至少连续存在两年的岩土层[1].在高纬度地区,沼泽湿地和冻土通常共生[2].多年冻土土壤有机氮经过矿化作用转换为铵态氮(NH4+-N)和硝态氮(NO3--N),通过硝化和反硝化作用产生温室气体氧化亚氮(N2O)逸散到大气中,从而对湿地生态系统和全球气候变化产生影响[3].高纬度多年冻土泥炭地土壤氮素储存量约40~60 Pg[4],在气候变暖的背景下,冻土退化、冻融循环过程的改变影响了土壤氮素的矿化过程[5],提高了冻土泥炭地N2O排放潜能[6],从而加剧全球变暖. ...
1
2016
... 国际冻土协会将多年冻土定义为温度在0 ℃或低于0 ℃至少连续存在两年的岩土层[1].在高纬度地区,沼泽湿地和冻土通常共生[2].多年冻土土壤有机氮经过矿化作用转换为铵态氮(NH4+-N)和硝态氮(NO3--N),通过硝化和反硝化作用产生温室气体氧化亚氮(N2O)逸散到大气中,从而对湿地生态系统和全球气候变化产生影响[3].高纬度多年冻土泥炭地土壤氮素储存量约40~60 Pg[4],在气候变暖的背景下,冻土退化、冻融循环过程的改变影响了土壤氮素的矿化过程[5],提高了冻土泥炭地N2O排放潜能[6],从而加剧全球变暖. ...
Discussion on the symbiotic mechanisms of swamp with permafrost: taking Da-Xiao Hinggan Mountains as examples
1
2000
... 国际冻土协会将多年冻土定义为温度在0 ℃或低于0 ℃至少连续存在两年的岩土层[1].在高纬度地区,沼泽湿地和冻土通常共生[2].多年冻土土壤有机氮经过矿化作用转换为铵态氮(NH4+-N)和硝态氮(NO3--N),通过硝化和反硝化作用产生温室气体氧化亚氮(N2O)逸散到大气中,从而对湿地生态系统和全球气候变化产生影响[3].高纬度多年冻土泥炭地土壤氮素储存量约40~60 Pg[4],在气候变暖的背景下,冻土退化、冻融循环过程的改变影响了土壤氮素的矿化过程[5],提高了冻土泥炭地N2O排放潜能[6],从而加剧全球变暖. ...
试论沼泽与冻土的共生机理——以中国大小兴安岭地区为例
1
2000
... 国际冻土协会将多年冻土定义为温度在0 ℃或低于0 ℃至少连续存在两年的岩土层[1].在高纬度地区,沼泽湿地和冻土通常共生[2].多年冻土土壤有机氮经过矿化作用转换为铵态氮(NH4+-N)和硝态氮(NO3--N),通过硝化和反硝化作用产生温室气体氧化亚氮(N2O)逸散到大气中,从而对湿地生态系统和全球气候变化产生影响[3].高纬度多年冻土泥炭地土壤氮素储存量约40~60 Pg[4],在气候变暖的背景下,冻土退化、冻融循环过程的改变影响了土壤氮素的矿化过程[5],提高了冻土泥炭地N2O排放潜能[6],从而加剧全球变暖. ...
Effects of temperature and nitrogen input on nitrogen mineralization of soils in the newly created marshes of the Yellow River estuary
1
2020
... 国际冻土协会将多年冻土定义为温度在0 ℃或低于0 ℃至少连续存在两年的岩土层[1].在高纬度地区,沼泽湿地和冻土通常共生[2].多年冻土土壤有机氮经过矿化作用转换为铵态氮(NH4+-N)和硝态氮(NO3--N),通过硝化和反硝化作用产生温室气体氧化亚氮(N2O)逸散到大气中,从而对湿地生态系统和全球气候变化产生影响[3].高纬度多年冻土泥炭地土壤氮素储存量约40~60 Pg[4],在气候变暖的背景下,冻土退化、冻融循环过程的改变影响了土壤氮素的矿化过程[5],提高了冻土泥炭地N2O排放潜能[6],从而加剧全球变暖. ...
黄河口碱蓬湿地土壤氮矿化特征对温度及氮输入的响应
1
2020
... 国际冻土协会将多年冻土定义为温度在0 ℃或低于0 ℃至少连续存在两年的岩土层[1].在高纬度地区,沼泽湿地和冻土通常共生[2].多年冻土土壤有机氮经过矿化作用转换为铵态氮(NH4+-N)和硝态氮(NO3--N),通过硝化和反硝化作用产生温室气体氧化亚氮(N2O)逸散到大气中,从而对湿地生态系统和全球气候变化产生影响[3].高纬度多年冻土泥炭地土壤氮素储存量约40~60 Pg[4],在气候变暖的背景下,冻土退化、冻融循环过程的改变影响了土壤氮素的矿化过程[5],提高了冻土泥炭地N2O排放潜能[6],从而加剧全球变暖. ...
Field information links permafrost carbon to physical vulnerabilities of thawing
1
2012
... 国际冻土协会将多年冻土定义为温度在0 ℃或低于0 ℃至少连续存在两年的岩土层[1].在高纬度地区,沼泽湿地和冻土通常共生[2].多年冻土土壤有机氮经过矿化作用转换为铵态氮(NH4+-N)和硝态氮(NO3--N),通过硝化和反硝化作用产生温室气体氧化亚氮(N2O)逸散到大气中,从而对湿地生态系统和全球气候变化产生影响[3].高纬度多年冻土泥炭地土壤氮素储存量约40~60 Pg[4],在气候变暖的背景下,冻土退化、冻融循环过程的改变影响了土壤氮素的矿化过程[5],提高了冻土泥炭地N2O排放潜能[6],从而加剧全球变暖. ...
Preliminary assessment of permafrost degradation and its trend in the Great and Small Xing’ an Mountains
1
2006
... 国际冻土协会将多年冻土定义为温度在0 ℃或低于0 ℃至少连续存在两年的岩土层[1].在高纬度地区,沼泽湿地和冻土通常共生[2].多年冻土土壤有机氮经过矿化作用转换为铵态氮(NH4+-N)和硝态氮(NO3--N),通过硝化和反硝化作用产生温室气体氧化亚氮(N2O)逸散到大气中,从而对湿地生态系统和全球气候变化产生影响[3].高纬度多年冻土泥炭地土壤氮素储存量约40~60 Pg[4],在气候变暖的背景下,冻土退化、冻融循环过程的改变影响了土壤氮素的矿化过程[5],提高了冻土泥炭地N2O排放潜能[6],从而加剧全球变暖. ...
大小兴安岭多年冻土退化及其趋势初步评估
1
2006
... 国际冻土协会将多年冻土定义为温度在0 ℃或低于0 ℃至少连续存在两年的岩土层[1].在高纬度地区,沼泽湿地和冻土通常共生[2].多年冻土土壤有机氮经过矿化作用转换为铵态氮(NH4+-N)和硝态氮(NO3--N),通过硝化和反硝化作用产生温室气体氧化亚氮(N2O)逸散到大气中,从而对湿地生态系统和全球气候变化产生影响[3].高纬度多年冻土泥炭地土壤氮素储存量约40~60 Pg[4],在气候变暖的背景下,冻土退化、冻融循环过程的改变影响了土壤氮素的矿化过程[5],提高了冻土泥炭地N2O排放潜能[6],从而加剧全球变暖. ...
Hot spots for nitrous oxide emissions found in different types of permafrost peatlands
1
2011
... 国际冻土协会将多年冻土定义为温度在0 ℃或低于0 ℃至少连续存在两年的岩土层[1].在高纬度地区,沼泽湿地和冻土通常共生[2].多年冻土土壤有机氮经过矿化作用转换为铵态氮(NH4+-N)和硝态氮(NO3--N),通过硝化和反硝化作用产生温室气体氧化亚氮(N2O)逸散到大气中,从而对湿地生态系统和全球气候变化产生影响[3].高纬度多年冻土泥炭地土壤氮素储存量约40~60 Pg[4],在气候变暖的背景下,冻土退化、冻融循环过程的改变影响了土壤氮素的矿化过程[5],提高了冻土泥炭地N2O排放潜能[6],从而加剧全球变暖. ...
Relationships of soil physical and microbial properties with nitrous oxide emission under effects of freezing-thawing cycles
1
2007
... 冻融作用是土壤由于气温变化而反复冻结融化产生的[7],是土壤水分和热量动态变化的表现形式[8].冻融过程直接改变了土壤的温度[9]、水分迁移[10]、微生物活性[11],从而加速有效氮素以气体形式或淋溶形式释放[12].此过程主要发生在植物非生长的春季和秋季[13-14].春季和秋季气温仍较低,一般认为气温较低时有机质分解较弱,然而,研究发现秋季土壤氮矿化仍非常强烈[15];秋季土壤微生物活性和营养物质含量发生变化[16],导致氮矿化过程不断累积.此外,冻融作用在非生长季产生的影响可能延续到冻融后期[17],秋季氮矿化的累积会导致春季冻融期无机氮的释放和N2O排放潜能. ...
冻融作用下土壤物理和微生物性状变化与氧化亚氮排放的关系
1
2007
... 冻融作用是土壤由于气温变化而反复冻结融化产生的[7],是土壤水分和热量动态变化的表现形式[8].冻融过程直接改变了土壤的温度[9]、水分迁移[10]、微生物活性[11],从而加速有效氮素以气体形式或淋溶形式释放[12].此过程主要发生在植物非生长的春季和秋季[13-14].春季和秋季气温仍较低,一般认为气温较低时有机质分解较弱,然而,研究发现秋季土壤氮矿化仍非常强烈[15];秋季土壤微生物活性和营养物质含量发生变化[16],导致氮矿化过程不断累积.此外,冻融作用在非生长季产生的影响可能延续到冻融后期[17],秋季氮矿化的累积会导致春季冻融期无机氮的释放和N2O排放潜能. ...
Effects of freeze-thaw on soil nitrogen transformation and n2o emission: a review
1
2013
... 冻融作用是土壤由于气温变化而反复冻结融化产生的[7],是土壤水分和热量动态变化的表现形式[8].冻融过程直接改变了土壤的温度[9]、水分迁移[10]、微生物活性[11],从而加速有效氮素以气体形式或淋溶形式释放[12].此过程主要发生在植物非生长的春季和秋季[13-14].春季和秋季气温仍较低,一般认为气温较低时有机质分解较弱,然而,研究发现秋季土壤氮矿化仍非常强烈[15];秋季土壤微生物活性和营养物质含量发生变化[16],导致氮矿化过程不断累积.此外,冻融作用在非生长季产生的影响可能延续到冻融后期[17],秋季氮矿化的累积会导致春季冻融期无机氮的释放和N2O排放潜能. ...
冻融对土壤氮素转化和N2O排放的影响研究进展
1
2013
... 冻融作用是土壤由于气温变化而反复冻结融化产生的[7],是土壤水分和热量动态变化的表现形式[8].冻融过程直接改变了土壤的温度[9]、水分迁移[10]、微生物活性[11],从而加速有效氮素以气体形式或淋溶形式释放[12].此过程主要发生在植物非生长的春季和秋季[13-14].春季和秋季气温仍较低,一般认为气温较低时有机质分解较弱,然而,研究发现秋季土壤氮矿化仍非常强烈[15];秋季土壤微生物活性和营养物质含量发生变化[16],导致氮矿化过程不断累积.此外,冻融作用在非生长季产生的影响可能延续到冻融后期[17],秋季氮矿化的累积会导致春季冻融期无机氮的释放和N2O排放潜能. ...
Spatiotemporal variation characteristics of soil water content and temperature within active layer at slope scale in the Fenghuoshan Basin, Tibetan Plateau
1
2020
... 冻融作用是土壤由于气温变化而反复冻结融化产生的[7],是土壤水分和热量动态变化的表现形式[8].冻融过程直接改变了土壤的温度[9]、水分迁移[10]、微生物活性[11],从而加速有效氮素以气体形式或淋溶形式释放[12].此过程主要发生在植物非生长的春季和秋季[13-14].春季和秋季气温仍较低,一般认为气温较低时有机质分解较弱,然而,研究发现秋季土壤氮矿化仍非常强烈[15];秋季土壤微生物活性和营养物质含量发生变化[16],导致氮矿化过程不断累积.此外,冻融作用在非生长季产生的影响可能延续到冻融后期[17],秋季氮矿化的累积会导致春季冻融期无机氮的释放和N2O排放潜能. ...
青藏高原风火山流域坡面尺度活动层土壤水热时空变化特征
1
2020
... 冻融作用是土壤由于气温变化而反复冻结融化产生的[7],是土壤水分和热量动态变化的表现形式[8].冻融过程直接改变了土壤的温度[9]、水分迁移[10]、微生物活性[11],从而加速有效氮素以气体形式或淋溶形式释放[12].此过程主要发生在植物非生长的春季和秋季[13-14].春季和秋季气温仍较低,一般认为气温较低时有机质分解较弱,然而,研究发现秋季土壤氮矿化仍非常强烈[15];秋季土壤微生物活性和营养物质含量发生变化[16],导致氮矿化过程不断累积.此外,冻融作用在非生长季产生的影响可能延续到冻融后期[17],秋季氮矿化的累积会导致春季冻融期无机氮的释放和N2O排放潜能. ...
Characteristics of hydro-thermal coupling during soil freezing-thawing process in seasonally frozen soil regions on the Tibetan Plateau
1
2020
... 冻融作用是土壤由于气温变化而反复冻结融化产生的[7],是土壤水分和热量动态变化的表现形式[8].冻融过程直接改变了土壤的温度[9]、水分迁移[10]、微生物活性[11],从而加速有效氮素以气体形式或淋溶形式释放[12].此过程主要发生在植物非生长的春季和秋季[13-14].春季和秋季气温仍较低,一般认为气温较低时有机质分解较弱,然而,研究发现秋季土壤氮矿化仍非常强烈[15];秋季土壤微生物活性和营养物质含量发生变化[16],导致氮矿化过程不断累积.此外,冻融作用在非生长季产生的影响可能延续到冻融后期[17],秋季氮矿化的累积会导致春季冻融期无机氮的释放和N2O排放潜能. ...
青藏高原季节冻土区土壤冻融过程水热耦合特征
1
2020
... 冻融作用是土壤由于气温变化而反复冻结融化产生的[7],是土壤水分和热量动态变化的表现形式[8].冻融过程直接改变了土壤的温度[9]、水分迁移[10]、微生物活性[11],从而加速有效氮素以气体形式或淋溶形式释放[12].此过程主要发生在植物非生长的春季和秋季[13-14].春季和秋季气温仍较低,一般认为气温较低时有机质分解较弱,然而,研究发现秋季土壤氮矿化仍非常强烈[15];秋季土壤微生物活性和营养物质含量发生变化[16],导致氮矿化过程不断累积.此外,冻融作用在非生长季产生的影响可能延续到冻融后期[17],秋季氮矿化的累积会导致春季冻融期无机氮的释放和N2O排放潜能. ...
Effects of freeze-thaw and soil structure on nitrous oxide produced in a clay soil
1
2000
... 冻融作用是土壤由于气温变化而反复冻结融化产生的[7],是土壤水分和热量动态变化的表现形式[8].冻融过程直接改变了土壤的温度[9]、水分迁移[10]、微生物活性[11],从而加速有效氮素以气体形式或淋溶形式释放[12].此过程主要发生在植物非生长的春季和秋季[13-14].春季和秋季气温仍较低,一般认为气温较低时有机质分解较弱,然而,研究发现秋季土壤氮矿化仍非常强烈[15];秋季土壤微生物活性和营养物质含量发生变化[16],导致氮矿化过程不断累积.此外,冻融作用在非生长季产生的影响可能延续到冻融后期[17],秋季氮矿化的累积会导致春季冻融期无机氮的释放和N2O排放潜能. ...
Effects of experimental freezing on soil nitrogen dynamics in soils from a net nitrification gradient in a nitrogen-saturated hardwood forest ecosystem
1
2010
... 冻融作用是土壤由于气温变化而反复冻结融化产生的[7],是土壤水分和热量动态变化的表现形式[8].冻融过程直接改变了土壤的温度[9]、水分迁移[10]、微生物活性[11],从而加速有效氮素以气体形式或淋溶形式释放[12].此过程主要发生在植物非生长的春季和秋季[13-14].春季和秋季气温仍较低,一般认为气温较低时有机质分解较弱,然而,研究发现秋季土壤氮矿化仍非常强烈[15];秋季土壤微生物活性和营养物质含量发生变化[16],导致氮矿化过程不断累积.此外,冻融作用在非生长季产生的影响可能延续到冻融后期[17],秋季氮矿化的累积会导致春季冻融期无机氮的释放和N2O排放潜能. ...
Effects of hard frost and freeze-thaw cycles on decomposer communities and N mineralisation in boreal forest soil
1
2003
... 冻融作用是土壤由于气温变化而反复冻结融化产生的[7],是土壤水分和热量动态变化的表现形式[8].冻融过程直接改变了土壤的温度[9]、水分迁移[10]、微生物活性[11],从而加速有效氮素以气体形式或淋溶形式释放[12].此过程主要发生在植物非生长的春季和秋季[13-14].春季和秋季气温仍较低,一般认为气温较低时有机质分解较弱,然而,研究发现秋季土壤氮矿化仍非常强烈[15];秋季土壤微生物活性和营养物质含量发生变化[16],导致氮矿化过程不断累积.此外,冻融作用在非生长季产生的影响可能延续到冻融后期[17],秋季氮矿化的累积会导致春季冻融期无机氮的释放和N2O排放潜能. ...
CO2 and N2O emissions from gleyic soils in the Russian tundra and a German forest during freeze-thaw periods: a microcosm study
1
2006
... 冻融作用是土壤由于气温变化而反复冻结融化产生的[7],是土壤水分和热量动态变化的表现形式[8].冻融过程直接改变了土壤的温度[9]、水分迁移[10]、微生物活性[11],从而加速有效氮素以气体形式或淋溶形式释放[12].此过程主要发生在植物非生长的春季和秋季[13-14].春季和秋季气温仍较低,一般认为气温较低时有机质分解较弱,然而,研究发现秋季土壤氮矿化仍非常强烈[15];秋季土壤微生物活性和营养物质含量发生变化[16],导致氮矿化过程不断累积.此外,冻融作用在非生长季产生的影响可能延续到冻融后期[17],秋季氮矿化的累积会导致春季冻融期无机氮的释放和N2O排放潜能. ...
Increased snow depth affects microbial activity and nitrogen mineralization in two Arctic tundra communities
1
2004
... 冻融作用是土壤由于气温变化而反复冻结融化产生的[7],是土壤水分和热量动态变化的表现形式[8].冻融过程直接改变了土壤的温度[9]、水分迁移[10]、微生物活性[11],从而加速有效氮素以气体形式或淋溶形式释放[12].此过程主要发生在植物非生长的春季和秋季[13-14].春季和秋季气温仍较低,一般认为气温较低时有机质分解较弱,然而,研究发现秋季土壤氮矿化仍非常强烈[15];秋季土壤微生物活性和营养物质含量发生变化[16],导致氮矿化过程不断累积.此外,冻融作用在非生长季产生的影响可能延续到冻融后期[17],秋季氮矿化的累积会导致春季冻融期无机氮的释放和N2O排放潜能. ...
Quantitative analysis of dynamic change of land use and its influencing factors in upper reaches of the Heihe River
1
2013
... 冻融作用是土壤由于气温变化而反复冻结融化产生的[7],是土壤水分和热量动态变化的表现形式[8].冻融过程直接改变了土壤的温度[9]、水分迁移[10]、微生物活性[11],从而加速有效氮素以气体形式或淋溶形式释放[12].此过程主要发生在植物非生长的春季和秋季[13-14].春季和秋季气温仍较低,一般认为气温较低时有机质分解较弱,然而,研究发现秋季土壤氮矿化仍非常强烈[15];秋季土壤微生物活性和营养物质含量发生变化[16],导致氮矿化过程不断累积.此外,冻融作用在非生长季产生的影响可能延续到冻融后期[17],秋季氮矿化的累积会导致春季冻融期无机氮的释放和N2O排放潜能. ...
黑河上游土地利用动态变化及影响因素的定量分析
1
2013
... 冻融作用是土壤由于气温变化而反复冻结融化产生的[7],是土壤水分和热量动态变化的表现形式[8].冻融过程直接改变了土壤的温度[9]、水分迁移[10]、微生物活性[11],从而加速有效氮素以气体形式或淋溶形式释放[12].此过程主要发生在植物非生长的春季和秋季[13-14].春季和秋季气温仍较低,一般认为气温较低时有机质分解较弱,然而,研究发现秋季土壤氮矿化仍非常强烈[15];秋季土壤微生物活性和营养物质含量发生变化[16],导致氮矿化过程不断累积.此外,冻融作用在非生长季产生的影响可能延续到冻融后期[17],秋季氮矿化的累积会导致春季冻融期无机氮的释放和N2O排放潜能. ...
Response of soil gross nitrogen mineralization processes to seasonal freeze-thawing in the forests of Tianshan Mountain
1
2020
... 冻融作用是土壤由于气温变化而反复冻结融化产生的[7],是土壤水分和热量动态变化的表现形式[8].冻融过程直接改变了土壤的温度[9]、水分迁移[10]、微生物活性[11],从而加速有效氮素以气体形式或淋溶形式释放[12].此过程主要发生在植物非生长的春季和秋季[13-14].春季和秋季气温仍较低,一般认为气温较低时有机质分解较弱,然而,研究发现秋季土壤氮矿化仍非常强烈[15];秋季土壤微生物活性和营养物质含量发生变化[16],导致氮矿化过程不断累积.此外,冻融作用在非生长季产生的影响可能延续到冻融后期[17],秋季氮矿化的累积会导致春季冻融期无机氮的释放和N2O排放潜能. ...
天山林区土壤总氮矿化过程对季节性冻融的响应
1
2020
... 冻融作用是土壤由于气温变化而反复冻结融化产生的[7],是土壤水分和热量动态变化的表现形式[8].冻融过程直接改变了土壤的温度[9]、水分迁移[10]、微生物活性[11],从而加速有效氮素以气体形式或淋溶形式释放[12].此过程主要发生在植物非生长的春季和秋季[13-14].春季和秋季气温仍较低,一般认为气温较低时有机质分解较弱,然而,研究发现秋季土壤氮矿化仍非常强烈[15];秋季土壤微生物活性和营养物质含量发生变化[16],导致氮矿化过程不断累积.此外,冻融作用在非生长季产生的影响可能延续到冻融后期[17],秋季氮矿化的累积会导致春季冻融期无机氮的释放和N2O排放潜能. ...
A review of affecting factors of soil nitrogen mineralization in forest ecosystems
1
2001
... 目前关于无机氮影响因素的研究主要集中在温度、水分、土壤理化性质、微生物、植被类型等方面[18-22],虽然获得了不少研究成果,但在不同自然生态系统中研究结果还存在较大差异,主要归结于土壤温度和水分.在冻融条件下频繁的水热交替会影响土壤微生物活性,改变土壤团聚体结构和对土壤无机氮固持[23].有研究发现,在一定模拟条件下,土壤水分和温度与土壤氮矿化之间存在线性相关[24];但也有研究发现土壤氮矿化与温度之间并没有直接关系[25].国内外在不同土地类型(湿地[26]、草地[27]、田地[28]、水稻[29])、不同坡面[30]、不同纬度[31-32]等条件下通过室内模拟和原位培养的方法已经开展了无机氮变化影响因子的研究.其中,室内模拟实验容易改变土壤温度和水分条件,可得到土壤无机氮变化规律,但是由于研究方法和冻融格局的差异,得到的结果也不一致[33].与室内模拟冻融实验相比,野外原位实验弥补了温度、水分极端变化的缺陷,很大程度上还原了无机氮在自然条件下的变化规律,但目前野外原位监测研究仍非常少见,导致秋季冻融对土壤无机氮变化的影响尚不明确.我国关于冻融条件下无机氮的研究主要集中在森林[34]、农田[35]、山地[31]、湿地[36]等生态系统,但在多年冻土泥炭地研究较少.因此,在气候变暖、多年冻土退化背景下,深入探讨秋季冻融对多年冻土泥炭地无机氮影响的研究是必要且紧迫的. ...
森林生态系统土壤氮矿化影响因素研究进展
1
2001
... 目前关于无机氮影响因素的研究主要集中在温度、水分、土壤理化性质、微生物、植被类型等方面[18-22],虽然获得了不少研究成果,但在不同自然生态系统中研究结果还存在较大差异,主要归结于土壤温度和水分.在冻融条件下频繁的水热交替会影响土壤微生物活性,改变土壤团聚体结构和对土壤无机氮固持[23].有研究发现,在一定模拟条件下,土壤水分和温度与土壤氮矿化之间存在线性相关[24];但也有研究发现土壤氮矿化与温度之间并没有直接关系[25].国内外在不同土地类型(湿地[26]、草地[27]、田地[28]、水稻[29])、不同坡面[30]、不同纬度[31-32]等条件下通过室内模拟和原位培养的方法已经开展了无机氮变化影响因子的研究.其中,室内模拟实验容易改变土壤温度和水分条件,可得到土壤无机氮变化规律,但是由于研究方法和冻融格局的差异,得到的结果也不一致[33].与室内模拟冻融实验相比,野外原位实验弥补了温度、水分极端变化的缺陷,很大程度上还原了无机氮在自然条件下的变化规律,但目前野外原位监测研究仍非常少见,导致秋季冻融对土壤无机氮变化的影响尚不明确.我国关于冻融条件下无机氮的研究主要集中在森林[34]、农田[35]、山地[31]、湿地[36]等生态系统,但在多年冻土泥炭地研究较少.因此,在气候变暖、多年冻土退化背景下,深入探讨秋季冻融对多年冻土泥炭地无机氮影响的研究是必要且紧迫的. ...
Relative nitrogen mineralization and nitrification in soils of two contrasting hardwood forests: Effects of site microclimate and initial soil chemistry
1987
Net nitrogen mineralization in soils under four forest communities in Hunan Province
2009
Seasonal dynamics of soil mineral nitrogen pools and nitrogen mineralization rate in different forests in subtropical China
2017
亚热带不同林分土壤矿质氮库及氮矿化速率的季节动态
2017
Nitrogen mineralization rate in different soil layers and its influence factors under plastic film mulched in Danjiangkou Reservoir area, China
1
2018
... 目前关于无机氮影响因素的研究主要集中在温度、水分、土壤理化性质、微生物、植被类型等方面[18-22],虽然获得了不少研究成果,但在不同自然生态系统中研究结果还存在较大差异,主要归结于土壤温度和水分.在冻融条件下频繁的水热交替会影响土壤微生物活性,改变土壤团聚体结构和对土壤无机氮固持[23].有研究发现,在一定模拟条件下,土壤水分和温度与土壤氮矿化之间存在线性相关[24];但也有研究发现土壤氮矿化与温度之间并没有直接关系[25].国内外在不同土地类型(湿地[26]、草地[27]、田地[28]、水稻[29])、不同坡面[30]、不同纬度[31-32]等条件下通过室内模拟和原位培养的方法已经开展了无机氮变化影响因子的研究.其中,室内模拟实验容易改变土壤温度和水分条件,可得到土壤无机氮变化规律,但是由于研究方法和冻融格局的差异,得到的结果也不一致[33].与室内模拟冻融实验相比,野外原位实验弥补了温度、水分极端变化的缺陷,很大程度上还原了无机氮在自然条件下的变化规律,但目前野外原位监测研究仍非常少见,导致秋季冻融对土壤无机氮变化的影响尚不明确.我国关于冻融条件下无机氮的研究主要集中在森林[34]、农田[35]、山地[31]、湿地[36]等生态系统,但在多年冻土泥炭地研究较少.因此,在气候变暖、多年冻土退化背景下,深入探讨秋季冻融对多年冻土泥炭地无机氮影响的研究是必要且紧迫的. ...
丹江口库区覆膜土壤不同土层氮素矿化速率及其影响因素
1
2018
... 目前关于无机氮影响因素的研究主要集中在温度、水分、土壤理化性质、微生物、植被类型等方面[18-22],虽然获得了不少研究成果,但在不同自然生态系统中研究结果还存在较大差异,主要归结于土壤温度和水分.在冻融条件下频繁的水热交替会影响土壤微生物活性,改变土壤团聚体结构和对土壤无机氮固持[23].有研究发现,在一定模拟条件下,土壤水分和温度与土壤氮矿化之间存在线性相关[24];但也有研究发现土壤氮矿化与温度之间并没有直接关系[25].国内外在不同土地类型(湿地[26]、草地[27]、田地[28]、水稻[29])、不同坡面[30]、不同纬度[31-32]等条件下通过室内模拟和原位培养的方法已经开展了无机氮变化影响因子的研究.其中,室内模拟实验容易改变土壤温度和水分条件,可得到土壤无机氮变化规律,但是由于研究方法和冻融格局的差异,得到的结果也不一致[33].与室内模拟冻融实验相比,野外原位实验弥补了温度、水分极端变化的缺陷,很大程度上还原了无机氮在自然条件下的变化规律,但目前野外原位监测研究仍非常少见,导致秋季冻融对土壤无机氮变化的影响尚不明确.我国关于冻融条件下无机氮的研究主要集中在森林[34]、农田[35]、山地[31]、湿地[36]等生态系统,但在多年冻土泥炭地研究较少.因此,在气候变暖、多年冻土退化背景下,深入探讨秋季冻融对多年冻土泥炭地无机氮影响的研究是必要且紧迫的. ...
Sources of C and N contributing to the flush in mineralization upon freeze-thaw cycles in soils
1
2002
... 目前关于无机氮影响因素的研究主要集中在温度、水分、土壤理化性质、微生物、植被类型等方面[18-22],虽然获得了不少研究成果,但在不同自然生态系统中研究结果还存在较大差异,主要归结于土壤温度和水分.在冻融条件下频繁的水热交替会影响土壤微生物活性,改变土壤团聚体结构和对土壤无机氮固持[23].有研究发现,在一定模拟条件下,土壤水分和温度与土壤氮矿化之间存在线性相关[24];但也有研究发现土壤氮矿化与温度之间并没有直接关系[25].国内外在不同土地类型(湿地[26]、草地[27]、田地[28]、水稻[29])、不同坡面[30]、不同纬度[31-32]等条件下通过室内模拟和原位培养的方法已经开展了无机氮变化影响因子的研究.其中,室内模拟实验容易改变土壤温度和水分条件,可得到土壤无机氮变化规律,但是由于研究方法和冻融格局的差异,得到的结果也不一致[33].与室内模拟冻融实验相比,野外原位实验弥补了温度、水分极端变化的缺陷,很大程度上还原了无机氮在自然条件下的变化规律,但目前野外原位监测研究仍非常少见,导致秋季冻融对土壤无机氮变化的影响尚不明确.我国关于冻融条件下无机氮的研究主要集中在森林[34]、农田[35]、山地[31]、湿地[36]等生态系统,但在多年冻土泥炭地研究较少.因此,在气候变暖、多年冻土退化背景下,深入探讨秋季冻融对多年冻土泥炭地无机氮影响的研究是必要且紧迫的. ...
Effects of temperature and moisture on soil nitrogen mineralization of Phyllostachys heterocycla plantation in the rainy area of Western China
1
2018
... 目前关于无机氮影响因素的研究主要集中在温度、水分、土壤理化性质、微生物、植被类型等方面[18-22],虽然获得了不少研究成果,但在不同自然生态系统中研究结果还存在较大差异,主要归结于土壤温度和水分.在冻融条件下频繁的水热交替会影响土壤微生物活性,改变土壤团聚体结构和对土壤无机氮固持[23].有研究发现,在一定模拟条件下,土壤水分和温度与土壤氮矿化之间存在线性相关[24];但也有研究发现土壤氮矿化与温度之间并没有直接关系[25].国内外在不同土地类型(湿地[26]、草地[27]、田地[28]、水稻[29])、不同坡面[30]、不同纬度[31-32]等条件下通过室内模拟和原位培养的方法已经开展了无机氮变化影响因子的研究.其中,室内模拟实验容易改变土壤温度和水分条件,可得到土壤无机氮变化规律,但是由于研究方法和冻融格局的差异,得到的结果也不一致[33].与室内模拟冻融实验相比,野外原位实验弥补了温度、水分极端变化的缺陷,很大程度上还原了无机氮在自然条件下的变化规律,但目前野外原位监测研究仍非常少见,导致秋季冻融对土壤无机氮变化的影响尚不明确.我国关于冻融条件下无机氮的研究主要集中在森林[34]、农田[35]、山地[31]、湿地[36]等生态系统,但在多年冻土泥炭地研究较少.因此,在气候变暖、多年冻土退化背景下,深入探讨秋季冻融对多年冻土泥炭地无机氮影响的研究是必要且紧迫的. ...
温度和水分对华西雨屏区毛竹林土壤氮矿化的影响
1
2018
... 目前关于无机氮影响因素的研究主要集中在温度、水分、土壤理化性质、微生物、植被类型等方面[18-22],虽然获得了不少研究成果,但在不同自然生态系统中研究结果还存在较大差异,主要归结于土壤温度和水分.在冻融条件下频繁的水热交替会影响土壤微生物活性,改变土壤团聚体结构和对土壤无机氮固持[23].有研究发现,在一定模拟条件下,土壤水分和温度与土壤氮矿化之间存在线性相关[24];但也有研究发现土壤氮矿化与温度之间并没有直接关系[25].国内外在不同土地类型(湿地[26]、草地[27]、田地[28]、水稻[29])、不同坡面[30]、不同纬度[31-32]等条件下通过室内模拟和原位培养的方法已经开展了无机氮变化影响因子的研究.其中,室内模拟实验容易改变土壤温度和水分条件,可得到土壤无机氮变化规律,但是由于研究方法和冻融格局的差异,得到的结果也不一致[33].与室内模拟冻融实验相比,野外原位实验弥补了温度、水分极端变化的缺陷,很大程度上还原了无机氮在自然条件下的变化规律,但目前野外原位监测研究仍非常少见,导致秋季冻融对土壤无机氮变化的影响尚不明确.我国关于冻融条件下无机氮的研究主要集中在森林[34]、农田[35]、山地[31]、湿地[36]等生态系统,但在多年冻土泥炭地研究较少.因此,在气候变暖、多年冻土退化背景下,深入探讨秋季冻融对多年冻土泥炭地无机氮影响的研究是必要且紧迫的. ...
Effect of temperature on humus respiration rate and nitrogen mineralization: implications for global climate change
1
1999
... 目前关于无机氮影响因素的研究主要集中在温度、水分、土壤理化性质、微生物、植被类型等方面[18-22],虽然获得了不少研究成果,但在不同自然生态系统中研究结果还存在较大差异,主要归结于土壤温度和水分.在冻融条件下频繁的水热交替会影响土壤微生物活性,改变土壤团聚体结构和对土壤无机氮固持[23].有研究发现,在一定模拟条件下,土壤水分和温度与土壤氮矿化之间存在线性相关[24];但也有研究发现土壤氮矿化与温度之间并没有直接关系[25].国内外在不同土地类型(湿地[26]、草地[27]、田地[28]、水稻[29])、不同坡面[30]、不同纬度[31-32]等条件下通过室内模拟和原位培养的方法已经开展了无机氮变化影响因子的研究.其中,室内模拟实验容易改变土壤温度和水分条件,可得到土壤无机氮变化规律,但是由于研究方法和冻融格局的差异,得到的结果也不一致[33].与室内模拟冻融实验相比,野外原位实验弥补了温度、水分极端变化的缺陷,很大程度上还原了无机氮在自然条件下的变化规律,但目前野外原位监测研究仍非常少见,导致秋季冻融对土壤无机氮变化的影响尚不明确.我国关于冻融条件下无机氮的研究主要集中在森林[34]、农田[35]、山地[31]、湿地[36]等生态系统,但在多年冻土泥炭地研究较少.因此,在气候变暖、多年冻土退化背景下,深入探讨秋季冻融对多年冻土泥炭地无机氮影响的研究是必要且紧迫的. ...
Effect of simulated emerald ash borer infestation on nitrogen cycling in black ash (Fraxinus nigra) wetlands in northern Minnesota, USA
1
2020
... 目前关于无机氮影响因素的研究主要集中在温度、水分、土壤理化性质、微生物、植被类型等方面[18-22],虽然获得了不少研究成果,但在不同自然生态系统中研究结果还存在较大差异,主要归结于土壤温度和水分.在冻融条件下频繁的水热交替会影响土壤微生物活性,改变土壤团聚体结构和对土壤无机氮固持[23].有研究发现,在一定模拟条件下,土壤水分和温度与土壤氮矿化之间存在线性相关[24];但也有研究发现土壤氮矿化与温度之间并没有直接关系[25].国内外在不同土地类型(湿地[26]、草地[27]、田地[28]、水稻[29])、不同坡面[30]、不同纬度[31-32]等条件下通过室内模拟和原位培养的方法已经开展了无机氮变化影响因子的研究.其中,室内模拟实验容易改变土壤温度和水分条件,可得到土壤无机氮变化规律,但是由于研究方法和冻融格局的差异,得到的结果也不一致[33].与室内模拟冻融实验相比,野外原位实验弥补了温度、水分极端变化的缺陷,很大程度上还原了无机氮在自然条件下的变化规律,但目前野外原位监测研究仍非常少见,导致秋季冻融对土壤无机氮变化的影响尚不明确.我国关于冻融条件下无机氮的研究主要集中在森林[34]、农田[35]、山地[31]、湿地[36]等生态系统,但在多年冻土泥炭地研究较少.因此,在气候变暖、多年冻土退化背景下,深入探讨秋季冻融对多年冻土泥炭地无机氮影响的研究是必要且紧迫的. ...
Effects of warming and nitrogen deposition on the coupling mechanism between soil nitrogen and phosphorus in Songnen Meadow Steppe, northeastern China
1
2013
... 目前关于无机氮影响因素的研究主要集中在温度、水分、土壤理化性质、微生物、植被类型等方面[18-22],虽然获得了不少研究成果,但在不同自然生态系统中研究结果还存在较大差异,主要归结于土壤温度和水分.在冻融条件下频繁的水热交替会影响土壤微生物活性,改变土壤团聚体结构和对土壤无机氮固持[23].有研究发现,在一定模拟条件下,土壤水分和温度与土壤氮矿化之间存在线性相关[24];但也有研究发现土壤氮矿化与温度之间并没有直接关系[25].国内外在不同土地类型(湿地[26]、草地[27]、田地[28]、水稻[29])、不同坡面[30]、不同纬度[31-32]等条件下通过室内模拟和原位培养的方法已经开展了无机氮变化影响因子的研究.其中,室内模拟实验容易改变土壤温度和水分条件,可得到土壤无机氮变化规律,但是由于研究方法和冻融格局的差异,得到的结果也不一致[33].与室内模拟冻融实验相比,野外原位实验弥补了温度、水分极端变化的缺陷,很大程度上还原了无机氮在自然条件下的变化规律,但目前野外原位监测研究仍非常少见,导致秋季冻融对土壤无机氮变化的影响尚不明确.我国关于冻融条件下无机氮的研究主要集中在森林[34]、农田[35]、山地[31]、湿地[36]等生态系统,但在多年冻土泥炭地研究较少.因此,在气候变暖、多年冻土退化背景下,深入探讨秋季冻融对多年冻土泥炭地无机氮影响的研究是必要且紧迫的. ...
Field and laboratory study of nitrogen mineralization dynamics in four Tunisian soils
1
2019
... 目前关于无机氮影响因素的研究主要集中在温度、水分、土壤理化性质、微生物、植被类型等方面[18-22],虽然获得了不少研究成果,但在不同自然生态系统中研究结果还存在较大差异,主要归结于土壤温度和水分.在冻融条件下频繁的水热交替会影响土壤微生物活性,改变土壤团聚体结构和对土壤无机氮固持[23].有研究发现,在一定模拟条件下,土壤水分和温度与土壤氮矿化之间存在线性相关[24];但也有研究发现土壤氮矿化与温度之间并没有直接关系[25].国内外在不同土地类型(湿地[26]、草地[27]、田地[28]、水稻[29])、不同坡面[30]、不同纬度[31-32]等条件下通过室内模拟和原位培养的方法已经开展了无机氮变化影响因子的研究.其中,室内模拟实验容易改变土壤温度和水分条件,可得到土壤无机氮变化规律,但是由于研究方法和冻融格局的差异,得到的结果也不一致[33].与室内模拟冻融实验相比,野外原位实验弥补了温度、水分极端变化的缺陷,很大程度上还原了无机氮在自然条件下的变化规律,但目前野外原位监测研究仍非常少见,导致秋季冻融对土壤无机氮变化的影响尚不明确.我国关于冻融条件下无机氮的研究主要集中在森林[34]、农田[35]、山地[31]、湿地[36]等生态系统,但在多年冻土泥炭地研究较少.因此,在气候变暖、多年冻土退化背景下,深入探讨秋季冻融对多年冻土泥炭地无机氮影响的研究是必要且紧迫的. ...
Soil aeration and relationship to inorganic nitrogen during aerobic cultivation of irrigated rice on a consolidated land parcel
1
2020
... 目前关于无机氮影响因素的研究主要集中在温度、水分、土壤理化性质、微生物、植被类型等方面[18-22],虽然获得了不少研究成果,但在不同自然生态系统中研究结果还存在较大差异,主要归结于土壤温度和水分.在冻融条件下频繁的水热交替会影响土壤微生物活性,改变土壤团聚体结构和对土壤无机氮固持[23].有研究发现,在一定模拟条件下,土壤水分和温度与土壤氮矿化之间存在线性相关[24];但也有研究发现土壤氮矿化与温度之间并没有直接关系[25].国内外在不同土地类型(湿地[26]、草地[27]、田地[28]、水稻[29])、不同坡面[30]、不同纬度[31-32]等条件下通过室内模拟和原位培养的方法已经开展了无机氮变化影响因子的研究.其中,室内模拟实验容易改变土壤温度和水分条件,可得到土壤无机氮变化规律,但是由于研究方法和冻融格局的差异,得到的结果也不一致[33].与室内模拟冻融实验相比,野外原位实验弥补了温度、水分极端变化的缺陷,很大程度上还原了无机氮在自然条件下的变化规律,但目前野外原位监测研究仍非常少见,导致秋季冻融对土壤无机氮变化的影响尚不明确.我国关于冻融条件下无机氮的研究主要集中在森林[34]、农田[35]、山地[31]、湿地[36]等生态系统,但在多年冻土泥炭地研究较少.因此,在气候变暖、多年冻土退化背景下,深入探讨秋季冻融对多年冻土泥炭地无机氮影响的研究是必要且紧迫的. ...
Discriminating surface soil inorganic nitrogen cycling under various land uses in a watershed with simulations of energy balanced temperature and slope introduced moisture
1
2020
... 目前关于无机氮影响因素的研究主要集中在温度、水分、土壤理化性质、微生物、植被类型等方面[18-22],虽然获得了不少研究成果,但在不同自然生态系统中研究结果还存在较大差异,主要归结于土壤温度和水分.在冻融条件下频繁的水热交替会影响土壤微生物活性,改变土壤团聚体结构和对土壤无机氮固持[23].有研究发现,在一定模拟条件下,土壤水分和温度与土壤氮矿化之间存在线性相关[24];但也有研究发现土壤氮矿化与温度之间并没有直接关系[25].国内外在不同土地类型(湿地[26]、草地[27]、田地[28]、水稻[29])、不同坡面[30]、不同纬度[31-32]等条件下通过室内模拟和原位培养的方法已经开展了无机氮变化影响因子的研究.其中,室内模拟实验容易改变土壤温度和水分条件,可得到土壤无机氮变化规律,但是由于研究方法和冻融格局的差异,得到的结果也不一致[33].与室内模拟冻融实验相比,野外原位实验弥补了温度、水分极端变化的缺陷,很大程度上还原了无机氮在自然条件下的变化规律,但目前野外原位监测研究仍非常少见,导致秋季冻融对土壤无机氮变化的影响尚不明确.我国关于冻融条件下无机氮的研究主要集中在森林[34]、农田[35]、山地[31]、湿地[36]等生态系统,但在多年冻土泥炭地研究较少.因此,在气候变暖、多年冻土退化背景下,深入探讨秋季冻融对多年冻土泥炭地无机氮影响的研究是必要且紧迫的. ...
Dynamic response of soil nitrogen to freeze-thaw processes in different cenotypes in the forests of the Tianshan Mountains
3
2019
... 目前关于无机氮影响因素的研究主要集中在温度、水分、土壤理化性质、微生物、植被类型等方面[18-22],虽然获得了不少研究成果,但在不同自然生态系统中研究结果还存在较大差异,主要归结于土壤温度和水分.在冻融条件下频繁的水热交替会影响土壤微生物活性,改变土壤团聚体结构和对土壤无机氮固持[23].有研究发现,在一定模拟条件下,土壤水分和温度与土壤氮矿化之间存在线性相关[24];但也有研究发现土壤氮矿化与温度之间并没有直接关系[25].国内外在不同土地类型(湿地[26]、草地[27]、田地[28]、水稻[29])、不同坡面[30]、不同纬度[31-32]等条件下通过室内模拟和原位培养的方法已经开展了无机氮变化影响因子的研究.其中,室内模拟实验容易改变土壤温度和水分条件,可得到土壤无机氮变化规律,但是由于研究方法和冻融格局的差异,得到的结果也不一致[33].与室内模拟冻融实验相比,野外原位实验弥补了温度、水分极端变化的缺陷,很大程度上还原了无机氮在自然条件下的变化规律,但目前野外原位监测研究仍非常少见,导致秋季冻融对土壤无机氮变化的影响尚不明确.我国关于冻融条件下无机氮的研究主要集中在森林[34]、农田[35]、山地[31]、湿地[36]等生态系统,但在多年冻土泥炭地研究较少.因此,在气候变暖、多年冻土退化背景下,深入探讨秋季冻融对多年冻土泥炭地无机氮影响的研究是必要且紧迫的. ...
... [31]、湿地[36]等生态系统,但在多年冻土泥炭地研究较少.因此,在气候变暖、多年冻土退化背景下,深入探讨秋季冻融对多年冻土泥炭地无机氮影响的研究是必要且紧迫的. ...
... Comparisons of inorganic nitrogen changes under controlled hydrothermal conditions
Table 5| 生态系统 | 研究地点 | 经纬度 | 实验类型 | 冻融处理/次数 | 含水量 | NH4+-N | NO3--N |
|---|
| 森林 | 长白山地区温带森林土壤 | 127° E, 42° N[34] | LT | -10 ℃→10 ℃, 12 h/1、3、5、7、11、15、22、28、36次 | 秋季低含水量(40%) | 不变 | 增加 |
| 127° E, 42° N[57] | LT | -20 ℃→ 15 ℃, 12 h/1、3、5、9、14、21、28、35次 | 最大田间持水量的60% | 增加 | 增加 |
| 河岸带不同植被类型 | 123° E, 41° N[58] | LT | -5 ℃、-20 ℃→ 5℃, 12 h/0、1、5、10、20、30次 | 原土壤含水量 | 增加 | 农田变化不显著 |
| 天山森林土壤 | 87° E, 43° N[31] | ST | 0 ℃以上→ 0 ℃以下, 3 h/3次 | 原土壤含水量 | 草地和灌丛表现为先减少后增加 | 增加 |
| 大兴安岭落叶松林土壤 | 121° E, 50° N[38] | LT | -5 ℃→ 8 ℃, 24 h/1、3、5、7、15次 | 60%土壤饱和含水量 | 低于恒温条件下的累积量 | 低于恒温条件下的累积量 |
| 草地 | 青藏高原高寒草甸区 | 103° E, 32° N[59] | LT | -4 ℃、-20 ℃ → 4℃, 8 h/1、3、5、25、60次 | 原土壤含水量 | 先增后减 | 先增后减 |
| 湿地 | 三江平原沼泽湿地 | 133° E, 47° N[36] | LT | -5 ℃、-25 ℃→ 5 ℃, 1 d/1、2、4、6、10次 | (352.5±1.6) g·kg-1 | 增加 | 增加 |
| 小兴安岭湿地土壤 | 129° E, 48° N[60] | LT | -20 ℃→ 5 ℃, 24 h/5次 | 原土壤含水量 | 先增后减 | 先减后增 |
| 128° E, 48° N[61] | LT | -5 ℃、-25 ℃→ 5 ℃, 24 h/0、1、2、4、9次 | 原土壤含水量 | 先增后减 | 先减后增 |
注:LT:室内模拟试验;ST:野外原位试验. ...
天山林区不同类型群落土壤氮素对冻融过程的动态响应
3
2019
... 目前关于无机氮影响因素的研究主要集中在温度、水分、土壤理化性质、微生物、植被类型等方面[18-22],虽然获得了不少研究成果,但在不同自然生态系统中研究结果还存在较大差异,主要归结于土壤温度和水分.在冻融条件下频繁的水热交替会影响土壤微生物活性,改变土壤团聚体结构和对土壤无机氮固持[23].有研究发现,在一定模拟条件下,土壤水分和温度与土壤氮矿化之间存在线性相关[24];但也有研究发现土壤氮矿化与温度之间并没有直接关系[25].国内外在不同土地类型(湿地[26]、草地[27]、田地[28]、水稻[29])、不同坡面[30]、不同纬度[31-32]等条件下通过室内模拟和原位培养的方法已经开展了无机氮变化影响因子的研究.其中,室内模拟实验容易改变土壤温度和水分条件,可得到土壤无机氮变化规律,但是由于研究方法和冻融格局的差异,得到的结果也不一致[33].与室内模拟冻融实验相比,野外原位实验弥补了温度、水分极端变化的缺陷,很大程度上还原了无机氮在自然条件下的变化规律,但目前野外原位监测研究仍非常少见,导致秋季冻融对土壤无机氮变化的影响尚不明确.我国关于冻融条件下无机氮的研究主要集中在森林[34]、农田[35]、山地[31]、湿地[36]等生态系统,但在多年冻土泥炭地研究较少.因此,在气候变暖、多年冻土退化背景下,深入探讨秋季冻融对多年冻土泥炭地无机氮影响的研究是必要且紧迫的. ...
... [31]、湿地[36]等生态系统,但在多年冻土泥炭地研究较少.因此,在气候变暖、多年冻土退化背景下,深入探讨秋季冻融对多年冻土泥炭地无机氮影响的研究是必要且紧迫的. ...
... Comparisons of inorganic nitrogen changes under controlled hydrothermal conditions
Table 5| 生态系统 | 研究地点 | 经纬度 | 实验类型 | 冻融处理/次数 | 含水量 | NH4+-N | NO3--N |
|---|
| 森林 | 长白山地区温带森林土壤 | 127° E, 42° N[34] | LT | -10 ℃→10 ℃, 12 h/1、3、5、7、11、15、22、28、36次 | 秋季低含水量(40%) | 不变 | 增加 |
| 127° E, 42° N[57] | LT | -20 ℃→ 15 ℃, 12 h/1、3、5、9、14、21、28、35次 | 最大田间持水量的60% | 增加 | 增加 |
| 河岸带不同植被类型 | 123° E, 41° N[58] | LT | -5 ℃、-20 ℃→ 5℃, 12 h/0、1、5、10、20、30次 | 原土壤含水量 | 增加 | 农田变化不显著 |
| 天山森林土壤 | 87° E, 43° N[31] | ST | 0 ℃以上→ 0 ℃以下, 3 h/3次 | 原土壤含水量 | 草地和灌丛表现为先减少后增加 | 增加 |
| 大兴安岭落叶松林土壤 | 121° E, 50° N[38] | LT | -5 ℃→ 8 ℃, 24 h/1、3、5、7、15次 | 60%土壤饱和含水量 | 低于恒温条件下的累积量 | 低于恒温条件下的累积量 |
| 草地 | 青藏高原高寒草甸区 | 103° E, 32° N[59] | LT | -4 ℃、-20 ℃ → 4℃, 8 h/1、3、5、25、60次 | 原土壤含水量 | 先增后减 | 先增后减 |
| 湿地 | 三江平原沼泽湿地 | 133° E, 47° N[36] | LT | -5 ℃、-25 ℃→ 5 ℃, 1 d/1、2、4、6、10次 | (352.5±1.6) g·kg-1 | 增加 | 增加 |
| 小兴安岭湿地土壤 | 129° E, 48° N[60] | LT | -20 ℃→ 5 ℃, 24 h/5次 | 原土壤含水量 | 先增后减 | 先减后增 |
| 128° E, 48° N[61] | LT | -5 ℃、-25 ℃→ 5 ℃, 24 h/0、1、2、4、9次 | 原土壤含水量 | 先增后减 | 先减后增 |
注:LT:室内模拟试验;ST:野外原位试验. ...
Effects of moisture and temperature on soil nitrogen mineralization of Cryptomeria fortunei plantations in rainy area of Western China
1
2014
... 目前关于无机氮影响因素的研究主要集中在温度、水分、土壤理化性质、微生物、植被类型等方面[18-22],虽然获得了不少研究成果,但在不同自然生态系统中研究结果还存在较大差异,主要归结于土壤温度和水分.在冻融条件下频繁的水热交替会影响土壤微生物活性,改变土壤团聚体结构和对土壤无机氮固持[23].有研究发现,在一定模拟条件下,土壤水分和温度与土壤氮矿化之间存在线性相关[24];但也有研究发现土壤氮矿化与温度之间并没有直接关系[25].国内外在不同土地类型(湿地[26]、草地[27]、田地[28]、水稻[29])、不同坡面[30]、不同纬度[31-32]等条件下通过室内模拟和原位培养的方法已经开展了无机氮变化影响因子的研究.其中,室内模拟实验容易改变土壤温度和水分条件,可得到土壤无机氮变化规律,但是由于研究方法和冻融格局的差异,得到的结果也不一致[33].与室内模拟冻融实验相比,野外原位实验弥补了温度、水分极端变化的缺陷,很大程度上还原了无机氮在自然条件下的变化规律,但目前野外原位监测研究仍非常少见,导致秋季冻融对土壤无机氮变化的影响尚不明确.我国关于冻融条件下无机氮的研究主要集中在森林[34]、农田[35]、山地[31]、湿地[36]等生态系统,但在多年冻土泥炭地研究较少.因此,在气候变暖、多年冻土退化背景下,深入探讨秋季冻融对多年冻土泥炭地无机氮影响的研究是必要且紧迫的. ...
水热条件对华西雨屏区柳杉人工林土壤氮矿化的影响
1
2014
... 目前关于无机氮影响因素的研究主要集中在温度、水分、土壤理化性质、微生物、植被类型等方面[18-22],虽然获得了不少研究成果,但在不同自然生态系统中研究结果还存在较大差异,主要归结于土壤温度和水分.在冻融条件下频繁的水热交替会影响土壤微生物活性,改变土壤团聚体结构和对土壤无机氮固持[23].有研究发现,在一定模拟条件下,土壤水分和温度与土壤氮矿化之间存在线性相关[24];但也有研究发现土壤氮矿化与温度之间并没有直接关系[25].国内外在不同土地类型(湿地[26]、草地[27]、田地[28]、水稻[29])、不同坡面[30]、不同纬度[31-32]等条件下通过室内模拟和原位培养的方法已经开展了无机氮变化影响因子的研究.其中,室内模拟实验容易改变土壤温度和水分条件,可得到土壤无机氮变化规律,但是由于研究方法和冻融格局的差异,得到的结果也不一致[33].与室内模拟冻融实验相比,野外原位实验弥补了温度、水分极端变化的缺陷,很大程度上还原了无机氮在自然条件下的变化规律,但目前野外原位监测研究仍非常少见,导致秋季冻融对土壤无机氮变化的影响尚不明确.我国关于冻融条件下无机氮的研究主要集中在森林[34]、农田[35]、山地[31]、湿地[36]等生态系统,但在多年冻土泥炭地研究较少.因此,在气候变暖、多年冻土退化背景下,深入探讨秋季冻融对多年冻土泥炭地无机氮影响的研究是必要且紧迫的. ...
Freeze-thaw cycles and their impacts on ecological process: a review
1
2008
... 目前关于无机氮影响因素的研究主要集中在温度、水分、土壤理化性质、微生物、植被类型等方面[18-22],虽然获得了不少研究成果,但在不同自然生态系统中研究结果还存在较大差异,主要归结于土壤温度和水分.在冻融条件下频繁的水热交替会影响土壤微生物活性,改变土壤团聚体结构和对土壤无机氮固持[23].有研究发现,在一定模拟条件下,土壤水分和温度与土壤氮矿化之间存在线性相关[24];但也有研究发现土壤氮矿化与温度之间并没有直接关系[25].国内外在不同土地类型(湿地[26]、草地[27]、田地[28]、水稻[29])、不同坡面[30]、不同纬度[31-32]等条件下通过室内模拟和原位培养的方法已经开展了无机氮变化影响因子的研究.其中,室内模拟实验容易改变土壤温度和水分条件,可得到土壤无机氮变化规律,但是由于研究方法和冻融格局的差异,得到的结果也不一致[33].与室内模拟冻融实验相比,野外原位实验弥补了温度、水分极端变化的缺陷,很大程度上还原了无机氮在自然条件下的变化规律,但目前野外原位监测研究仍非常少见,导致秋季冻融对土壤无机氮变化的影响尚不明确.我国关于冻融条件下无机氮的研究主要集中在森林[34]、农田[35]、山地[31]、湿地[36]等生态系统,但在多年冻土泥炭地研究较少.因此,在气候变暖、多年冻土退化背景下,深入探讨秋季冻融对多年冻土泥炭地无机氮影响的研究是必要且紧迫的. ...
土壤冻融交替生态效应研究进展
1
2008
... 目前关于无机氮影响因素的研究主要集中在温度、水分、土壤理化性质、微生物、植被类型等方面[18-22],虽然获得了不少研究成果,但在不同自然生态系统中研究结果还存在较大差异,主要归结于土壤温度和水分.在冻融条件下频繁的水热交替会影响土壤微生物活性,改变土壤团聚体结构和对土壤无机氮固持[23].有研究发现,在一定模拟条件下,土壤水分和温度与土壤氮矿化之间存在线性相关[24];但也有研究发现土壤氮矿化与温度之间并没有直接关系[25].国内外在不同土地类型(湿地[26]、草地[27]、田地[28]、水稻[29])、不同坡面[30]、不同纬度[31-32]等条件下通过室内模拟和原位培养的方法已经开展了无机氮变化影响因子的研究.其中,室内模拟实验容易改变土壤温度和水分条件,可得到土壤无机氮变化规律,但是由于研究方法和冻融格局的差异,得到的结果也不一致[33].与室内模拟冻融实验相比,野外原位实验弥补了温度、水分极端变化的缺陷,很大程度上还原了无机氮在自然条件下的变化规律,但目前野外原位监测研究仍非常少见,导致秋季冻融对土壤无机氮变化的影响尚不明确.我国关于冻融条件下无机氮的研究主要集中在森林[34]、农田[35]、山地[31]、湿地[36]等生态系统,但在多年冻土泥炭地研究较少.因此,在气候变暖、多年冻土退化背景下,深入探讨秋季冻融对多年冻土泥炭地无机氮影响的研究是必要且紧迫的. ...
Effects of freezing-thawing on soil carbon and nitrogen mineralization in temperate forest ecosystems
3
2017
... 目前关于无机氮影响因素的研究主要集中在温度、水分、土壤理化性质、微生物、植被类型等方面[18-22],虽然获得了不少研究成果,但在不同自然生态系统中研究结果还存在较大差异,主要归结于土壤温度和水分.在冻融条件下频繁的水热交替会影响土壤微生物活性,改变土壤团聚体结构和对土壤无机氮固持[23].有研究发现,在一定模拟条件下,土壤水分和温度与土壤氮矿化之间存在线性相关[24];但也有研究发现土壤氮矿化与温度之间并没有直接关系[25].国内外在不同土地类型(湿地[26]、草地[27]、田地[28]、水稻[29])、不同坡面[30]、不同纬度[31-32]等条件下通过室内模拟和原位培养的方法已经开展了无机氮变化影响因子的研究.其中,室内模拟实验容易改变土壤温度和水分条件,可得到土壤无机氮变化规律,但是由于研究方法和冻融格局的差异,得到的结果也不一致[33].与室内模拟冻融实验相比,野外原位实验弥补了温度、水分极端变化的缺陷,很大程度上还原了无机氮在自然条件下的变化规律,但目前野外原位监测研究仍非常少见,导致秋季冻融对土壤无机氮变化的影响尚不明确.我国关于冻融条件下无机氮的研究主要集中在森林[34]、农田[35]、山地[31]、湿地[36]等生态系统,但在多年冻土泥炭地研究较少.因此,在气候变暖、多年冻土退化背景下,深入探讨秋季冻融对多年冻土泥炭地无机氮影响的研究是必要且紧迫的. ...
... Comparisons of inorganic nitrogen changes under controlled hydrothermal conditions
Table 5| 生态系统 | 研究地点 | 经纬度 | 实验类型 | 冻融处理/次数 | 含水量 | NH4+-N | NO3--N |
|---|
| 森林 | 长白山地区温带森林土壤 | 127° E, 42° N[34] | LT | -10 ℃→10 ℃, 12 h/1、3、5、7、11、15、22、28、36次 | 秋季低含水量(40%) | 不变 | 增加 |
| 127° E, 42° N[57] | LT | -20 ℃→ 15 ℃, 12 h/1、3、5、9、14、21、28、35次 | 最大田间持水量的60% | 增加 | 增加 |
| 河岸带不同植被类型 | 123° E, 41° N[58] | LT | -5 ℃、-20 ℃→ 5℃, 12 h/0、1、5、10、20、30次 | 原土壤含水量 | 增加 | 农田变化不显著 |
| 天山森林土壤 | 87° E, 43° N[31] | ST | 0 ℃以上→ 0 ℃以下, 3 h/3次 | 原土壤含水量 | 草地和灌丛表现为先减少后增加 | 增加 |
| 大兴安岭落叶松林土壤 | 121° E, 50° N[38] | LT | -5 ℃→ 8 ℃, 24 h/1、3、5、7、15次 | 60%土壤饱和含水量 | 低于恒温条件下的累积量 | 低于恒温条件下的累积量 |
| 草地 | 青藏高原高寒草甸区 | 103° E, 32° N[59] | LT | -4 ℃、-20 ℃ → 4℃, 8 h/1、3、5、25、60次 | 原土壤含水量 | 先增后减 | 先增后减 |
| 湿地 | 三江平原沼泽湿地 | 133° E, 47° N[36] | LT | -5 ℃、-25 ℃→ 5 ℃, 1 d/1、2、4、6、10次 | (352.5±1.6) g·kg-1 | 增加 | 增加 |
| 小兴安岭湿地土壤 | 129° E, 48° N[60] | LT | -20 ℃→ 5 ℃, 24 h/5次 | 原土壤含水量 | 先增后减 | 先减后增 |
| 128° E, 48° N[61] | LT | -5 ℃、-25 ℃→ 5 ℃, 24 h/0、1、2、4、9次 | 原土壤含水量 | 先增后减 | 先减后增 |
注:LT:室内模拟试验;ST:野外原位试验. ...
... 经逐步回归分析表明(表2~4),在不同冻融阶段,无机氮对环境因子的响应程度也存在差异.在整个秋季冻融期,BM浅层(10~20 cm)含水量对无机氮含量的影响程度最大(R2=0.301),这是由于浅层土壤含水量更容易影响土壤通气性和土壤微生物活性,进而改变影响了土壤基质的养分循环,从而对无机氮转化产生影响.通常认为无机氮对温度的响应程度大于含水量[62],但是BM样地在整个秋季冻融时期,温度和含水量都处于较低水平且变化幅度较小,与温度相比含水量对土壤微生物活性起有效作用,这与赵媛[34]在研究秋末冻融循环对长白山地区无机氮的影响结果相似,即无机氮对含水量的响应程度大于温度.在秋季冻融前、中和后期,3种多年冻土泥炭地浅层(0~20 cm)土壤的环境因子与无机氮释放有关,其中浅层土壤温度对无机氮释放的贡献率最大(R2=0.685),这主要受当地非生长季寒冷气候的影响,温度通过改变土壤中生物酶和氨氧化细菌、硝化细菌的活性来影响氮的分解和无机氮的释放[63].但是不同土壤环境条件下微生物对温度敏感程度有很大差异[64],本研究发现,受非生长季温度逐渐降低的影响,XA仅在秋季冻融前期土壤温度对无机氮释放有贡献(R2=0.685),在秋季冻融后期不存在相关关系,这与表5中草地生态系统下青藏高原高寒草甸区研究发现短期的冻融交替可以显著增加土壤中的氮矿化累积结果相似.说明与长期冻融相比,短期温度变化会破坏土壤团聚结构从而影响氨氧化细菌的活性,一部分微生物为了维持繁殖不断汲取死亡微生物的养分,从而促进了微生物的活性,影响土壤氮矿化速度.但也有研究发现,短期冻融对土壤氮矿化并没有影响[57],土壤微生物会对温度变化做出调节反应,从而无机氮变化不显著.秋季冻融期无机氮对环境因子的响应程度存在以上差异的原因可能有以下四方面:第一,样地沿一定坡度选取,地势高低不同造成了土壤含水量存在显著差异.第二,受到地表植被的影响,土壤微生物的类别和数量存在差异因此对氮矿化程度不同[44].第三,与凋落物有关,不同凋落物分解的有机质不同会影响土壤有效基质[65].第四,3种泥炭地土壤的容重、孔隙度和pH值等物理性质存在差异[66].这些要素的综合影响造成了不同泥炭地间,以及冻融期不同时期间无机氮含量对温度和含水量响应程度的不同. ...
冻融对温带森林土壤碳、氮矿化作用的影响
3
2017
... 目前关于无机氮影响因素的研究主要集中在温度、水分、土壤理化性质、微生物、植被类型等方面[18-22],虽然获得了不少研究成果,但在不同自然生态系统中研究结果还存在较大差异,主要归结于土壤温度和水分.在冻融条件下频繁的水热交替会影响土壤微生物活性,改变土壤团聚体结构和对土壤无机氮固持[23].有研究发现,在一定模拟条件下,土壤水分和温度与土壤氮矿化之间存在线性相关[24];但也有研究发现土壤氮矿化与温度之间并没有直接关系[25].国内外在不同土地类型(湿地[26]、草地[27]、田地[28]、水稻[29])、不同坡面[30]、不同纬度[31-32]等条件下通过室内模拟和原位培养的方法已经开展了无机氮变化影响因子的研究.其中,室内模拟实验容易改变土壤温度和水分条件,可得到土壤无机氮变化规律,但是由于研究方法和冻融格局的差异,得到的结果也不一致[33].与室内模拟冻融实验相比,野外原位实验弥补了温度、水分极端变化的缺陷,很大程度上还原了无机氮在自然条件下的变化规律,但目前野外原位监测研究仍非常少见,导致秋季冻融对土壤无机氮变化的影响尚不明确.我国关于冻融条件下无机氮的研究主要集中在森林[34]、农田[35]、山地[31]、湿地[36]等生态系统,但在多年冻土泥炭地研究较少.因此,在气候变暖、多年冻土退化背景下,深入探讨秋季冻融对多年冻土泥炭地无机氮影响的研究是必要且紧迫的. ...
... Comparisons of inorganic nitrogen changes under controlled hydrothermal conditions
Table 5| 生态系统 | 研究地点 | 经纬度 | 实验类型 | 冻融处理/次数 | 含水量 | NH4+-N | NO3--N |
|---|
| 森林 | 长白山地区温带森林土壤 | 127° E, 42° N[34] | LT | -10 ℃→10 ℃, 12 h/1、3、5、7、11、15、22、28、36次 | 秋季低含水量(40%) | 不变 | 增加 |
| 127° E, 42° N[57] | LT | -20 ℃→ 15 ℃, 12 h/1、3、5、9、14、21、28、35次 | 最大田间持水量的60% | 增加 | 增加 |
| 河岸带不同植被类型 | 123° E, 41° N[58] | LT | -5 ℃、-20 ℃→ 5℃, 12 h/0、1、5、10、20、30次 | 原土壤含水量 | 增加 | 农田变化不显著 |
| 天山森林土壤 | 87° E, 43° N[31] | ST | 0 ℃以上→ 0 ℃以下, 3 h/3次 | 原土壤含水量 | 草地和灌丛表现为先减少后增加 | 增加 |
| 大兴安岭落叶松林土壤 | 121° E, 50° N[38] | LT | -5 ℃→ 8 ℃, 24 h/1、3、5、7、15次 | 60%土壤饱和含水量 | 低于恒温条件下的累积量 | 低于恒温条件下的累积量 |
| 草地 | 青藏高原高寒草甸区 | 103° E, 32° N[59] | LT | -4 ℃、-20 ℃ → 4℃, 8 h/1、3、5、25、60次 | 原土壤含水量 | 先增后减 | 先增后减 |
| 湿地 | 三江平原沼泽湿地 | 133° E, 47° N[36] | LT | -5 ℃、-25 ℃→ 5 ℃, 1 d/1、2、4、6、10次 | (352.5±1.6) g·kg-1 | 增加 | 增加 |
| 小兴安岭湿地土壤 | 129° E, 48° N[60] | LT | -20 ℃→ 5 ℃, 24 h/5次 | 原土壤含水量 | 先增后减 | 先减后增 |
| 128° E, 48° N[61] | LT | -5 ℃、-25 ℃→ 5 ℃, 24 h/0、1、2、4、9次 | 原土壤含水量 | 先增后减 | 先减后增 |
注:LT:室内模拟试验;ST:野外原位试验. ...
... 经逐步回归分析表明(表2~4),在不同冻融阶段,无机氮对环境因子的响应程度也存在差异.在整个秋季冻融期,BM浅层(10~20 cm)含水量对无机氮含量的影响程度最大(R2=0.301),这是由于浅层土壤含水量更容易影响土壤通气性和土壤微生物活性,进而改变影响了土壤基质的养分循环,从而对无机氮转化产生影响.通常认为无机氮对温度的响应程度大于含水量[62],但是BM样地在整个秋季冻融时期,温度和含水量都处于较低水平且变化幅度较小,与温度相比含水量对土壤微生物活性起有效作用,这与赵媛[34]在研究秋末冻融循环对长白山地区无机氮的影响结果相似,即无机氮对含水量的响应程度大于温度.在秋季冻融前、中和后期,3种多年冻土泥炭地浅层(0~20 cm)土壤的环境因子与无机氮释放有关,其中浅层土壤温度对无机氮释放的贡献率最大(R2=0.685),这主要受当地非生长季寒冷气候的影响,温度通过改变土壤中生物酶和氨氧化细菌、硝化细菌的活性来影响氮的分解和无机氮的释放[63].但是不同土壤环境条件下微生物对温度敏感程度有很大差异[64],本研究发现,受非生长季温度逐渐降低的影响,XA仅在秋季冻融前期土壤温度对无机氮释放有贡献(R2=0.685),在秋季冻融后期不存在相关关系,这与表5中草地生态系统下青藏高原高寒草甸区研究发现短期的冻融交替可以显著增加土壤中的氮矿化累积结果相似.说明与长期冻融相比,短期温度变化会破坏土壤团聚结构从而影响氨氧化细菌的活性,一部分微生物为了维持繁殖不断汲取死亡微生物的养分,从而促进了微生物的活性,影响土壤氮矿化速度.但也有研究发现,短期冻融对土壤氮矿化并没有影响[57],土壤微生物会对温度变化做出调节反应,从而无机氮变化不显著.秋季冻融期无机氮对环境因子的响应程度存在以上差异的原因可能有以下四方面:第一,样地沿一定坡度选取,地势高低不同造成了土壤含水量存在显著差异.第二,受到地表植被的影响,土壤微生物的类别和数量存在差异因此对氮矿化程度不同[44].第三,与凋落物有关,不同凋落物分解的有机质不同会影响土壤有效基质[65].第四,3种泥炭地土壤的容重、孔隙度和pH值等物理性质存在差异[66].这些要素的综合影响造成了不同泥炭地间,以及冻融期不同时期间无机氮含量对温度和含水量响应程度的不同. ...
Regulation effects of freezing-thawing cycle on farmland brown soil nitrogen transformation process
1
2015
... 目前关于无机氮影响因素的研究主要集中在温度、水分、土壤理化性质、微生物、植被类型等方面[18-22],虽然获得了不少研究成果,但在不同自然生态系统中研究结果还存在较大差异,主要归结于土壤温度和水分.在冻融条件下频繁的水热交替会影响土壤微生物活性,改变土壤团聚体结构和对土壤无机氮固持[23].有研究发现,在一定模拟条件下,土壤水分和温度与土壤氮矿化之间存在线性相关[24];但也有研究发现土壤氮矿化与温度之间并没有直接关系[25].国内外在不同土地类型(湿地[26]、草地[27]、田地[28]、水稻[29])、不同坡面[30]、不同纬度[31-32]等条件下通过室内模拟和原位培养的方法已经开展了无机氮变化影响因子的研究.其中,室内模拟实验容易改变土壤温度和水分条件,可得到土壤无机氮变化规律,但是由于研究方法和冻融格局的差异,得到的结果也不一致[33].与室内模拟冻融实验相比,野外原位实验弥补了温度、水分极端变化的缺陷,很大程度上还原了无机氮在自然条件下的变化规律,但目前野外原位监测研究仍非常少见,导致秋季冻融对土壤无机氮变化的影响尚不明确.我国关于冻融条件下无机氮的研究主要集中在森林[34]、农田[35]、山地[31]、湿地[36]等生态系统,但在多年冻土泥炭地研究较少.因此,在气候变暖、多年冻土退化背景下,深入探讨秋季冻融对多年冻土泥炭地无机氮影响的研究是必要且紧迫的. ...
冻融交替对农田棕壤氮素转化过程的调控效应
1
2015
... 目前关于无机氮影响因素的研究主要集中在温度、水分、土壤理化性质、微生物、植被类型等方面[18-22],虽然获得了不少研究成果,但在不同自然生态系统中研究结果还存在较大差异,主要归结于土壤温度和水分.在冻融条件下频繁的水热交替会影响土壤微生物活性,改变土壤团聚体结构和对土壤无机氮固持[23].有研究发现,在一定模拟条件下,土壤水分和温度与土壤氮矿化之间存在线性相关[24];但也有研究发现土壤氮矿化与温度之间并没有直接关系[25].国内外在不同土地类型(湿地[26]、草地[27]、田地[28]、水稻[29])、不同坡面[30]、不同纬度[31-32]等条件下通过室内模拟和原位培养的方法已经开展了无机氮变化影响因子的研究.其中,室内模拟实验容易改变土壤温度和水分条件,可得到土壤无机氮变化规律,但是由于研究方法和冻融格局的差异,得到的结果也不一致[33].与室内模拟冻融实验相比,野外原位实验弥补了温度、水分极端变化的缺陷,很大程度上还原了无机氮在自然条件下的变化规律,但目前野外原位监测研究仍非常少见,导致秋季冻融对土壤无机氮变化的影响尚不明确.我国关于冻融条件下无机氮的研究主要集中在森林[34]、农田[35]、山地[31]、湿地[36]等生态系统,但在多年冻土泥炭地研究较少.因此,在气候变暖、多年冻土退化背景下,深入探讨秋季冻融对多年冻土泥炭地无机氮影响的研究是必要且紧迫的. ...
Effects of freezing and thawing on dissolved organic carbon and nitrogen pool and nitrogen mineralization in typical wetland soils from Sanjiang Plain, Heilongjiang, China
2
2008
... 目前关于无机氮影响因素的研究主要集中在温度、水分、土壤理化性质、微生物、植被类型等方面[18-22],虽然获得了不少研究成果,但在不同自然生态系统中研究结果还存在较大差异,主要归结于土壤温度和水分.在冻融条件下频繁的水热交替会影响土壤微生物活性,改变土壤团聚体结构和对土壤无机氮固持[23].有研究发现,在一定模拟条件下,土壤水分和温度与土壤氮矿化之间存在线性相关[24];但也有研究发现土壤氮矿化与温度之间并没有直接关系[25].国内外在不同土地类型(湿地[26]、草地[27]、田地[28]、水稻[29])、不同坡面[30]、不同纬度[31-32]等条件下通过室内模拟和原位培养的方法已经开展了无机氮变化影响因子的研究.其中,室内模拟实验容易改变土壤温度和水分条件,可得到土壤无机氮变化规律,但是由于研究方法和冻融格局的差异,得到的结果也不一致[33].与室内模拟冻融实验相比,野外原位实验弥补了温度、水分极端变化的缺陷,很大程度上还原了无机氮在自然条件下的变化规律,但目前野外原位监测研究仍非常少见,导致秋季冻融对土壤无机氮变化的影响尚不明确.我国关于冻融条件下无机氮的研究主要集中在森林[34]、农田[35]、山地[31]、湿地[36]等生态系统,但在多年冻土泥炭地研究较少.因此,在气候变暖、多年冻土退化背景下,深入探讨秋季冻融对多年冻土泥炭地无机氮影响的研究是必要且紧迫的. ...
... Comparisons of inorganic nitrogen changes under controlled hydrothermal conditions
Table 5| 生态系统 | 研究地点 | 经纬度 | 实验类型 | 冻融处理/次数 | 含水量 | NH4+-N | NO3--N |
|---|
| 森林 | 长白山地区温带森林土壤 | 127° E, 42° N[34] | LT | -10 ℃→10 ℃, 12 h/1、3、5、7、11、15、22、28、36次 | 秋季低含水量(40%) | 不变 | 增加 |
| 127° E, 42° N[57] | LT | -20 ℃→ 15 ℃, 12 h/1、3、5、9、14、21、28、35次 | 最大田间持水量的60% | 增加 | 增加 |
| 河岸带不同植被类型 | 123° E, 41° N[58] | LT | -5 ℃、-20 ℃→ 5℃, 12 h/0、1、5、10、20、30次 | 原土壤含水量 | 增加 | 农田变化不显著 |
| 天山森林土壤 | 87° E, 43° N[31] | ST | 0 ℃以上→ 0 ℃以下, 3 h/3次 | 原土壤含水量 | 草地和灌丛表现为先减少后增加 | 增加 |
| 大兴安岭落叶松林土壤 | 121° E, 50° N[38] | LT | -5 ℃→ 8 ℃, 24 h/1、3、5、7、15次 | 60%土壤饱和含水量 | 低于恒温条件下的累积量 | 低于恒温条件下的累积量 |
| 草地 | 青藏高原高寒草甸区 | 103° E, 32° N[59] | LT | -4 ℃、-20 ℃ → 4℃, 8 h/1、3、5、25、60次 | 原土壤含水量 | 先增后减 | 先增后减 |
| 湿地 | 三江平原沼泽湿地 | 133° E, 47° N[36] | LT | -5 ℃、-25 ℃→ 5 ℃, 1 d/1、2、4、6、10次 | (352.5±1.6) g·kg-1 | 增加 | 增加 |
| 小兴安岭湿地土壤 | 129° E, 48° N[60] | LT | -20 ℃→ 5 ℃, 24 h/5次 | 原土壤含水量 | 先增后减 | 先减后增 |
| 128° E, 48° N[61] | LT | -5 ℃、-25 ℃→ 5 ℃, 24 h/0、1、2、4、9次 | 原土壤含水量 | 先增后减 | 先减后增 |
注:LT:室内模拟试验;ST:野外原位试验. ...
冻融对湿地土壤可溶性碳、氮和氮矿化的影响
2
2008
... 目前关于无机氮影响因素的研究主要集中在温度、水分、土壤理化性质、微生物、植被类型等方面[18-22],虽然获得了不少研究成果,但在不同自然生态系统中研究结果还存在较大差异,主要归结于土壤温度和水分.在冻融条件下频繁的水热交替会影响土壤微生物活性,改变土壤团聚体结构和对土壤无机氮固持[23].有研究发现,在一定模拟条件下,土壤水分和温度与土壤氮矿化之间存在线性相关[24];但也有研究发现土壤氮矿化与温度之间并没有直接关系[25].国内外在不同土地类型(湿地[26]、草地[27]、田地[28]、水稻[29])、不同坡面[30]、不同纬度[31-32]等条件下通过室内模拟和原位培养的方法已经开展了无机氮变化影响因子的研究.其中,室内模拟实验容易改变土壤温度和水分条件,可得到土壤无机氮变化规律,但是由于研究方法和冻融格局的差异,得到的结果也不一致[33].与室内模拟冻融实验相比,野外原位实验弥补了温度、水分极端变化的缺陷,很大程度上还原了无机氮在自然条件下的变化规律,但目前野外原位监测研究仍非常少见,导致秋季冻融对土壤无机氮变化的影响尚不明确.我国关于冻融条件下无机氮的研究主要集中在森林[34]、农田[35]、山地[31]、湿地[36]等生态系统,但在多年冻土泥炭地研究较少.因此,在气候变暖、多年冻土退化背景下,深入探讨秋季冻融对多年冻土泥炭地无机氮影响的研究是必要且紧迫的. ...
... Comparisons of inorganic nitrogen changes under controlled hydrothermal conditions
Table 5| 生态系统 | 研究地点 | 经纬度 | 实验类型 | 冻融处理/次数 | 含水量 | NH4+-N | NO3--N |
|---|
| 森林 | 长白山地区温带森林土壤 | 127° E, 42° N[34] | LT | -10 ℃→10 ℃, 12 h/1、3、5、7、11、15、22、28、36次 | 秋季低含水量(40%) | 不变 | 增加 |
| 127° E, 42° N[57] | LT | -20 ℃→ 15 ℃, 12 h/1、3、5、9、14、21、28、35次 | 最大田间持水量的60% | 增加 | 增加 |
| 河岸带不同植被类型 | 123° E, 41° N[58] | LT | -5 ℃、-20 ℃→ 5℃, 12 h/0、1、5、10、20、30次 | 原土壤含水量 | 增加 | 农田变化不显著 |
| 天山森林土壤 | 87° E, 43° N[31] | ST | 0 ℃以上→ 0 ℃以下, 3 h/3次 | 原土壤含水量 | 草地和灌丛表现为先减少后增加 | 增加 |
| 大兴安岭落叶松林土壤 | 121° E, 50° N[38] | LT | -5 ℃→ 8 ℃, 24 h/1、3、5、7、15次 | 60%土壤饱和含水量 | 低于恒温条件下的累积量 | 低于恒温条件下的累积量 |
| 草地 | 青藏高原高寒草甸区 | 103° E, 32° N[59] | LT | -4 ℃、-20 ℃ → 4℃, 8 h/1、3、5、25、60次 | 原土壤含水量 | 先增后减 | 先增后减 |
| 湿地 | 三江平原沼泽湿地 | 133° E, 47° N[36] | LT | -5 ℃、-25 ℃→ 5 ℃, 1 d/1、2、4、6、10次 | (352.5±1.6) g·kg-1 | 增加 | 增加 |
| 小兴安岭湿地土壤 | 129° E, 48° N[60] | LT | -20 ℃→ 5 ℃, 24 h/5次 | 原土壤含水量 | 先增后减 | 先减后增 |
| 128° E, 48° N[61] | LT | -5 ℃、-25 ℃→ 5 ℃, 24 h/0、1、2、4、9次 | 原土壤含水量 | 先增后减 | 先减后增 |
注:LT:室内模拟试验;ST:野外原位试验. ...
Study on the process of soil carbon and nitrogen mineralization in cold-temperate coniferous forests under freezing and thawing
1
2011
... 大兴安岭多年冻土泥炭地位于欧亚大陆多年冻土区的南缘,由于纬度高,并伴随着昼夜温差大的气候特点,是全球变暖响应敏感的地区之一,该区多年冻土泥炭地具有湿冷、滞水、厚度浅、稳定性较差等特点[37].近年来,由于全球变暖,该区冻土退化、水位下降,群落结构发生变化,从而导致生产力发生变化,必然会对土壤无机氮的时空变化产生影响.目前关于大兴安岭冻土区的研究主要集中在不同植被类型[38]土壤碳氮矿化[39]、微量元素[40]的分布、温室气体排放[41]等方面.然而,在野外原位探究多年冻土泥炭地无机氮对秋季冻融的响应方面的研究很是缺乏.因此,本研究以大兴安岭三种多年冻土泥炭地为研究对象,旨在探明秋季冻融期间土壤温度和含水量变化对浅层和深层多年冻土泥炭土壤无机氮动态的影响,也为进一步探究该过程中温室气体(N2O)排放对全球变暖响应的研究提供基础数据. ...
气候变化条件下东北地区多年冻土变化预测
1
2011
... 大兴安岭多年冻土泥炭地位于欧亚大陆多年冻土区的南缘,由于纬度高,并伴随着昼夜温差大的气候特点,是全球变暖响应敏感的地区之一,该区多年冻土泥炭地具有湿冷、滞水、厚度浅、稳定性较差等特点[37].近年来,由于全球变暖,该区冻土退化、水位下降,群落结构发生变化,从而导致生产力发生变化,必然会对土壤无机氮的时空变化产生影响.目前关于大兴安岭冻土区的研究主要集中在不同植被类型[38]土壤碳氮矿化[39]、微量元素[40]的分布、温室气体排放[41]等方面.然而,在野外原位探究多年冻土泥炭地无机氮对秋季冻融的响应方面的研究很是缺乏.因此,本研究以大兴安岭三种多年冻土泥炭地为研究对象,旨在探明秋季冻融期间土壤温度和含水量变化对浅层和深层多年冻土泥炭土壤无机氮动态的影响,也为进一步探究该过程中温室气体(N2O)排放对全球变暖响应的研究提供基础数据. ...
Soil carbon and nitrogen mineralization in a Larix gmelinii forest during freeze-thaw cycles
2
2014
... 大兴安岭多年冻土泥炭地位于欧亚大陆多年冻土区的南缘,由于纬度高,并伴随着昼夜温差大的气候特点,是全球变暖响应敏感的地区之一,该区多年冻土泥炭地具有湿冷、滞水、厚度浅、稳定性较差等特点[37].近年来,由于全球变暖,该区冻土退化、水位下降,群落结构发生变化,从而导致生产力发生变化,必然会对土壤无机氮的时空变化产生影响.目前关于大兴安岭冻土区的研究主要集中在不同植被类型[38]土壤碳氮矿化[39]、微量元素[40]的分布、温室气体排放[41]等方面.然而,在野外原位探究多年冻土泥炭地无机氮对秋季冻融的响应方面的研究很是缺乏.因此,本研究以大兴安岭三种多年冻土泥炭地为研究对象,旨在探明秋季冻融期间土壤温度和含水量变化对浅层和深层多年冻土泥炭土壤无机氮动态的影响,也为进一步探究该过程中温室气体(N2O)排放对全球变暖响应的研究提供基础数据. ...
... Comparisons of inorganic nitrogen changes under controlled hydrothermal conditions
Table 5| 生态系统 | 研究地点 | 经纬度 | 实验类型 | 冻融处理/次数 | 含水量 | NH4+-N | NO3--N |
|---|
| 森林 | 长白山地区温带森林土壤 | 127° E, 42° N[34] | LT | -10 ℃→10 ℃, 12 h/1、3、5、7、11、15、22、28、36次 | 秋季低含水量(40%) | 不变 | 增加 |
| 127° E, 42° N[57] | LT | -20 ℃→ 15 ℃, 12 h/1、3、5、9、14、21、28、35次 | 最大田间持水量的60% | 增加 | 增加 |
| 河岸带不同植被类型 | 123° E, 41° N[58] | LT | -5 ℃、-20 ℃→ 5℃, 12 h/0、1、5、10、20、30次 | 原土壤含水量 | 增加 | 农田变化不显著 |
| 天山森林土壤 | 87° E, 43° N[31] | ST | 0 ℃以上→ 0 ℃以下, 3 h/3次 | 原土壤含水量 | 草地和灌丛表现为先减少后增加 | 增加 |
| 大兴安岭落叶松林土壤 | 121° E, 50° N[38] | LT | -5 ℃→ 8 ℃, 24 h/1、3、5、7、15次 | 60%土壤饱和含水量 | 低于恒温条件下的累积量 | 低于恒温条件下的累积量 |
| 草地 | 青藏高原高寒草甸区 | 103° E, 32° N[59] | LT | -4 ℃、-20 ℃ → 4℃, 8 h/1、3、5、25、60次 | 原土壤含水量 | 先增后减 | 先增后减 |
| 湿地 | 三江平原沼泽湿地 | 133° E, 47° N[36] | LT | -5 ℃、-25 ℃→ 5 ℃, 1 d/1、2、4、6、10次 | (352.5±1.6) g·kg-1 | 增加 | 增加 |
| 小兴安岭湿地土壤 | 129° E, 48° N[60] | LT | -20 ℃→ 5 ℃, 24 h/5次 | 原土壤含水量 | 先增后减 | 先减后增 |
| 128° E, 48° N[61] | LT | -5 ℃、-25 ℃→ 5 ℃, 24 h/0、1、2、4、9次 | 原土壤含水量 | 先增后减 | 先减后增 |
注:LT:室内模拟试验;ST:野外原位试验. ...
冻融作用下寒温带针叶林土壤碳氮矿化过程研究
2
2014
... 大兴安岭多年冻土泥炭地位于欧亚大陆多年冻土区的南缘,由于纬度高,并伴随着昼夜温差大的气候特点,是全球变暖响应敏感的地区之一,该区多年冻土泥炭地具有湿冷、滞水、厚度浅、稳定性较差等特点[37].近年来,由于全球变暖,该区冻土退化、水位下降,群落结构发生变化,从而导致生产力发生变化,必然会对土壤无机氮的时空变化产生影响.目前关于大兴安岭冻土区的研究主要集中在不同植被类型[38]土壤碳氮矿化[39]、微量元素[40]的分布、温室气体排放[41]等方面.然而,在野外原位探究多年冻土泥炭地无机氮对秋季冻融的响应方面的研究很是缺乏.因此,本研究以大兴安岭三种多年冻土泥炭地为研究对象,旨在探明秋季冻融期间土壤温度和含水量变化对浅层和深层多年冻土泥炭土壤无机氮动态的影响,也为进一步探究该过程中温室气体(N2O)排放对全球变暖响应的研究提供基础数据. ...
... Comparisons of inorganic nitrogen changes under controlled hydrothermal conditions
Table 5| 生态系统 | 研究地点 | 经纬度 | 实验类型 | 冻融处理/次数 | 含水量 | NH4+-N | NO3--N |
|---|
| 森林 | 长白山地区温带森林土壤 | 127° E, 42° N[34] | LT | -10 ℃→10 ℃, 12 h/1、3、5、7、11、15、22、28、36次 | 秋季低含水量(40%) | 不变 | 增加 |
| 127° E, 42° N[57] | LT | -20 ℃→ 15 ℃, 12 h/1、3、5、9、14、21、28、35次 | 最大田间持水量的60% | 增加 | 增加 |
| 河岸带不同植被类型 | 123° E, 41° N[58] | LT | -5 ℃、-20 ℃→ 5℃, 12 h/0、1、5、10、20、30次 | 原土壤含水量 | 增加 | 农田变化不显著 |
| 天山森林土壤 | 87° E, 43° N[31] | ST | 0 ℃以上→ 0 ℃以下, 3 h/3次 | 原土壤含水量 | 草地和灌丛表现为先减少后增加 | 增加 |
| 大兴安岭落叶松林土壤 | 121° E, 50° N[38] | LT | -5 ℃→ 8 ℃, 24 h/1、3、5、7、15次 | 60%土壤饱和含水量 | 低于恒温条件下的累积量 | 低于恒温条件下的累积量 |
| 草地 | 青藏高原高寒草甸区 | 103° E, 32° N[59] | LT | -4 ℃、-20 ℃ → 4℃, 8 h/1、3、5、25、60次 | 原土壤含水量 | 先增后减 | 先增后减 |
| 湿地 | 三江平原沼泽湿地 | 133° E, 47° N[36] | LT | -5 ℃、-25 ℃→ 5 ℃, 1 d/1、2、4、6、10次 | (352.5±1.6) g·kg-1 | 增加 | 增加 |
| 小兴安岭湿地土壤 | 129° E, 48° N[60] | LT | -20 ℃→ 5 ℃, 24 h/5次 | 原土壤含水量 | 先增后减 | 先减后增 |
| 128° E, 48° N[61] | LT | -5 ℃、-25 ℃→ 5 ℃, 24 h/0、1、2、4、9次 | 原土壤含水量 | 先增后减 | 先减后增 |
注:LT:室内模拟试验;ST:野外原位试验. ...
Effects of freezing-thawing cycles on soil organic carbon mineralization in the peatland ecosystems from continuous permafrost zone, Great Hinggan Mountains
1
2018
... 大兴安岭多年冻土泥炭地位于欧亚大陆多年冻土区的南缘,由于纬度高,并伴随着昼夜温差大的气候特点,是全球变暖响应敏感的地区之一,该区多年冻土泥炭地具有湿冷、滞水、厚度浅、稳定性较差等特点[37].近年来,由于全球变暖,该区冻土退化、水位下降,群落结构发生变化,从而导致生产力发生变化,必然会对土壤无机氮的时空变化产生影响.目前关于大兴安岭冻土区的研究主要集中在不同植被类型[38]土壤碳氮矿化[39]、微量元素[40]的分布、温室气体排放[41]等方面.然而,在野外原位探究多年冻土泥炭地无机氮对秋季冻融的响应方面的研究很是缺乏.因此,本研究以大兴安岭三种多年冻土泥炭地为研究对象,旨在探明秋季冻融期间土壤温度和含水量变化对浅层和深层多年冻土泥炭土壤无机氮动态的影响,也为进一步探究该过程中温室气体(N2O)排放对全球变暖响应的研究提供基础数据. ...
冻融作用对大兴安岭多年冻土区泥炭地土壤有机碳矿化的影响研究
1
2018
... 大兴安岭多年冻土泥炭地位于欧亚大陆多年冻土区的南缘,由于纬度高,并伴随着昼夜温差大的气候特点,是全球变暖响应敏感的地区之一,该区多年冻土泥炭地具有湿冷、滞水、厚度浅、稳定性较差等特点[37].近年来,由于全球变暖,该区冻土退化、水位下降,群落结构发生变化,从而导致生产力发生变化,必然会对土壤无机氮的时空变化产生影响.目前关于大兴安岭冻土区的研究主要集中在不同植被类型[38]土壤碳氮矿化[39]、微量元素[40]的分布、温室气体排放[41]等方面.然而,在野外原位探究多年冻土泥炭地无机氮对秋季冻融的响应方面的研究很是缺乏.因此,本研究以大兴安岭三种多年冻土泥炭地为研究对象,旨在探明秋季冻融期间土壤温度和含水量变化对浅层和深层多年冻土泥炭土壤无机氮动态的影响,也为进一步探究该过程中温室气体(N2O)排放对全球变暖响应的研究提供基础数据. ...
Trace element dispersion characteristics of peat in the Da Hingan Mountains and environmental significance
1
1997
... 大兴安岭多年冻土泥炭地位于欧亚大陆多年冻土区的南缘,由于纬度高,并伴随着昼夜温差大的气候特点,是全球变暖响应敏感的地区之一,该区多年冻土泥炭地具有湿冷、滞水、厚度浅、稳定性较差等特点[37].近年来,由于全球变暖,该区冻土退化、水位下降,群落结构发生变化,从而导致生产力发生变化,必然会对土壤无机氮的时空变化产生影响.目前关于大兴安岭冻土区的研究主要集中在不同植被类型[38]土壤碳氮矿化[39]、微量元素[40]的分布、温室气体排放[41]等方面.然而,在野外原位探究多年冻土泥炭地无机氮对秋季冻融的响应方面的研究很是缺乏.因此,本研究以大兴安岭三种多年冻土泥炭地为研究对象,旨在探明秋季冻融期间土壤温度和含水量变化对浅层和深层多年冻土泥炭土壤无机氮动态的影响,也为进一步探究该过程中温室气体(N2O)排放对全球变暖响应的研究提供基础数据. ...
大兴安岭泥炭微量元素分布特征及其环境意义
1
1997
... 大兴安岭多年冻土泥炭地位于欧亚大陆多年冻土区的南缘,由于纬度高,并伴随着昼夜温差大的气候特点,是全球变暖响应敏感的地区之一,该区多年冻土泥炭地具有湿冷、滞水、厚度浅、稳定性较差等特点[37].近年来,由于全球变暖,该区冻土退化、水位下降,群落结构发生变化,从而导致生产力发生变化,必然会对土壤无机氮的时空变化产生影响.目前关于大兴安岭冻土区的研究主要集中在不同植被类型[38]土壤碳氮矿化[39]、微量元素[40]的分布、温室气体排放[41]等方面.然而,在野外原位探究多年冻土泥炭地无机氮对秋季冻融的响应方面的研究很是缺乏.因此,本研究以大兴安岭三种多年冻土泥炭地为研究对象,旨在探明秋季冻融期间土壤温度和含水量变化对浅层和深层多年冻土泥炭土壤无机氮动态的影响,也为进一步探究该过程中温室气体(N2O)排放对全球变暖响应的研究提供基础数据. ...
Characteristics of greenhouse gas emissions from seven swamp types in the permafrost region of Daxing’ an Mountains, northeast China
1
2020
... 大兴安岭多年冻土泥炭地位于欧亚大陆多年冻土区的南缘,由于纬度高,并伴随着昼夜温差大的气候特点,是全球变暖响应敏感的地区之一,该区多年冻土泥炭地具有湿冷、滞水、厚度浅、稳定性较差等特点[37].近年来,由于全球变暖,该区冻土退化、水位下降,群落结构发生变化,从而导致生产力发生变化,必然会对土壤无机氮的时空变化产生影响.目前关于大兴安岭冻土区的研究主要集中在不同植被类型[38]土壤碳氮矿化[39]、微量元素[40]的分布、温室气体排放[41]等方面.然而,在野外原位探究多年冻土泥炭地无机氮对秋季冻融的响应方面的研究很是缺乏.因此,本研究以大兴安岭三种多年冻土泥炭地为研究对象,旨在探明秋季冻融期间土壤温度和含水量变化对浅层和深层多年冻土泥炭土壤无机氮动态的影响,也为进一步探究该过程中温室气体(N2O)排放对全球变暖响应的研究提供基础数据. ...
大兴安岭永久冻土区7种沼泽类型土壤温室气体排放特征
1
2020
... 大兴安岭多年冻土泥炭地位于欧亚大陆多年冻土区的南缘,由于纬度高,并伴随着昼夜温差大的气候特点,是全球变暖响应敏感的地区之一,该区多年冻土泥炭地具有湿冷、滞水、厚度浅、稳定性较差等特点[37].近年来,由于全球变暖,该区冻土退化、水位下降,群落结构发生变化,从而导致生产力发生变化,必然会对土壤无机氮的时空变化产生影响.目前关于大兴安岭冻土区的研究主要集中在不同植被类型[38]土壤碳氮矿化[39]、微量元素[40]的分布、温室气体排放[41]等方面.然而,在野外原位探究多年冻土泥炭地无机氮对秋季冻融的响应方面的研究很是缺乏.因此,本研究以大兴安岭三种多年冻土泥炭地为研究对象,旨在探明秋季冻融期间土壤温度和含水量变化对浅层和深层多年冻土泥炭土壤无机氮动态的影响,也为进一步探究该过程中温室气体(N2O)排放对全球变暖响应的研究提供基础数据. ...
Non-growing-season soil respiration is controlled by freezing and thawing processes in the summer monsoon-dominated Tibetan alpine grassland
1
2014
... 通过对研究区秋季冻融期大气、土壤温度的连续观测,将研究区最低气温开始位于0 ℃以下(研究区9月中下旬)至最高气温位于0 ℃以下并持续3~5天的(研究区11月上旬)这段时间作为本研究的秋季冻融期[42-43].其中9月25日—10月7日为秋季冻融前期,该地区的大气和土壤温度均呈下降趋势.10月10日—10月22日为秋季冻融中期,日最低气温降至0 ℃,截至10月24日—11月5日为秋季冻融后期,日最高气温和地表温度均低于0 ℃,且持续数天.根据大兴安岭多年冻土泥炭层土壤平均深度,把土壤垂直剖面分为两部分:0~20 cm为浅层泥炭土;20~50 cm为深层泥炭土. ...
Annual ecosystem respiration is resistant to changes in freeze-thaw periods in semi-arid permafrost
1
2020
... 通过对研究区秋季冻融期大气、土壤温度的连续观测,将研究区最低气温开始位于0 ℃以下(研究区9月中下旬)至最高气温位于0 ℃以下并持续3~5天的(研究区11月上旬)这段时间作为本研究的秋季冻融期[42-43].其中9月25日—10月7日为秋季冻融前期,该地区的大气和土壤温度均呈下降趋势.10月10日—10月22日为秋季冻融中期,日最低气温降至0 ℃,截至10月24日—11月5日为秋季冻融后期,日最高气温和地表温度均低于0 ℃,且持续数天.根据大兴安岭多年冻土泥炭层土壤平均深度,把土壤垂直剖面分为两部分:0~20 cm为浅层泥炭土;20~50 cm为深层泥炭土. ...
Indoor simulation study on carbon and nitrogen contents and enzyme activities of soils in permafrost region in Greater Khingan Mountains
2
2018
... 本研究中,3种多年冻土泥炭地土壤NH4+-N、NO3--N含量的变化范围分别为:(1.00±0.00)~(20.60±0.20) mg·kg-1、(0.02±0.01)~(14.64±1.11) mg·kg-1,较该区域7月(98.43~216 mg·kg-1、15.58~17.07 mg·kg-1)变化明显降低[44],说明秋季冻融期温度变化会影响北方泥炭地土壤无机氮的动态.但是也有研究发现秋季温度变化对冻土区无机氮含量没有影响[45],其原因可能是土壤在不同的生态系统中,产生了对环境和温度耐受力不同的土壤微生物,而土壤微生物是影响土壤中无机氮变化的关键因素[46],从而使无机氮变化相对复杂. ...
... 经逐步回归分析表明(表2~4),在不同冻融阶段,无机氮对环境因子的响应程度也存在差异.在整个秋季冻融期,BM浅层(10~20 cm)含水量对无机氮含量的影响程度最大(R2=0.301),这是由于浅层土壤含水量更容易影响土壤通气性和土壤微生物活性,进而改变影响了土壤基质的养分循环,从而对无机氮转化产生影响.通常认为无机氮对温度的响应程度大于含水量[62],但是BM样地在整个秋季冻融时期,温度和含水量都处于较低水平且变化幅度较小,与温度相比含水量对土壤微生物活性起有效作用,这与赵媛[34]在研究秋末冻融循环对长白山地区无机氮的影响结果相似,即无机氮对含水量的响应程度大于温度.在秋季冻融前、中和后期,3种多年冻土泥炭地浅层(0~20 cm)土壤的环境因子与无机氮释放有关,其中浅层土壤温度对无机氮释放的贡献率最大(R2=0.685),这主要受当地非生长季寒冷气候的影响,温度通过改变土壤中生物酶和氨氧化细菌、硝化细菌的活性来影响氮的分解和无机氮的释放[63].但是不同土壤环境条件下微生物对温度敏感程度有很大差异[64],本研究发现,受非生长季温度逐渐降低的影响,XA仅在秋季冻融前期土壤温度对无机氮释放有贡献(R2=0.685),在秋季冻融后期不存在相关关系,这与表5中草地生态系统下青藏高原高寒草甸区研究发现短期的冻融交替可以显著增加土壤中的氮矿化累积结果相似.说明与长期冻融相比,短期温度变化会破坏土壤团聚结构从而影响氨氧化细菌的活性,一部分微生物为了维持繁殖不断汲取死亡微生物的养分,从而促进了微生物的活性,影响土壤氮矿化速度.但也有研究发现,短期冻融对土壤氮矿化并没有影响[57],土壤微生物会对温度变化做出调节反应,从而无机氮变化不显著.秋季冻融期无机氮对环境因子的响应程度存在以上差异的原因可能有以下四方面:第一,样地沿一定坡度选取,地势高低不同造成了土壤含水量存在显著差异.第二,受到地表植被的影响,土壤微生物的类别和数量存在差异因此对氮矿化程度不同[44].第三,与凋落物有关,不同凋落物分解的有机质不同会影响土壤有效基质[65].第四,3种泥炭地土壤的容重、孔隙度和pH值等物理性质存在差异[66].这些要素的综合影响造成了不同泥炭地间,以及冻融期不同时期间无机氮含量对温度和含水量响应程度的不同. ...
大兴安岭冻土区泥炭地土壤碳、氮含量和酶活性室内模拟研究
2
2018
... 本研究中,3种多年冻土泥炭地土壤NH4+-N、NO3--N含量的变化范围分别为:(1.00±0.00)~(20.60±0.20) mg·kg-1、(0.02±0.01)~(14.64±1.11) mg·kg-1,较该区域7月(98.43~216 mg·kg-1、15.58~17.07 mg·kg-1)变化明显降低[44],说明秋季冻融期温度变化会影响北方泥炭地土壤无机氮的动态.但是也有研究发现秋季温度变化对冻土区无机氮含量没有影响[45],其原因可能是土壤在不同的生态系统中,产生了对环境和温度耐受力不同的土壤微生物,而土壤微生物是影响土壤中无机氮变化的关键因素[46],从而使无机氮变化相对复杂. ...
... 经逐步回归分析表明(表2~4),在不同冻融阶段,无机氮对环境因子的响应程度也存在差异.在整个秋季冻融期,BM浅层(10~20 cm)含水量对无机氮含量的影响程度最大(R2=0.301),这是由于浅层土壤含水量更容易影响土壤通气性和土壤微生物活性,进而改变影响了土壤基质的养分循环,从而对无机氮转化产生影响.通常认为无机氮对温度的响应程度大于含水量[62],但是BM样地在整个秋季冻融时期,温度和含水量都处于较低水平且变化幅度较小,与温度相比含水量对土壤微生物活性起有效作用,这与赵媛[34]在研究秋末冻融循环对长白山地区无机氮的影响结果相似,即无机氮对含水量的响应程度大于温度.在秋季冻融前、中和后期,3种多年冻土泥炭地浅层(0~20 cm)土壤的环境因子与无机氮释放有关,其中浅层土壤温度对无机氮释放的贡献率最大(R2=0.685),这主要受当地非生长季寒冷气候的影响,温度通过改变土壤中生物酶和氨氧化细菌、硝化细菌的活性来影响氮的分解和无机氮的释放[63].但是不同土壤环境条件下微生物对温度敏感程度有很大差异[64],本研究发现,受非生长季温度逐渐降低的影响,XA仅在秋季冻融前期土壤温度对无机氮释放有贡献(R2=0.685),在秋季冻融后期不存在相关关系,这与表5中草地生态系统下青藏高原高寒草甸区研究发现短期的冻融交替可以显著增加土壤中的氮矿化累积结果相似.说明与长期冻融相比,短期温度变化会破坏土壤团聚结构从而影响氨氧化细菌的活性,一部分微生物为了维持繁殖不断汲取死亡微生物的养分,从而促进了微生物的活性,影响土壤氮矿化速度.但也有研究发现,短期冻融对土壤氮矿化并没有影响[57],土壤微生物会对温度变化做出调节反应,从而无机氮变化不显著.秋季冻融期无机氮对环境因子的响应程度存在以上差异的原因可能有以下四方面:第一,样地沿一定坡度选取,地势高低不同造成了土壤含水量存在显著差异.第二,受到地表植被的影响,土壤微生物的类别和数量存在差异因此对氮矿化程度不同[44].第三,与凋落物有关,不同凋落物分解的有机质不同会影响土壤有效基质[65].第四,3种泥炭地土壤的容重、孔隙度和pH值等物理性质存在差异[66].这些要素的综合影响造成了不同泥炭地间,以及冻融期不同时期间无机氮含量对温度和含水量响应程度的不同. ...
Effects of mild winter freezing on soil nitrogen and carbon dynamics in a northern hardwood forest
1
2001
... 本研究中,3种多年冻土泥炭地土壤NH4+-N、NO3--N含量的变化范围分别为:(1.00±0.00)~(20.60±0.20) mg·kg-1、(0.02±0.01)~(14.64±1.11) mg·kg-1,较该区域7月(98.43~216 mg·kg-1、15.58~17.07 mg·kg-1)变化明显降低[44],说明秋季冻融期温度变化会影响北方泥炭地土壤无机氮的动态.但是也有研究发现秋季温度变化对冻土区无机氮含量没有影响[45],其原因可能是土壤在不同的生态系统中,产生了对环境和温度耐受力不同的土壤微生物,而土壤微生物是影响土壤中无机氮变化的关键因素[46],从而使无机氮变化相对复杂. ...
The influence of hydrothermal conditions change on nitrogen transformation and enzyme activities in black soil in Northeast China
1
2015
... 本研究中,3种多年冻土泥炭地土壤NH4+-N、NO3--N含量的变化范围分别为:(1.00±0.00)~(20.60±0.20) mg·kg-1、(0.02±0.01)~(14.64±1.11) mg·kg-1,较该区域7月(98.43~216 mg·kg-1、15.58~17.07 mg·kg-1)变化明显降低[44],说明秋季冻融期温度变化会影响北方泥炭地土壤无机氮的动态.但是也有研究发现秋季温度变化对冻土区无机氮含量没有影响[45],其原因可能是土壤在不同的生态系统中,产生了对环境和温度耐受力不同的土壤微生物,而土壤微生物是影响土壤中无机氮变化的关键因素[46],从而使无机氮变化相对复杂. ...
东北黑土氮素转化和酶活性对水热条件变化的响应
1
2015
... 本研究中,3种多年冻土泥炭地土壤NH4+-N、NO3--N含量的变化范围分别为:(1.00±0.00)~(20.60±0.20) mg·kg-1、(0.02±0.01)~(14.64±1.11) mg·kg-1,较该区域7月(98.43~216 mg·kg-1、15.58~17.07 mg·kg-1)变化明显降低[44],说明秋季冻融期温度变化会影响北方泥炭地土壤无机氮的动态.但是也有研究发现秋季温度变化对冻土区无机氮含量没有影响[45],其原因可能是土壤在不同的生态系统中,产生了对环境和温度耐受力不同的土壤微生物,而土壤微生物是影响土壤中无机氮变化的关键因素[46],从而使无机氮变化相对复杂. ...
Soil nitrogen mineralization characteristics of the natural coniferous forest in northern Daxing’an Mountains, Northeast China
1
2019
... 本研究发现大兴安岭冻土区土壤有效氮以NH4+-N为主,这与肖瑞晗在研究本区域结果相一致[47].由于NO3--N携带被土壤胶体排斥的负电荷[48],一方面在非生长季受降水等影响向较深土壤淋溶,另一方面受到反硝化作用以挥发方式损失,从而导致土壤中NH4+-N明显高于NO3--N.本研究中,XA样地土壤浅层无机氮的含量明显高于深层,这一结果与鲁博权等[49]研究结果相似,即无机氮在垂直方向上具有空间异质性.这是由于冻土区植被根系在汲取氮素时会使有机质在浅层聚集,且浅层土壤因水热条件和土壤基质有效性条件好,氮矿化程度高于微生物的固持[50],而深层土壤则相反,从而导致无机氮含量较低.我们发现在整个秋季冻融时期,土壤NH4+-N在3种冻土泥炭地各土层间不存在显著差异(P>0.05);而NO3--N在浅层和深层土壤中呈现出显著性差异(P<0.05),其中XY受饱和含水量的影响NO3--N平均含量较高,说明较高含水量会促进土壤矿化作用.可能是由于这种条件下微生物活性的增强提高了氨化而抑制了硝化作用,使NH4+-N含量不断累积同时不断向NO3--N转换[51].不同泥炭地间无机氮含量以及组分会形成以上差异,说明不同泥炭地间土壤微生物会因为环境因子的不同形成差异,从而导致土壤氮矿化程度不同[52],其次由于不同土层间土壤物理性质(结构、孔隙度)的差异,造成了NO3--N淋溶、NH4+-N吸附程度不同. ...
大兴安岭北部天然针叶林土壤氮矿化特征
1
2019
... 本研究发现大兴安岭冻土区土壤有效氮以NH4+-N为主,这与肖瑞晗在研究本区域结果相一致[47].由于NO3--N携带被土壤胶体排斥的负电荷[48],一方面在非生长季受降水等影响向较深土壤淋溶,另一方面受到反硝化作用以挥发方式损失,从而导致土壤中NH4+-N明显高于NO3--N.本研究中,XA样地土壤浅层无机氮的含量明显高于深层,这一结果与鲁博权等[49]研究结果相似,即无机氮在垂直方向上具有空间异质性.这是由于冻土区植被根系在汲取氮素时会使有机质在浅层聚集,且浅层土壤因水热条件和土壤基质有效性条件好,氮矿化程度高于微生物的固持[50],而深层土壤则相反,从而导致无机氮含量较低.我们发现在整个秋季冻融时期,土壤NH4+-N在3种冻土泥炭地各土层间不存在显著差异(P>0.05);而NO3--N在浅层和深层土壤中呈现出显著性差异(P<0.05),其中XY受饱和含水量的影响NO3--N平均含量较高,说明较高含水量会促进土壤矿化作用.可能是由于这种条件下微生物活性的增强提高了氨化而抑制了硝化作用,使NH4+-N含量不断累积同时不断向NO3--N转换[51].不同泥炭地间无机氮含量以及组分会形成以上差异,说明不同泥炭地间土壤微生物会因为环境因子的不同形成差异,从而导致土壤氮矿化程度不同[52],其次由于不同土层间土壤物理性质(结构、孔隙度)的差异,造成了NO3--N淋溶、NH4+-N吸附程度不同. ...
Effects of temperature on soil nitrogen mineralization in three subalpine forests of western Sichuan, China
1
2017
... 本研究发现大兴安岭冻土区土壤有效氮以NH4+-N为主,这与肖瑞晗在研究本区域结果相一致[47].由于NO3--N携带被土壤胶体排斥的负电荷[48],一方面在非生长季受降水等影响向较深土壤淋溶,另一方面受到反硝化作用以挥发方式损失,从而导致土壤中NH4+-N明显高于NO3--N.本研究中,XA样地土壤浅层无机氮的含量明显高于深层,这一结果与鲁博权等[49]研究结果相似,即无机氮在垂直方向上具有空间异质性.这是由于冻土区植被根系在汲取氮素时会使有机质在浅层聚集,且浅层土壤因水热条件和土壤基质有效性条件好,氮矿化程度高于微生物的固持[50],而深层土壤则相反,从而导致无机氮含量较低.我们发现在整个秋季冻融时期,土壤NH4+-N在3种冻土泥炭地各土层间不存在显著差异(P>0.05);而NO3--N在浅层和深层土壤中呈现出显著性差异(P<0.05),其中XY受饱和含水量的影响NO3--N平均含量较高,说明较高含水量会促进土壤矿化作用.可能是由于这种条件下微生物活性的增强提高了氨化而抑制了硝化作用,使NH4+-N含量不断累积同时不断向NO3--N转换[51].不同泥炭地间无机氮含量以及组分会形成以上差异,说明不同泥炭地间土壤微生物会因为环境因子的不同形成差异,从而导致土壤氮矿化程度不同[52],其次由于不同土层间土壤物理性质(结构、孔隙度)的差异,造成了NO3--N淋溶、NH4+-N吸附程度不同. ...
温度对川西亚高山3种森林土壤氮矿化的影响
1
2017
... 本研究发现大兴安岭冻土区土壤有效氮以NH4+-N为主,这与肖瑞晗在研究本区域结果相一致[47].由于NO3--N携带被土壤胶体排斥的负电荷[48],一方面在非生长季受降水等影响向较深土壤淋溶,另一方面受到反硝化作用以挥发方式损失,从而导致土壤中NH4+-N明显高于NO3--N.本研究中,XA样地土壤浅层无机氮的含量明显高于深层,这一结果与鲁博权等[49]研究结果相似,即无机氮在垂直方向上具有空间异质性.这是由于冻土区植被根系在汲取氮素时会使有机质在浅层聚集,且浅层土壤因水热条件和土壤基质有效性条件好,氮矿化程度高于微生物的固持[50],而深层土壤则相反,从而导致无机氮含量较低.我们发现在整个秋季冻融时期,土壤NH4+-N在3种冻土泥炭地各土层间不存在显著差异(P>0.05);而NO3--N在浅层和深层土壤中呈现出显著性差异(P<0.05),其中XY受饱和含水量的影响NO3--N平均含量较高,说明较高含水量会促进土壤矿化作用.可能是由于这种条件下微生物活性的增强提高了氨化而抑制了硝化作用,使NH4+-N含量不断累积同时不断向NO3--N转换[51].不同泥炭地间无机氮含量以及组分会形成以上差异,说明不同泥炭地间土壤微生物会因为环境因子的不同形成差异,从而导致土壤氮矿化程度不同[52],其次由于不同土层间土壤物理性质(结构、孔隙度)的差异,造成了NO3--N淋溶、NH4+-N吸附程度不同. ...
The effects of freezing-thawing process on soil active organic carbon and nitrogen mineralization in Daxing’ an Mountains forests
1
2019
... 本研究发现大兴安岭冻土区土壤有效氮以NH4+-N为主,这与肖瑞晗在研究本区域结果相一致[47].由于NO3--N携带被土壤胶体排斥的负电荷[48],一方面在非生长季受降水等影响向较深土壤淋溶,另一方面受到反硝化作用以挥发方式损失,从而导致土壤中NH4+-N明显高于NO3--N.本研究中,XA样地土壤浅层无机氮的含量明显高于深层,这一结果与鲁博权等[49]研究结果相似,即无机氮在垂直方向上具有空间异质性.这是由于冻土区植被根系在汲取氮素时会使有机质在浅层聚集,且浅层土壤因水热条件和土壤基质有效性条件好,氮矿化程度高于微生物的固持[50],而深层土壤则相反,从而导致无机氮含量较低.我们发现在整个秋季冻融时期,土壤NH4+-N在3种冻土泥炭地各土层间不存在显著差异(P>0.05);而NO3--N在浅层和深层土壤中呈现出显著性差异(P<0.05),其中XY受饱和含水量的影响NO3--N平均含量较高,说明较高含水量会促进土壤矿化作用.可能是由于这种条件下微生物活性的增强提高了氨化而抑制了硝化作用,使NH4+-N含量不断累积同时不断向NO3--N转换[51].不同泥炭地间无机氮含量以及组分会形成以上差异,说明不同泥炭地间土壤微生物会因为环境因子的不同形成差异,从而导致土壤氮矿化程度不同[52],其次由于不同土层间土壤物理性质(结构、孔隙度)的差异,造成了NO3--N淋溶、NH4+-N吸附程度不同. ...
冻融作用对大兴安岭典型森林土壤活性有机碳和氮矿化的影响
1
2019
... 本研究发现大兴安岭冻土区土壤有效氮以NH4+-N为主,这与肖瑞晗在研究本区域结果相一致[47].由于NO3--N携带被土壤胶体排斥的负电荷[48],一方面在非生长季受降水等影响向较深土壤淋溶,另一方面受到反硝化作用以挥发方式损失,从而导致土壤中NH4+-N明显高于NO3--N.本研究中,XA样地土壤浅层无机氮的含量明显高于深层,这一结果与鲁博权等[49]研究结果相似,即无机氮在垂直方向上具有空间异质性.这是由于冻土区植被根系在汲取氮素时会使有机质在浅层聚集,且浅层土壤因水热条件和土壤基质有效性条件好,氮矿化程度高于微生物的固持[50],而深层土壤则相反,从而导致无机氮含量较低.我们发现在整个秋季冻融时期,土壤NH4+-N在3种冻土泥炭地各土层间不存在显著差异(P>0.05);而NO3--N在浅层和深层土壤中呈现出显著性差异(P<0.05),其中XY受饱和含水量的影响NO3--N平均含量较高,说明较高含水量会促进土壤矿化作用.可能是由于这种条件下微生物活性的增强提高了氨化而抑制了硝化作用,使NH4+-N含量不断累积同时不断向NO3--N转换[51].不同泥炭地间无机氮含量以及组分会形成以上差异,说明不同泥炭地间土壤微生物会因为环境因子的不同形成差异,从而导致土壤氮矿化程度不同[52],其次由于不同土层间土壤物理性质(结构、孔隙度)的差异,造成了NO3--N淋溶、NH4+-N吸附程度不同. ...
Effects of experimental warming on soil nitrogen transformation in alpine scrubland of eastern Qinghai-Tibet Plateau, China
2
2021
... 本研究发现大兴安岭冻土区土壤有效氮以NH4+-N为主,这与肖瑞晗在研究本区域结果相一致[47].由于NO3--N携带被土壤胶体排斥的负电荷[48],一方面在非生长季受降水等影响向较深土壤淋溶,另一方面受到反硝化作用以挥发方式损失,从而导致土壤中NH4+-N明显高于NO3--N.本研究中,XA样地土壤浅层无机氮的含量明显高于深层,这一结果与鲁博权等[49]研究结果相似,即无机氮在垂直方向上具有空间异质性.这是由于冻土区植被根系在汲取氮素时会使有机质在浅层聚集,且浅层土壤因水热条件和土壤基质有效性条件好,氮矿化程度高于微生物的固持[50],而深层土壤则相反,从而导致无机氮含量较低.我们发现在整个秋季冻融时期,土壤NH4+-N在3种冻土泥炭地各土层间不存在显著差异(P>0.05);而NO3--N在浅层和深层土壤中呈现出显著性差异(P<0.05),其中XY受饱和含水量的影响NO3--N平均含量较高,说明较高含水量会促进土壤矿化作用.可能是由于这种条件下微生物活性的增强提高了氨化而抑制了硝化作用,使NH4+-N含量不断累积同时不断向NO3--N转换[51].不同泥炭地间无机氮含量以及组分会形成以上差异,说明不同泥炭地间土壤微生物会因为环境因子的不同形成差异,从而导致土壤氮矿化程度不同[52],其次由于不同土层间土壤物理性质(结构、孔隙度)的差异,造成了NO3--N淋溶、NH4+-N吸附程度不同. ...
... 有研究认为单一的温度[50]或水分[55]条件会对无机氮造成影响,本实验发现在秋季冻融时期土壤温度和含水量与无机氮含量之间存在相关关系,但是与水热的交互作用之间不存在相关性,这与马秀艳[56]在研究该区结果不同,产生差异的原因可能是因为野外原位实验与室内模拟实验条件下,环境因子变化的复杂程度不同,造成了无机氮与水热交互作用的相关性存在差异. ...
模拟增温对青藏高原东部高寒灌丛土壤氮转化的影响
2
2021
... 本研究发现大兴安岭冻土区土壤有效氮以NH4+-N为主,这与肖瑞晗在研究本区域结果相一致[47].由于NO3--N携带被土壤胶体排斥的负电荷[48],一方面在非生长季受降水等影响向较深土壤淋溶,另一方面受到反硝化作用以挥发方式损失,从而导致土壤中NH4+-N明显高于NO3--N.本研究中,XA样地土壤浅层无机氮的含量明显高于深层,这一结果与鲁博权等[49]研究结果相似,即无机氮在垂直方向上具有空间异质性.这是由于冻土区植被根系在汲取氮素时会使有机质在浅层聚集,且浅层土壤因水热条件和土壤基质有效性条件好,氮矿化程度高于微生物的固持[50],而深层土壤则相反,从而导致无机氮含量较低.我们发现在整个秋季冻融时期,土壤NH4+-N在3种冻土泥炭地各土层间不存在显著差异(P>0.05);而NO3--N在浅层和深层土壤中呈现出显著性差异(P<0.05),其中XY受饱和含水量的影响NO3--N平均含量较高,说明较高含水量会促进土壤矿化作用.可能是由于这种条件下微生物活性的增强提高了氨化而抑制了硝化作用,使NH4+-N含量不断累积同时不断向NO3--N转换[51].不同泥炭地间无机氮含量以及组分会形成以上差异,说明不同泥炭地间土壤微生物会因为环境因子的不同形成差异,从而导致土壤氮矿化程度不同[52],其次由于不同土层间土壤物理性质(结构、孔隙度)的差异,造成了NO3--N淋溶、NH4+-N吸附程度不同. ...
... 有研究认为单一的温度[50]或水分[55]条件会对无机氮造成影响,本实验发现在秋季冻融时期土壤温度和含水量与无机氮含量之间存在相关关系,但是与水热的交互作用之间不存在相关性,这与马秀艳[56]在研究该区结果不同,产生差异的原因可能是因为野外原位实验与室内模拟实验条件下,环境因子变化的复杂程度不同,造成了无机氮与水热交互作用的相关性存在差异. ...
Effect of freezing-thawing on nitrogen mineralization in vegetation soils of four landscape zones of Changbai Mountain
1
2011
... 本研究发现大兴安岭冻土区土壤有效氮以NH4+-N为主,这与肖瑞晗在研究本区域结果相一致[47].由于NO3--N携带被土壤胶体排斥的负电荷[48],一方面在非生长季受降水等影响向较深土壤淋溶,另一方面受到反硝化作用以挥发方式损失,从而导致土壤中NH4+-N明显高于NO3--N.本研究中,XA样地土壤浅层无机氮的含量明显高于深层,这一结果与鲁博权等[49]研究结果相似,即无机氮在垂直方向上具有空间异质性.这是由于冻土区植被根系在汲取氮素时会使有机质在浅层聚集,且浅层土壤因水热条件和土壤基质有效性条件好,氮矿化程度高于微生物的固持[50],而深层土壤则相反,从而导致无机氮含量较低.我们发现在整个秋季冻融时期,土壤NH4+-N在3种冻土泥炭地各土层间不存在显著差异(P>0.05);而NO3--N在浅层和深层土壤中呈现出显著性差异(P<0.05),其中XY受饱和含水量的影响NO3--N平均含量较高,说明较高含水量会促进土壤矿化作用.可能是由于这种条件下微生物活性的增强提高了氨化而抑制了硝化作用,使NH4+-N含量不断累积同时不断向NO3--N转换[51].不同泥炭地间无机氮含量以及组分会形成以上差异,说明不同泥炭地间土壤微生物会因为环境因子的不同形成差异,从而导致土壤氮矿化程度不同[52],其次由于不同土层间土壤物理性质(结构、孔隙度)的差异,造成了NO3--N淋溶、NH4+-N吸附程度不同. ...
Variations in microbial community composition through two soil depth profiles
1
2003
... 本研究发现大兴安岭冻土区土壤有效氮以NH4+-N为主,这与肖瑞晗在研究本区域结果相一致[47].由于NO3--N携带被土壤胶体排斥的负电荷[48],一方面在非生长季受降水等影响向较深土壤淋溶,另一方面受到反硝化作用以挥发方式损失,从而导致土壤中NH4+-N明显高于NO3--N.本研究中,XA样地土壤浅层无机氮的含量明显高于深层,这一结果与鲁博权等[49]研究结果相似,即无机氮在垂直方向上具有空间异质性.这是由于冻土区植被根系在汲取氮素时会使有机质在浅层聚集,且浅层土壤因水热条件和土壤基质有效性条件好,氮矿化程度高于微生物的固持[50],而深层土壤则相反,从而导致无机氮含量较低.我们发现在整个秋季冻融时期,土壤NH4+-N在3种冻土泥炭地各土层间不存在显著差异(P>0.05);而NO3--N在浅层和深层土壤中呈现出显著性差异(P<0.05),其中XY受饱和含水量的影响NO3--N平均含量较高,说明较高含水量会促进土壤矿化作用.可能是由于这种条件下微生物活性的增强提高了氨化而抑制了硝化作用,使NH4+-N含量不断累积同时不断向NO3--N转换[51].不同泥炭地间无机氮含量以及组分会形成以上差异,说明不同泥炭地间土壤微生物会因为环境因子的不同形成差异,从而导致土壤氮矿化程度不同[52],其次由于不同土层间土壤物理性质(结构、孔隙度)的差异,造成了NO3--N淋溶、NH4+-N吸附程度不同. ...
Effects of temperature and freeze-thaw on soil nitrogen mineralization in typical Calamagrostis angustifolia wetlands in Sanjiang Plain
1
2011
... 影响冻土区泥炭地无机氮含量的因素多且复杂,其中土壤温度和含水量是影响无机氮转化的主要环境因子[53].在不同生态系统条件下,控制水热条件对无机氮的影响存在差异(表5):森林生态系统中长白山地区温带森林土壤以及湿地生态系统中三江平原沼泽湿地和小兴安岭湿地土壤在经过冻融模拟实验后,均发现培养后无机氮含量高于培养前,这与本研究发现秋季冻融后期土壤无机氮含量明显高于冻融前期的结果相一致,说明土壤氮矿化过程在秋季冻融后期受到强烈影响使硝化底物逐渐聚集,此时环境变化不足以影响硝化细菌活性且硝化过程强于反硝化过程[54],因而导致无机氮含量上升.但是受培养温度不同的影响,研究发现森林系统中大兴安岭落叶松林土壤冻融交替会减少土壤中有效氮的累积. ...
沼泽湿地土壤氮矿化对温度变化及冻融的响应
1
2011
... 影响冻土区泥炭地无机氮含量的因素多且复杂,其中土壤温度和含水量是影响无机氮转化的主要环境因子[53].在不同生态系统条件下,控制水热条件对无机氮的影响存在差异(表5):森林生态系统中长白山地区温带森林土壤以及湿地生态系统中三江平原沼泽湿地和小兴安岭湿地土壤在经过冻融模拟实验后,均发现培养后无机氮含量高于培养前,这与本研究发现秋季冻融后期土壤无机氮含量明显高于冻融前期的结果相一致,说明土壤氮矿化过程在秋季冻融后期受到强烈影响使硝化底物逐渐聚集,此时环境变化不足以影响硝化细菌活性且硝化过程强于反硝化过程[54],因而导致无机氮含量上升.但是受培养温度不同的影响,研究发现森林系统中大兴安岭落叶松林土壤冻融交替会减少土壤中有效氮的累积. ...
Responses of terrestrial nitrogen pools and dynamics to different patterns of freeze-thaw cycle: A meta-analysis
1
2018
... 影响冻土区泥炭地无机氮含量的因素多且复杂,其中土壤温度和含水量是影响无机氮转化的主要环境因子[53].在不同生态系统条件下,控制水热条件对无机氮的影响存在差异(表5):森林生态系统中长白山地区温带森林土壤以及湿地生态系统中三江平原沼泽湿地和小兴安岭湿地土壤在经过冻融模拟实验后,均发现培养后无机氮含量高于培养前,这与本研究发现秋季冻融后期土壤无机氮含量明显高于冻融前期的结果相一致,说明土壤氮矿化过程在秋季冻融后期受到强烈影响使硝化底物逐渐聚集,此时环境变化不足以影响硝化细菌活性且硝化过程强于反硝化过程[54],因而导致无机氮含量上升.但是受培养温度不同的影响,研究发现森林系统中大兴安岭落叶松林土壤冻融交替会减少土壤中有效氮的累积. ...
Characteristics of soil nitrogen mineralization in the main forest types in cold temperate zone during the spring freezing-thawing period
1
2020
... 有研究认为单一的温度[50]或水分[55]条件会对无机氮造成影响,本实验发现在秋季冻融时期土壤温度和含水量与无机氮含量之间存在相关关系,但是与水热的交互作用之间不存在相关性,这与马秀艳[56]在研究该区结果不同,产生差异的原因可能是因为野外原位实验与室内模拟实验条件下,环境因子变化的复杂程度不同,造成了无机氮与水热交互作用的相关性存在差异. ...
春季冻融期寒温带主要森林类型土壤氮矿化特征
1
2020
... 有研究认为单一的温度[50]或水分[55]条件会对无机氮造成影响,本实验发现在秋季冻融时期土壤温度和含水量与无机氮含量之间存在相关关系,但是与水热的交互作用之间不存在相关性,这与马秀艳[56]在研究该区结果不同,产生差异的原因可能是因为野外原位实验与室内模拟实验条件下,环境因子变化的复杂程度不同,造成了无机氮与水热交互作用的相关性存在差异. ...
Effects of temperature and moisture changes on functional gene abundance of soil nitrogen cycle in permafrost peatland
1
2021
... 有研究认为单一的温度[50]或水分[55]条件会对无机氮造成影响,本实验发现在秋季冻融时期土壤温度和含水量与无机氮含量之间存在相关关系,但是与水热的交互作用之间不存在相关性,这与马秀艳[56]在研究该区结果不同,产生差异的原因可能是因为野外原位实验与室内模拟实验条件下,环境因子变化的复杂程度不同,造成了无机氮与水热交互作用的相关性存在差异. ...
温度和水分变化对冻土区泥炭地土壤氮循环功能基因丰度的影响
1
2021
... 有研究认为单一的温度[50]或水分[55]条件会对无机氮造成影响,本实验发现在秋季冻融时期土壤温度和含水量与无机氮含量之间存在相关关系,但是与水热的交互作用之间不存在相关性,这与马秀艳[56]在研究该区结果不同,产生差异的原因可能是因为野外原位实验与室内模拟实验条件下,环境因子变化的复杂程度不同,造成了无机氮与水热交互作用的相关性存在差异. ...
Effect of freezing-thawing on the carbon and nitrogen mineralization in Changbai Mountain
2
2012
... Comparisons of inorganic nitrogen changes under controlled hydrothermal conditions
Table 5| 生态系统 | 研究地点 | 经纬度 | 实验类型 | 冻融处理/次数 | 含水量 | NH4+-N | NO3--N |
|---|
| 森林 | 长白山地区温带森林土壤 | 127° E, 42° N[34] | LT | -10 ℃→10 ℃, 12 h/1、3、5、7、11、15、22、28、36次 | 秋季低含水量(40%) | 不变 | 增加 |
| 127° E, 42° N[57] | LT | -20 ℃→ 15 ℃, 12 h/1、3、5、9、14、21、28、35次 | 最大田间持水量的60% | 增加 | 增加 |
| 河岸带不同植被类型 | 123° E, 41° N[58] | LT | -5 ℃、-20 ℃→ 5℃, 12 h/0、1、5、10、20、30次 | 原土壤含水量 | 增加 | 农田变化不显著 |
| 天山森林土壤 | 87° E, 43° N[31] | ST | 0 ℃以上→ 0 ℃以下, 3 h/3次 | 原土壤含水量 | 草地和灌丛表现为先减少后增加 | 增加 |
| 大兴安岭落叶松林土壤 | 121° E, 50° N[38] | LT | -5 ℃→ 8 ℃, 24 h/1、3、5、7、15次 | 60%土壤饱和含水量 | 低于恒温条件下的累积量 | 低于恒温条件下的累积量 |
| 草地 | 青藏高原高寒草甸区 | 103° E, 32° N[59] | LT | -4 ℃、-20 ℃ → 4℃, 8 h/1、3、5、25、60次 | 原土壤含水量 | 先增后减 | 先增后减 |
| 湿地 | 三江平原沼泽湿地 | 133° E, 47° N[36] | LT | -5 ℃、-25 ℃→ 5 ℃, 1 d/1、2、4、6、10次 | (352.5±1.6) g·kg-1 | 增加 | 增加 |
| 小兴安岭湿地土壤 | 129° E, 48° N[60] | LT | -20 ℃→ 5 ℃, 24 h/5次 | 原土壤含水量 | 先增后减 | 先减后增 |
| 128° E, 48° N[61] | LT | -5 ℃、-25 ℃→ 5 ℃, 24 h/0、1、2、4、9次 | 原土壤含水量 | 先增后减 | 先减后增 |
注:LT:室内模拟试验;ST:野外原位试验. ...
... 经逐步回归分析表明(表2~4),在不同冻融阶段,无机氮对环境因子的响应程度也存在差异.在整个秋季冻融期,BM浅层(10~20 cm)含水量对无机氮含量的影响程度最大(R2=0.301),这是由于浅层土壤含水量更容易影响土壤通气性和土壤微生物活性,进而改变影响了土壤基质的养分循环,从而对无机氮转化产生影响.通常认为无机氮对温度的响应程度大于含水量[62],但是BM样地在整个秋季冻融时期,温度和含水量都处于较低水平且变化幅度较小,与温度相比含水量对土壤微生物活性起有效作用,这与赵媛[34]在研究秋末冻融循环对长白山地区无机氮的影响结果相似,即无机氮对含水量的响应程度大于温度.在秋季冻融前、中和后期,3种多年冻土泥炭地浅层(0~20 cm)土壤的环境因子与无机氮释放有关,其中浅层土壤温度对无机氮释放的贡献率最大(R2=0.685),这主要受当地非生长季寒冷气候的影响,温度通过改变土壤中生物酶和氨氧化细菌、硝化细菌的活性来影响氮的分解和无机氮的释放[63].但是不同土壤环境条件下微生物对温度敏感程度有很大差异[64],本研究发现,受非生长季温度逐渐降低的影响,XA仅在秋季冻融前期土壤温度对无机氮释放有贡献(R2=0.685),在秋季冻融后期不存在相关关系,这与表5中草地生态系统下青藏高原高寒草甸区研究发现短期的冻融交替可以显著增加土壤中的氮矿化累积结果相似.说明与长期冻融相比,短期温度变化会破坏土壤团聚结构从而影响氨氧化细菌的活性,一部分微生物为了维持繁殖不断汲取死亡微生物的养分,从而促进了微生物的活性,影响土壤氮矿化速度.但也有研究发现,短期冻融对土壤氮矿化并没有影响[57],土壤微生物会对温度变化做出调节反应,从而无机氮变化不显著.秋季冻融期无机氮对环境因子的响应程度存在以上差异的原因可能有以下四方面:第一,样地沿一定坡度选取,地势高低不同造成了土壤含水量存在显著差异.第二,受到地表植被的影响,土壤微生物的类别和数量存在差异因此对氮矿化程度不同[44].第三,与凋落物有关,不同凋落物分解的有机质不同会影响土壤有效基质[65].第四,3种泥炭地土壤的容重、孔隙度和pH值等物理性质存在差异[66].这些要素的综合影响造成了不同泥炭地间,以及冻融期不同时期间无机氮含量对温度和含水量响应程度的不同. ...
冻融对长白山森林土壤碳氮矿化的影响
2
2012
... Comparisons of inorganic nitrogen changes under controlled hydrothermal conditions
Table 5| 生态系统 | 研究地点 | 经纬度 | 实验类型 | 冻融处理/次数 | 含水量 | NH4+-N | NO3--N |
|---|
| 森林 | 长白山地区温带森林土壤 | 127° E, 42° N[34] | LT | -10 ℃→10 ℃, 12 h/1、3、5、7、11、15、22、28、36次 | 秋季低含水量(40%) | 不变 | 增加 |
| 127° E, 42° N[57] | LT | -20 ℃→ 15 ℃, 12 h/1、3、5、9、14、21、28、35次 | 最大田间持水量的60% | 增加 | 增加 |
| 河岸带不同植被类型 | 123° E, 41° N[58] | LT | -5 ℃、-20 ℃→ 5℃, 12 h/0、1、5、10、20、30次 | 原土壤含水量 | 增加 | 农田变化不显著 |
| 天山森林土壤 | 87° E, 43° N[31] | ST | 0 ℃以上→ 0 ℃以下, 3 h/3次 | 原土壤含水量 | 草地和灌丛表现为先减少后增加 | 增加 |
| 大兴安岭落叶松林土壤 | 121° E, 50° N[38] | LT | -5 ℃→ 8 ℃, 24 h/1、3、5、7、15次 | 60%土壤饱和含水量 | 低于恒温条件下的累积量 | 低于恒温条件下的累积量 |
| 草地 | 青藏高原高寒草甸区 | 103° E, 32° N[59] | LT | -4 ℃、-20 ℃ → 4℃, 8 h/1、3、5、25、60次 | 原土壤含水量 | 先增后减 | 先增后减 |
| 湿地 | 三江平原沼泽湿地 | 133° E, 47° N[36] | LT | -5 ℃、-25 ℃→ 5 ℃, 1 d/1、2、4、6、10次 | (352.5±1.6) g·kg-1 | 增加 | 增加 |
| 小兴安岭湿地土壤 | 129° E, 48° N[60] | LT | -20 ℃→ 5 ℃, 24 h/5次 | 原土壤含水量 | 先增后减 | 先减后增 |
| 128° E, 48° N[61] | LT | -5 ℃、-25 ℃→ 5 ℃, 24 h/0、1、2、4、9次 | 原土壤含水量 | 先增后减 | 先减后增 |
注:LT:室内模拟试验;ST:野外原位试验. ...
... 经逐步回归分析表明(表2~4),在不同冻融阶段,无机氮对环境因子的响应程度也存在差异.在整个秋季冻融期,BM浅层(10~20 cm)含水量对无机氮含量的影响程度最大(R2=0.301),这是由于浅层土壤含水量更容易影响土壤通气性和土壤微生物活性,进而改变影响了土壤基质的养分循环,从而对无机氮转化产生影响.通常认为无机氮对温度的响应程度大于含水量[62],但是BM样地在整个秋季冻融时期,温度和含水量都处于较低水平且变化幅度较小,与温度相比含水量对土壤微生物活性起有效作用,这与赵媛[34]在研究秋末冻融循环对长白山地区无机氮的影响结果相似,即无机氮对含水量的响应程度大于温度.在秋季冻融前、中和后期,3种多年冻土泥炭地浅层(0~20 cm)土壤的环境因子与无机氮释放有关,其中浅层土壤温度对无机氮释放的贡献率最大(R2=0.685),这主要受当地非生长季寒冷气候的影响,温度通过改变土壤中生物酶和氨氧化细菌、硝化细菌的活性来影响氮的分解和无机氮的释放[63].但是不同土壤环境条件下微生物对温度敏感程度有很大差异[64],本研究发现,受非生长季温度逐渐降低的影响,XA仅在秋季冻融前期土壤温度对无机氮释放有贡献(R2=0.685),在秋季冻融后期不存在相关关系,这与表5中草地生态系统下青藏高原高寒草甸区研究发现短期的冻融交替可以显著增加土壤中的氮矿化累积结果相似.说明与长期冻融相比,短期温度变化会破坏土壤团聚结构从而影响氨氧化细菌的活性,一部分微生物为了维持繁殖不断汲取死亡微生物的养分,从而促进了微生物的活性,影响土壤氮矿化速度.但也有研究发现,短期冻融对土壤氮矿化并没有影响[57],土壤微生物会对温度变化做出调节反应,从而无机氮变化不显著.秋季冻融期无机氮对环境因子的响应程度存在以上差异的原因可能有以下四方面:第一,样地沿一定坡度选取,地势高低不同造成了土壤含水量存在显著差异.第二,受到地表植被的影响,土壤微生物的类别和数量存在差异因此对氮矿化程度不同[44].第三,与凋落物有关,不同凋落物分解的有机质不同会影响土壤有效基质[65].第四,3种泥炭地土壤的容重、孔隙度和pH值等物理性质存在差异[66].这些要素的综合影响造成了不同泥炭地间,以及冻融期不同时期间无机氮含量对温度和含水量响应程度的不同. ...
Responses of soil nitrogen transformation process to freezing-thawing cycle in a riparian zone under three different vegetation types
1
2013
... Comparisons of inorganic nitrogen changes under controlled hydrothermal conditions
Table 5| 生态系统 | 研究地点 | 经纬度 | 实验类型 | 冻融处理/次数 | 含水量 | NH4+-N | NO3--N |
|---|
| 森林 | 长白山地区温带森林土壤 | 127° E, 42° N[34] | LT | -10 ℃→10 ℃, 12 h/1、3、5、7、11、15、22、28、36次 | 秋季低含水量(40%) | 不变 | 增加 |
| 127° E, 42° N[57] | LT | -20 ℃→ 15 ℃, 12 h/1、3、5、9、14、21、28、35次 | 最大田间持水量的60% | 增加 | 增加 |
| 河岸带不同植被类型 | 123° E, 41° N[58] | LT | -5 ℃、-20 ℃→ 5℃, 12 h/0、1、5、10、20、30次 | 原土壤含水量 | 增加 | 农田变化不显著 |
| 天山森林土壤 | 87° E, 43° N[31] | ST | 0 ℃以上→ 0 ℃以下, 3 h/3次 | 原土壤含水量 | 草地和灌丛表现为先减少后增加 | 增加 |
| 大兴安岭落叶松林土壤 | 121° E, 50° N[38] | LT | -5 ℃→ 8 ℃, 24 h/1、3、5、7、15次 | 60%土壤饱和含水量 | 低于恒温条件下的累积量 | 低于恒温条件下的累积量 |
| 草地 | 青藏高原高寒草甸区 | 103° E, 32° N[59] | LT | -4 ℃、-20 ℃ → 4℃, 8 h/1、3、5、25、60次 | 原土壤含水量 | 先增后减 | 先增后减 |
| 湿地 | 三江平原沼泽湿地 | 133° E, 47° N[36] | LT | -5 ℃、-25 ℃→ 5 ℃, 1 d/1、2、4、6、10次 | (352.5±1.6) g·kg-1 | 增加 | 增加 |
| 小兴安岭湿地土壤 | 129° E, 48° N[60] | LT | -20 ℃→ 5 ℃, 24 h/5次 | 原土壤含水量 | 先增后减 | 先减后增 |
| 128° E, 48° N[61] | LT | -5 ℃、-25 ℃→ 5 ℃, 24 h/0、1、2、4、9次 | 原土壤含水量 | 先增后减 | 先减后增 |
注:LT:室内模拟试验;ST:野外原位试验. ...
河岸带不同植被类型土壤氮素转化过程对冻融交替的响应
1
2013
... Comparisons of inorganic nitrogen changes under controlled hydrothermal conditions
Table 5| 生态系统 | 研究地点 | 经纬度 | 实验类型 | 冻融处理/次数 | 含水量 | NH4+-N | NO3--N |
|---|
| 森林 | 长白山地区温带森林土壤 | 127° E, 42° N[34] | LT | -10 ℃→10 ℃, 12 h/1、3、5、7、11、15、22、28、36次 | 秋季低含水量(40%) | 不变 | 增加 |
| 127° E, 42° N[57] | LT | -20 ℃→ 15 ℃, 12 h/1、3、5、9、14、21、28、35次 | 最大田间持水量的60% | 增加 | 增加 |
| 河岸带不同植被类型 | 123° E, 41° N[58] | LT | -5 ℃、-20 ℃→ 5℃, 12 h/0、1、5、10、20、30次 | 原土壤含水量 | 增加 | 农田变化不显著 |
| 天山森林土壤 | 87° E, 43° N[31] | ST | 0 ℃以上→ 0 ℃以下, 3 h/3次 | 原土壤含水量 | 草地和灌丛表现为先减少后增加 | 增加 |
| 大兴安岭落叶松林土壤 | 121° E, 50° N[38] | LT | -5 ℃→ 8 ℃, 24 h/1、3、5、7、15次 | 60%土壤饱和含水量 | 低于恒温条件下的累积量 | 低于恒温条件下的累积量 |
| 草地 | 青藏高原高寒草甸区 | 103° E, 32° N[59] | LT | -4 ℃、-20 ℃ → 4℃, 8 h/1、3、5、25、60次 | 原土壤含水量 | 先增后减 | 先增后减 |
| 湿地 | 三江平原沼泽湿地 | 133° E, 47° N[36] | LT | -5 ℃、-25 ℃→ 5 ℃, 1 d/1、2、4、6、10次 | (352.5±1.6) g·kg-1 | 增加 | 增加 |
| 小兴安岭湿地土壤 | 129° E, 48° N[60] | LT | -20 ℃→ 5 ℃, 24 h/5次 | 原土壤含水量 | 先增后减 | 先减后增 |
| 128° E, 48° N[61] | LT | -5 ℃、-25 ℃→ 5 ℃, 24 h/0、1、2、4、9次 | 原土壤含水量 | 先增后减 | 先减后增 |
注:LT:室内模拟试验;ST:野外原位试验. ...
Effects of freezing and thawing alternation on soil nitrogen pool in the alpine meadow
1
2010
... Comparisons of inorganic nitrogen changes under controlled hydrothermal conditions
Table 5| 生态系统 | 研究地点 | 经纬度 | 实验类型 | 冻融处理/次数 | 含水量 | NH4+-N | NO3--N |
|---|
| 森林 | 长白山地区温带森林土壤 | 127° E, 42° N[34] | LT | -10 ℃→10 ℃, 12 h/1、3、5、7、11、15、22、28、36次 | 秋季低含水量(40%) | 不变 | 增加 |
| 127° E, 42° N[57] | LT | -20 ℃→ 15 ℃, 12 h/1、3、5、9、14、21、28、35次 | 最大田间持水量的60% | 增加 | 增加 |
| 河岸带不同植被类型 | 123° E, 41° N[58] | LT | -5 ℃、-20 ℃→ 5℃, 12 h/0、1、5、10、20、30次 | 原土壤含水量 | 增加 | 农田变化不显著 |
| 天山森林土壤 | 87° E, 43° N[31] | ST | 0 ℃以上→ 0 ℃以下, 3 h/3次 | 原土壤含水量 | 草地和灌丛表现为先减少后增加 | 增加 |
| 大兴安岭落叶松林土壤 | 121° E, 50° N[38] | LT | -5 ℃→ 8 ℃, 24 h/1、3、5、7、15次 | 60%土壤饱和含水量 | 低于恒温条件下的累积量 | 低于恒温条件下的累积量 |
| 草地 | 青藏高原高寒草甸区 | 103° E, 32° N[59] | LT | -4 ℃、-20 ℃ → 4℃, 8 h/1、3、5、25、60次 | 原土壤含水量 | 先增后减 | 先增后减 |
| 湿地 | 三江平原沼泽湿地 | 133° E, 47° N[36] | LT | -5 ℃、-25 ℃→ 5 ℃, 1 d/1、2、4、6、10次 | (352.5±1.6) g·kg-1 | 增加 | 增加 |
| 小兴安岭湿地土壤 | 129° E, 48° N[60] | LT | -20 ℃→ 5 ℃, 24 h/5次 | 原土壤含水量 | 先增后减 | 先减后增 |
| 128° E, 48° N[61] | LT | -5 ℃、-25 ℃→ 5 ℃, 24 h/0、1、2、4、9次 | 原土壤含水量 | 先增后减 | 先减后增 |
注:LT:室内模拟试验;ST:野外原位试验. ...
冻融交替对高寒草甸土壤氮素的影响
1
2010
... Comparisons of inorganic nitrogen changes under controlled hydrothermal conditions
Table 5| 生态系统 | 研究地点 | 经纬度 | 实验类型 | 冻融处理/次数 | 含水量 | NH4+-N | NO3--N |
|---|
| 森林 | 长白山地区温带森林土壤 | 127° E, 42° N[34] | LT | -10 ℃→10 ℃, 12 h/1、3、5、7、11、15、22、28、36次 | 秋季低含水量(40%) | 不变 | 增加 |
| 127° E, 42° N[57] | LT | -20 ℃→ 15 ℃, 12 h/1、3、5、9、14、21、28、35次 | 最大田间持水量的60% | 增加 | 增加 |
| 河岸带不同植被类型 | 123° E, 41° N[58] | LT | -5 ℃、-20 ℃→ 5℃, 12 h/0、1、5、10、20、30次 | 原土壤含水量 | 增加 | 农田变化不显著 |
| 天山森林土壤 | 87° E, 43° N[31] | ST | 0 ℃以上→ 0 ℃以下, 3 h/3次 | 原土壤含水量 | 草地和灌丛表现为先减少后增加 | 增加 |
| 大兴安岭落叶松林土壤 | 121° E, 50° N[38] | LT | -5 ℃→ 8 ℃, 24 h/1、3、5、7、15次 | 60%土壤饱和含水量 | 低于恒温条件下的累积量 | 低于恒温条件下的累积量 |
| 草地 | 青藏高原高寒草甸区 | 103° E, 32° N[59] | LT | -4 ℃、-20 ℃ → 4℃, 8 h/1、3、5、25、60次 | 原土壤含水量 | 先增后减 | 先增后减 |
| 湿地 | 三江平原沼泽湿地 | 133° E, 47° N[36] | LT | -5 ℃、-25 ℃→ 5 ℃, 1 d/1、2、4、6、10次 | (352.5±1.6) g·kg-1 | 增加 | 增加 |
| 小兴安岭湿地土壤 | 129° E, 48° N[60] | LT | -20 ℃→ 5 ℃, 24 h/5次 | 原土壤含水量 | 先增后减 | 先减后增 |
| 128° E, 48° N[61] | LT | -5 ℃、-25 ℃→ 5 ℃, 24 h/0、1、2、4、9次 | 原土壤含水量 | 先增后减 | 先减后增 |
注:LT:室内模拟试验;ST:野外原位试验. ...
Effects of freeze-thaw cycles on soil microbial carbon, nitrogen and nitrogen conversion in Xiaoxing’anling boreal wetland of China
1
2013
... Comparisons of inorganic nitrogen changes under controlled hydrothermal conditions
Table 5| 生态系统 | 研究地点 | 经纬度 | 实验类型 | 冻融处理/次数 | 含水量 | NH4+-N | NO3--N |
|---|
| 森林 | 长白山地区温带森林土壤 | 127° E, 42° N[34] | LT | -10 ℃→10 ℃, 12 h/1、3、5、7、11、15、22、28、36次 | 秋季低含水量(40%) | 不变 | 增加 |
| 127° E, 42° N[57] | LT | -20 ℃→ 15 ℃, 12 h/1、3、5、9、14、21、28、35次 | 最大田间持水量的60% | 增加 | 增加 |
| 河岸带不同植被类型 | 123° E, 41° N[58] | LT | -5 ℃、-20 ℃→ 5℃, 12 h/0、1、5、10、20、30次 | 原土壤含水量 | 增加 | 农田变化不显著 |
| 天山森林土壤 | 87° E, 43° N[31] | ST | 0 ℃以上→ 0 ℃以下, 3 h/3次 | 原土壤含水量 | 草地和灌丛表现为先减少后增加 | 增加 |
| 大兴安岭落叶松林土壤 | 121° E, 50° N[38] | LT | -5 ℃→ 8 ℃, 24 h/1、3、5、7、15次 | 60%土壤饱和含水量 | 低于恒温条件下的累积量 | 低于恒温条件下的累积量 |
| 草地 | 青藏高原高寒草甸区 | 103° E, 32° N[59] | LT | -4 ℃、-20 ℃ → 4℃, 8 h/1、3、5、25、60次 | 原土壤含水量 | 先增后减 | 先增后减 |
| 湿地 | 三江平原沼泽湿地 | 133° E, 47° N[36] | LT | -5 ℃、-25 ℃→ 5 ℃, 1 d/1、2、4、6、10次 | (352.5±1.6) g·kg-1 | 增加 | 增加 |
| 小兴安岭湿地土壤 | 129° E, 48° N[60] | LT | -20 ℃→ 5 ℃, 24 h/5次 | 原土壤含水量 | 先增后减 | 先减后增 |
| 128° E, 48° N[61] | LT | -5 ℃、-25 ℃→ 5 ℃, 24 h/0、1、2、4、9次 | 原土壤含水量 | 先增后减 | 先减后增 |
注:LT:室内模拟试验;ST:野外原位试验. ...
冻融对小兴安岭湿地土壤微生物碳、氮和氮转换的影响
1
2013
... Comparisons of inorganic nitrogen changes under controlled hydrothermal conditions
Table 5| 生态系统 | 研究地点 | 经纬度 | 实验类型 | 冻融处理/次数 | 含水量 | NH4+-N | NO3--N |
|---|
| 森林 | 长白山地区温带森林土壤 | 127° E, 42° N[34] | LT | -10 ℃→10 ℃, 12 h/1、3、5、7、11、15、22、28、36次 | 秋季低含水量(40%) | 不变 | 增加 |
| 127° E, 42° N[57] | LT | -20 ℃→ 15 ℃, 12 h/1、3、5、9、14、21、28、35次 | 最大田间持水量的60% | 增加 | 增加 |
| 河岸带不同植被类型 | 123° E, 41° N[58] | LT | -5 ℃、-20 ℃→ 5℃, 12 h/0、1、5、10、20、30次 | 原土壤含水量 | 增加 | 农田变化不显著 |
| 天山森林土壤 | 87° E, 43° N[31] | ST | 0 ℃以上→ 0 ℃以下, 3 h/3次 | 原土壤含水量 | 草地和灌丛表现为先减少后增加 | 增加 |
| 大兴安岭落叶松林土壤 | 121° E, 50° N[38] | LT | -5 ℃→ 8 ℃, 24 h/1、3、5、7、15次 | 60%土壤饱和含水量 | 低于恒温条件下的累积量 | 低于恒温条件下的累积量 |
| 草地 | 青藏高原高寒草甸区 | 103° E, 32° N[59] | LT | -4 ℃、-20 ℃ → 4℃, 8 h/1、3、5、25、60次 | 原土壤含水量 | 先增后减 | 先增后减 |
| 湿地 | 三江平原沼泽湿地 | 133° E, 47° N[36] | LT | -5 ℃、-25 ℃→ 5 ℃, 1 d/1、2、4、6、10次 | (352.5±1.6) g·kg-1 | 增加 | 增加 |
| 小兴安岭湿地土壤 | 129° E, 48° N[60] | LT | -20 ℃→ 5 ℃, 24 h/5次 | 原土壤含水量 | 先增后减 | 先减后增 |
| 128° E, 48° N[61] | LT | -5 ℃、-25 ℃→ 5 ℃, 24 h/0、1、2、4、9次 | 原土壤含水量 | 先增后减 | 先减后增 |
注:LT:室内模拟试验;ST:野外原位试验. ...
Effect of freeze-thaw action on dissolved organic carbon and nitrogen mineralization of wetland soil in Xiaoxing’an Mountains
1
2015
... Comparisons of inorganic nitrogen changes under controlled hydrothermal conditions
Table 5| 生态系统 | 研究地点 | 经纬度 | 实验类型 | 冻融处理/次数 | 含水量 | NH4+-N | NO3--N |
|---|
| 森林 | 长白山地区温带森林土壤 | 127° E, 42° N[34] | LT | -10 ℃→10 ℃, 12 h/1、3、5、7、11、15、22、28、36次 | 秋季低含水量(40%) | 不变 | 增加 |
| 127° E, 42° N[57] | LT | -20 ℃→ 15 ℃, 12 h/1、3、5、9、14、21、28、35次 | 最大田间持水量的60% | 增加 | 增加 |
| 河岸带不同植被类型 | 123° E, 41° N[58] | LT | -5 ℃、-20 ℃→ 5℃, 12 h/0、1、5、10、20、30次 | 原土壤含水量 | 增加 | 农田变化不显著 |
| 天山森林土壤 | 87° E, 43° N[31] | ST | 0 ℃以上→ 0 ℃以下, 3 h/3次 | 原土壤含水量 | 草地和灌丛表现为先减少后增加 | 增加 |
| 大兴安岭落叶松林土壤 | 121° E, 50° N[38] | LT | -5 ℃→ 8 ℃, 24 h/1、3、5、7、15次 | 60%土壤饱和含水量 | 低于恒温条件下的累积量 | 低于恒温条件下的累积量 |
| 草地 | 青藏高原高寒草甸区 | 103° E, 32° N[59] | LT | -4 ℃、-20 ℃ → 4℃, 8 h/1、3、5、25、60次 | 原土壤含水量 | 先增后减 | 先增后减 |
| 湿地 | 三江平原沼泽湿地 | 133° E, 47° N[36] | LT | -5 ℃、-25 ℃→ 5 ℃, 1 d/1、2、4、6、10次 | (352.5±1.6) g·kg-1 | 增加 | 增加 |
| 小兴安岭湿地土壤 | 129° E, 48° N[60] | LT | -20 ℃→ 5 ℃, 24 h/5次 | 原土壤含水量 | 先增后减 | 先减后增 |
| 128° E, 48° N[61] | LT | -5 ℃、-25 ℃→ 5 ℃, 24 h/0、1、2、4、9次 | 原土壤含水量 | 先增后减 | 先减后增 |
注:LT:室内模拟试验;ST:野外原位试验. ...
冻融作用对小兴安岭湿地土壤溶解性有机碳和氮素矿化的影响
1
2015
... Comparisons of inorganic nitrogen changes under controlled hydrothermal conditions
Table 5| 生态系统 | 研究地点 | 经纬度 | 实验类型 | 冻融处理/次数 | 含水量 | NH4+-N | NO3--N |
|---|
| 森林 | 长白山地区温带森林土壤 | 127° E, 42° N[34] | LT | -10 ℃→10 ℃, 12 h/1、3、5、7、11、15、22、28、36次 | 秋季低含水量(40%) | 不变 | 增加 |
| 127° E, 42° N[57] | LT | -20 ℃→ 15 ℃, 12 h/1、3、5、9、14、21、28、35次 | 最大田间持水量的60% | 增加 | 增加 |
| 河岸带不同植被类型 | 123° E, 41° N[58] | LT | -5 ℃、-20 ℃→ 5℃, 12 h/0、1、5、10、20、30次 | 原土壤含水量 | 增加 | 农田变化不显著 |
| 天山森林土壤 | 87° E, 43° N[31] | ST | 0 ℃以上→ 0 ℃以下, 3 h/3次 | 原土壤含水量 | 草地和灌丛表现为先减少后增加 | 增加 |
| 大兴安岭落叶松林土壤 | 121° E, 50° N[38] | LT | -5 ℃→ 8 ℃, 24 h/1、3、5、7、15次 | 60%土壤饱和含水量 | 低于恒温条件下的累积量 | 低于恒温条件下的累积量 |
| 草地 | 青藏高原高寒草甸区 | 103° E, 32° N[59] | LT | -4 ℃、-20 ℃ → 4℃, 8 h/1、3、5、25、60次 | 原土壤含水量 | 先增后减 | 先增后减 |
| 湿地 | 三江平原沼泽湿地 | 133° E, 47° N[36] | LT | -5 ℃、-25 ℃→ 5 ℃, 1 d/1、2、4、6、10次 | (352.5±1.6) g·kg-1 | 增加 | 增加 |
| 小兴安岭湿地土壤 | 129° E, 48° N[60] | LT | -20 ℃→ 5 ℃, 24 h/5次 | 原土壤含水量 | 先增后减 | 先减后增 |
| 128° E, 48° N[61] | LT | -5 ℃、-25 ℃→ 5 ℃, 24 h/0、1、2、4、9次 | 原土壤含水量 | 先增后减 | 先减后增 |
注:LT:室内模拟试验;ST:野外原位试验. ...
Role of soil water content in the carbon and nitrogen dynamics of Lumbricus terrestris L. burrow soil
1
2005
... 经逐步回归分析表明(表2~4),在不同冻融阶段,无机氮对环境因子的响应程度也存在差异.在整个秋季冻融期,BM浅层(10~20 cm)含水量对无机氮含量的影响程度最大(R2=0.301),这是由于浅层土壤含水量更容易影响土壤通气性和土壤微生物活性,进而改变影响了土壤基质的养分循环,从而对无机氮转化产生影响.通常认为无机氮对温度的响应程度大于含水量[62],但是BM样地在整个秋季冻融时期,温度和含水量都处于较低水平且变化幅度较小,与温度相比含水量对土壤微生物活性起有效作用,这与赵媛[34]在研究秋末冻融循环对长白山地区无机氮的影响结果相似,即无机氮对含水量的响应程度大于温度.在秋季冻融前、中和后期,3种多年冻土泥炭地浅层(0~20 cm)土壤的环境因子与无机氮释放有关,其中浅层土壤温度对无机氮释放的贡献率最大(R2=0.685),这主要受当地非生长季寒冷气候的影响,温度通过改变土壤中生物酶和氨氧化细菌、硝化细菌的活性来影响氮的分解和无机氮的释放[63].但是不同土壤环境条件下微生物对温度敏感程度有很大差异[64],本研究发现,受非生长季温度逐渐降低的影响,XA仅在秋季冻融前期土壤温度对无机氮释放有贡献(R2=0.685),在秋季冻融后期不存在相关关系,这与表5中草地生态系统下青藏高原高寒草甸区研究发现短期的冻融交替可以显著增加土壤中的氮矿化累积结果相似.说明与长期冻融相比,短期温度变化会破坏土壤团聚结构从而影响氨氧化细菌的活性,一部分微生物为了维持繁殖不断汲取死亡微生物的养分,从而促进了微生物的活性,影响土壤氮矿化速度.但也有研究发现,短期冻融对土壤氮矿化并没有影响[57],土壤微生物会对温度变化做出调节反应,从而无机氮变化不显著.秋季冻融期无机氮对环境因子的响应程度存在以上差异的原因可能有以下四方面:第一,样地沿一定坡度选取,地势高低不同造成了土壤含水量存在显著差异.第二,受到地表植被的影响,土壤微生物的类别和数量存在差异因此对氮矿化程度不同[44].第三,与凋落物有关,不同凋落物分解的有机质不同会影响土壤有效基质[65].第四,3种泥炭地土壤的容重、孔隙度和pH值等物理性质存在差异[66].这些要素的综合影响造成了不同泥炭地间,以及冻融期不同时期间无机氮含量对温度和含水量响应程度的不同. ...
On the variability of respiration in terrestrial ecosystems: moving beyond Q10
1
2006
... 经逐步回归分析表明(表2~4),在不同冻融阶段,无机氮对环境因子的响应程度也存在差异.在整个秋季冻融期,BM浅层(10~20 cm)含水量对无机氮含量的影响程度最大(R2=0.301),这是由于浅层土壤含水量更容易影响土壤通气性和土壤微生物活性,进而改变影响了土壤基质的养分循环,从而对无机氮转化产生影响.通常认为无机氮对温度的响应程度大于含水量[62],但是BM样地在整个秋季冻融时期,温度和含水量都处于较低水平且变化幅度较小,与温度相比含水量对土壤微生物活性起有效作用,这与赵媛[34]在研究秋末冻融循环对长白山地区无机氮的影响结果相似,即无机氮对含水量的响应程度大于温度.在秋季冻融前、中和后期,3种多年冻土泥炭地浅层(0~20 cm)土壤的环境因子与无机氮释放有关,其中浅层土壤温度对无机氮释放的贡献率最大(R2=0.685),这主要受当地非生长季寒冷气候的影响,温度通过改变土壤中生物酶和氨氧化细菌、硝化细菌的活性来影响氮的分解和无机氮的释放[63].但是不同土壤环境条件下微生物对温度敏感程度有很大差异[64],本研究发现,受非生长季温度逐渐降低的影响,XA仅在秋季冻融前期土壤温度对无机氮释放有贡献(R2=0.685),在秋季冻融后期不存在相关关系,这与表5中草地生态系统下青藏高原高寒草甸区研究发现短期的冻融交替可以显著增加土壤中的氮矿化累积结果相似.说明与长期冻融相比,短期温度变化会破坏土壤团聚结构从而影响氨氧化细菌的活性,一部分微生物为了维持繁殖不断汲取死亡微生物的养分,从而促进了微生物的活性,影响土壤氮矿化速度.但也有研究发现,短期冻融对土壤氮矿化并没有影响[57],土壤微生物会对温度变化做出调节反应,从而无机氮变化不显著.秋季冻融期无机氮对环境因子的响应程度存在以上差异的原因可能有以下四方面:第一,样地沿一定坡度选取,地势高低不同造成了土壤含水量存在显著差异.第二,受到地表植被的影响,土壤微生物的类别和数量存在差异因此对氮矿化程度不同[44].第三,与凋落物有关,不同凋落物分解的有机质不同会影响土壤有效基质[65].第四,3种泥炭地土壤的容重、孔隙度和pH值等物理性质存在差异[66].这些要素的综合影响造成了不同泥炭地间,以及冻融期不同时期间无机氮含量对温度和含水量响应程度的不同. ...
Soil enzyme activity and microbial biomass of Haloxylon ammodendron community under freeze-thaw action
1
2015
... 经逐步回归分析表明(表2~4),在不同冻融阶段,无机氮对环境因子的响应程度也存在差异.在整个秋季冻融期,BM浅层(10~20 cm)含水量对无机氮含量的影响程度最大(R2=0.301),这是由于浅层土壤含水量更容易影响土壤通气性和土壤微生物活性,进而改变影响了土壤基质的养分循环,从而对无机氮转化产生影响.通常认为无机氮对温度的响应程度大于含水量[62],但是BM样地在整个秋季冻融时期,温度和含水量都处于较低水平且变化幅度较小,与温度相比含水量对土壤微生物活性起有效作用,这与赵媛[34]在研究秋末冻融循环对长白山地区无机氮的影响结果相似,即无机氮对含水量的响应程度大于温度.在秋季冻融前、中和后期,3种多年冻土泥炭地浅层(0~20 cm)土壤的环境因子与无机氮释放有关,其中浅层土壤温度对无机氮释放的贡献率最大(R2=0.685),这主要受当地非生长季寒冷气候的影响,温度通过改变土壤中生物酶和氨氧化细菌、硝化细菌的活性来影响氮的分解和无机氮的释放[63].但是不同土壤环境条件下微生物对温度敏感程度有很大差异[64],本研究发现,受非生长季温度逐渐降低的影响,XA仅在秋季冻融前期土壤温度对无机氮释放有贡献(R2=0.685),在秋季冻融后期不存在相关关系,这与表5中草地生态系统下青藏高原高寒草甸区研究发现短期的冻融交替可以显著增加土壤中的氮矿化累积结果相似.说明与长期冻融相比,短期温度变化会破坏土壤团聚结构从而影响氨氧化细菌的活性,一部分微生物为了维持繁殖不断汲取死亡微生物的养分,从而促进了微生物的活性,影响土壤氮矿化速度.但也有研究发现,短期冻融对土壤氮矿化并没有影响[57],土壤微生物会对温度变化做出调节反应,从而无机氮变化不显著.秋季冻融期无机氮对环境因子的响应程度存在以上差异的原因可能有以下四方面:第一,样地沿一定坡度选取,地势高低不同造成了土壤含水量存在显著差异.第二,受到地表植被的影响,土壤微生物的类别和数量存在差异因此对氮矿化程度不同[44].第三,与凋落物有关,不同凋落物分解的有机质不同会影响土壤有效基质[65].第四,3种泥炭地土壤的容重、孔隙度和pH值等物理性质存在差异[66].这些要素的综合影响造成了不同泥炭地间,以及冻融期不同时期间无机氮含量对温度和含水量响应程度的不同. ...
艾比湖地区冻融作用对梭梭群落土壤酶活性及微生物数量的影响
1
2015
... 经逐步回归分析表明(表2~4),在不同冻融阶段,无机氮对环境因子的响应程度也存在差异.在整个秋季冻融期,BM浅层(10~20 cm)含水量对无机氮含量的影响程度最大(R2=0.301),这是由于浅层土壤含水量更容易影响土壤通气性和土壤微生物活性,进而改变影响了土壤基质的养分循环,从而对无机氮转化产生影响.通常认为无机氮对温度的响应程度大于含水量[62],但是BM样地在整个秋季冻融时期,温度和含水量都处于较低水平且变化幅度较小,与温度相比含水量对土壤微生物活性起有效作用,这与赵媛[34]在研究秋末冻融循环对长白山地区无机氮的影响结果相似,即无机氮对含水量的响应程度大于温度.在秋季冻融前、中和后期,3种多年冻土泥炭地浅层(0~20 cm)土壤的环境因子与无机氮释放有关,其中浅层土壤温度对无机氮释放的贡献率最大(R2=0.685),这主要受当地非生长季寒冷气候的影响,温度通过改变土壤中生物酶和氨氧化细菌、硝化细菌的活性来影响氮的分解和无机氮的释放[63].但是不同土壤环境条件下微生物对温度敏感程度有很大差异[64],本研究发现,受非生长季温度逐渐降低的影响,XA仅在秋季冻融前期土壤温度对无机氮释放有贡献(R2=0.685),在秋季冻融后期不存在相关关系,这与表5中草地生态系统下青藏高原高寒草甸区研究发现短期的冻融交替可以显著增加土壤中的氮矿化累积结果相似.说明与长期冻融相比,短期温度变化会破坏土壤团聚结构从而影响氨氧化细菌的活性,一部分微生物为了维持繁殖不断汲取死亡微生物的养分,从而促进了微生物的活性,影响土壤氮矿化速度.但也有研究发现,短期冻融对土壤氮矿化并没有影响[57],土壤微生物会对温度变化做出调节反应,从而无机氮变化不显著.秋季冻融期无机氮对环境因子的响应程度存在以上差异的原因可能有以下四方面:第一,样地沿一定坡度选取,地势高低不同造成了土壤含水量存在显著差异.第二,受到地表植被的影响,土壤微生物的类别和数量存在差异因此对氮矿化程度不同[44].第三,与凋落物有关,不同凋落物分解的有机质不同会影响土壤有效基质[65].第四,3种泥炭地土壤的容重、孔隙度和pH值等物理性质存在差异[66].这些要素的综合影响造成了不同泥炭地间,以及冻融期不同时期间无机氮含量对温度和含水量响应程度的不同. ...
A review on the development of study on hydrothermal characteristics of active layer in permafrost areas in Qinghai-Tibet Plateau
1
2020
... 经逐步回归分析表明(表2~4),在不同冻融阶段,无机氮对环境因子的响应程度也存在差异.在整个秋季冻融期,BM浅层(10~20 cm)含水量对无机氮含量的影响程度最大(R2=0.301),这是由于浅层土壤含水量更容易影响土壤通气性和土壤微生物活性,进而改变影响了土壤基质的养分循环,从而对无机氮转化产生影响.通常认为无机氮对温度的响应程度大于含水量[62],但是BM样地在整个秋季冻融时期,温度和含水量都处于较低水平且变化幅度较小,与温度相比含水量对土壤微生物活性起有效作用,这与赵媛[34]在研究秋末冻融循环对长白山地区无机氮的影响结果相似,即无机氮对含水量的响应程度大于温度.在秋季冻融前、中和后期,3种多年冻土泥炭地浅层(0~20 cm)土壤的环境因子与无机氮释放有关,其中浅层土壤温度对无机氮释放的贡献率最大(R2=0.685),这主要受当地非生长季寒冷气候的影响,温度通过改变土壤中生物酶和氨氧化细菌、硝化细菌的活性来影响氮的分解和无机氮的释放[63].但是不同土壤环境条件下微生物对温度敏感程度有很大差异[64],本研究发现,受非生长季温度逐渐降低的影响,XA仅在秋季冻融前期土壤温度对无机氮释放有贡献(R2=0.685),在秋季冻融后期不存在相关关系,这与表5中草地生态系统下青藏高原高寒草甸区研究发现短期的冻融交替可以显著增加土壤中的氮矿化累积结果相似.说明与长期冻融相比,短期温度变化会破坏土壤团聚结构从而影响氨氧化细菌的活性,一部分微生物为了维持繁殖不断汲取死亡微生物的养分,从而促进了微生物的活性,影响土壤氮矿化速度.但也有研究发现,短期冻融对土壤氮矿化并没有影响[57],土壤微生物会对温度变化做出调节反应,从而无机氮变化不显著.秋季冻融期无机氮对环境因子的响应程度存在以上差异的原因可能有以下四方面:第一,样地沿一定坡度选取,地势高低不同造成了土壤含水量存在显著差异.第二,受到地表植被的影响,土壤微生物的类别和数量存在差异因此对氮矿化程度不同[44].第三,与凋落物有关,不同凋落物分解的有机质不同会影响土壤有效基质[65].第四,3种泥炭地土壤的容重、孔隙度和pH值等物理性质存在差异[66].这些要素的综合影响造成了不同泥炭地间,以及冻融期不同时期间无机氮含量对温度和含水量响应程度的不同. ...
青藏高原多年冻土区活动层水热特性研究进展
1
2020
... 经逐步回归分析表明(表2~4),在不同冻融阶段,无机氮对环境因子的响应程度也存在差异.在整个秋季冻融期,BM浅层(10~20 cm)含水量对无机氮含量的影响程度最大(R2=0.301),这是由于浅层土壤含水量更容易影响土壤通气性和土壤微生物活性,进而改变影响了土壤基质的养分循环,从而对无机氮转化产生影响.通常认为无机氮对温度的响应程度大于含水量[62],但是BM样地在整个秋季冻融时期,温度和含水量都处于较低水平且变化幅度较小,与温度相比含水量对土壤微生物活性起有效作用,这与赵媛[34]在研究秋末冻融循环对长白山地区无机氮的影响结果相似,即无机氮对含水量的响应程度大于温度.在秋季冻融前、中和后期,3种多年冻土泥炭地浅层(0~20 cm)土壤的环境因子与无机氮释放有关,其中浅层土壤温度对无机氮释放的贡献率最大(R2=0.685),这主要受当地非生长季寒冷气候的影响,温度通过改变土壤中生物酶和氨氧化细菌、硝化细菌的活性来影响氮的分解和无机氮的释放[63].但是不同土壤环境条件下微生物对温度敏感程度有很大差异[64],本研究发现,受非生长季温度逐渐降低的影响,XA仅在秋季冻融前期土壤温度对无机氮释放有贡献(R2=0.685),在秋季冻融后期不存在相关关系,这与表5中草地生态系统下青藏高原高寒草甸区研究发现短期的冻融交替可以显著增加土壤中的氮矿化累积结果相似.说明与长期冻融相比,短期温度变化会破坏土壤团聚结构从而影响氨氧化细菌的活性,一部分微生物为了维持繁殖不断汲取死亡微生物的养分,从而促进了微生物的活性,影响土壤氮矿化速度.但也有研究发现,短期冻融对土壤氮矿化并没有影响[57],土壤微生物会对温度变化做出调节反应,从而无机氮变化不显著.秋季冻融期无机氮对环境因子的响应程度存在以上差异的原因可能有以下四方面:第一,样地沿一定坡度选取,地势高低不同造成了土壤含水量存在显著差异.第二,受到地表植被的影响,土壤微生物的类别和数量存在差异因此对氮矿化程度不同[44].第三,与凋落物有关,不同凋落物分解的有机质不同会影响土壤有效基质[65].第四,3种泥炭地土壤的容重、孔隙度和pH值等物理性质存在差异[66].这些要素的综合影响造成了不同泥炭地间,以及冻融期不同时期间无机氮含量对温度和含水量响应程度的不同. ...
Interaction and spatial distribution of wetland nitrogen processes
1
1997
... 经逐步回归分析表明(表2~4),在不同冻融阶段,无机氮对环境因子的响应程度也存在差异.在整个秋季冻融期,BM浅层(10~20 cm)含水量对无机氮含量的影响程度最大(R2=0.301),这是由于浅层土壤含水量更容易影响土壤通气性和土壤微生物活性,进而改变影响了土壤基质的养分循环,从而对无机氮转化产生影响.通常认为无机氮对温度的响应程度大于含水量[62],但是BM样地在整个秋季冻融时期,温度和含水量都处于较低水平且变化幅度较小,与温度相比含水量对土壤微生物活性起有效作用,这与赵媛[34]在研究秋末冻融循环对长白山地区无机氮的影响结果相似,即无机氮对含水量的响应程度大于温度.在秋季冻融前、中和后期,3种多年冻土泥炭地浅层(0~20 cm)土壤的环境因子与无机氮释放有关,其中浅层土壤温度对无机氮释放的贡献率最大(R2=0.685),这主要受当地非生长季寒冷气候的影响,温度通过改变土壤中生物酶和氨氧化细菌、硝化细菌的活性来影响氮的分解和无机氮的释放[63].但是不同土壤环境条件下微生物对温度敏感程度有很大差异[64],本研究发现,受非生长季温度逐渐降低的影响,XA仅在秋季冻融前期土壤温度对无机氮释放有贡献(R2=0.685),在秋季冻融后期不存在相关关系,这与表5中草地生态系统下青藏高原高寒草甸区研究发现短期的冻融交替可以显著增加土壤中的氮矿化累积结果相似.说明与长期冻融相比,短期温度变化会破坏土壤团聚结构从而影响氨氧化细菌的活性,一部分微生物为了维持繁殖不断汲取死亡微生物的养分,从而促进了微生物的活性,影响土壤氮矿化速度.但也有研究发现,短期冻融对土壤氮矿化并没有影响[57],土壤微生物会对温度变化做出调节反应,从而无机氮变化不显著.秋季冻融期无机氮对环境因子的响应程度存在以上差异的原因可能有以下四方面:第一,样地沿一定坡度选取,地势高低不同造成了土壤含水量存在显著差异.第二,受到地表植被的影响,土壤微生物的类别和数量存在差异因此对氮矿化程度不同[44].第三,与凋落物有关,不同凋落物分解的有机质不同会影响土壤有效基质[65].第四,3种泥炭地土壤的容重、孔隙度和pH值等物理性质存在差异[66].这些要素的综合影响造成了不同泥炭地间,以及冻融期不同时期间无机氮含量对温度和含水量响应程度的不同. ...
Simulating soil freeze/thaw cycles typical of winter alpine conditions: implications for N and P availability
1
2007
... 在整个秋季冻融期,频繁的冻融交替一方面会改变土壤理化性质影响氮元素的依附能力,释放出固定的NH4+-N[67],另一方面会使部分微生物死亡释放出细胞中的矿质氮,为生存微生物提供了养分,因此提高了非生长季土壤氮矿化速量.本研究中,XY浅层土壤NH4+-N在秋季冻融中期出现峰值之后下降,可能是因为在秋季冻融初期土壤环境因子(其中浅层土壤温度对NH4+-N贡献最显著为66.4%)变化使土壤粒子不断收缩造成土壤胶体中固定NH4+-N被释放而导致暂时升高,秋季冻融后期土壤理化性质遭到破坏使土壤孔隙度增大对NH4+-N的固持能力下降因而不断淋溶减少[68].XA样地在秋季冻融中期,一方面浅层环境因子对NO3--N贡献为59.4%且存在显著负相关关系(P<0.05)因而使浅层NO3--N含量明显上升,另一方面由于泥炭地土层黏重且渗透性差导致NO3--N淋溶性较差而上升[69]. ...
Research advances in soil nitrogen transformation as related to drying/wetting cycles
1
2010
... 在整个秋季冻融期,频繁的冻融交替一方面会改变土壤理化性质影响氮元素的依附能力,释放出固定的NH4+-N[67],另一方面会使部分微生物死亡释放出细胞中的矿质氮,为生存微生物提供了养分,因此提高了非生长季土壤氮矿化速量.本研究中,XY浅层土壤NH4+-N在秋季冻融中期出现峰值之后下降,可能是因为在秋季冻融初期土壤环境因子(其中浅层土壤温度对NH4+-N贡献最显著为66.4%)变化使土壤粒子不断收缩造成土壤胶体中固定NH4+-N被释放而导致暂时升高,秋季冻融后期土壤理化性质遭到破坏使土壤孔隙度增大对NH4+-N的固持能力下降因而不断淋溶减少[68].XA样地在秋季冻融中期,一方面浅层环境因子对NO3--N贡献为59.4%且存在显著负相关关系(P<0.05)因而使浅层NO3--N含量明显上升,另一方面由于泥炭地土层黏重且渗透性差导致NO3--N淋溶性较差而上升[69]. ...
干湿交替条件下土壤氮素转化及其影响研究进展
1
2010
... 在整个秋季冻融期,频繁的冻融交替一方面会改变土壤理化性质影响氮元素的依附能力,释放出固定的NH4+-N[67],另一方面会使部分微生物死亡释放出细胞中的矿质氮,为生存微生物提供了养分,因此提高了非生长季土壤氮矿化速量.本研究中,XY浅层土壤NH4+-N在秋季冻融中期出现峰值之后下降,可能是因为在秋季冻融初期土壤环境因子(其中浅层土壤温度对NH4+-N贡献最显著为66.4%)变化使土壤粒子不断收缩造成土壤胶体中固定NH4+-N被释放而导致暂时升高,秋季冻融后期土壤理化性质遭到破坏使土壤孔隙度增大对NH4+-N的固持能力下降因而不断淋溶减少[68].XA样地在秋季冻融中期,一方面浅层环境因子对NO3--N贡献为59.4%且存在显著负相关关系(P<0.05)因而使浅层NO3--N含量明显上升,另一方面由于泥炭地土层黏重且渗透性差导致NO3--N淋溶性较差而上升[69]. ...
Discussion on the dependence of wetland and frozen soil in Daxinganling area
1
2007
... 在整个秋季冻融期,频繁的冻融交替一方面会改变土壤理化性质影响氮元素的依附能力,释放出固定的NH4+-N[67],另一方面会使部分微生物死亡释放出细胞中的矿质氮,为生存微生物提供了养分,因此提高了非生长季土壤氮矿化速量.本研究中,XY浅层土壤NH4+-N在秋季冻融中期出现峰值之后下降,可能是因为在秋季冻融初期土壤环境因子(其中浅层土壤温度对NH4+-N贡献最显著为66.4%)变化使土壤粒子不断收缩造成土壤胶体中固定NH4+-N被释放而导致暂时升高,秋季冻融后期土壤理化性质遭到破坏使土壤孔隙度增大对NH4+-N的固持能力下降因而不断淋溶减少[68].XA样地在秋季冻融中期,一方面浅层环境因子对NO3--N贡献为59.4%且存在显著负相关关系(P<0.05)因而使浅层NO3--N含量明显上升,另一方面由于泥炭地土层黏重且渗透性差导致NO3--N淋溶性较差而上升[69]. ...
试论大兴安岭地区湿地与冻土的依存关系
1
2007
... 在整个秋季冻融期,频繁的冻融交替一方面会改变土壤理化性质影响氮元素的依附能力,释放出固定的NH4+-N[67],另一方面会使部分微生物死亡释放出细胞中的矿质氮,为生存微生物提供了养分,因此提高了非生长季土壤氮矿化速量.本研究中,XY浅层土壤NH4+-N在秋季冻融中期出现峰值之后下降,可能是因为在秋季冻融初期土壤环境因子(其中浅层土壤温度对NH4+-N贡献最显著为66.4%)变化使土壤粒子不断收缩造成土壤胶体中固定NH4+-N被释放而导致暂时升高,秋季冻融后期土壤理化性质遭到破坏使土壤孔隙度增大对NH4+-N的固持能力下降因而不断淋溶减少[68].XA样地在秋季冻融中期,一方面浅层环境因子对NO3--N贡献为59.4%且存在显著负相关关系(P<0.05)因而使浅层NO3--N含量明显上升,另一方面由于泥炭地土层黏重且渗透性差导致NO3--N淋溶性较差而上升[69]. ...


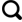


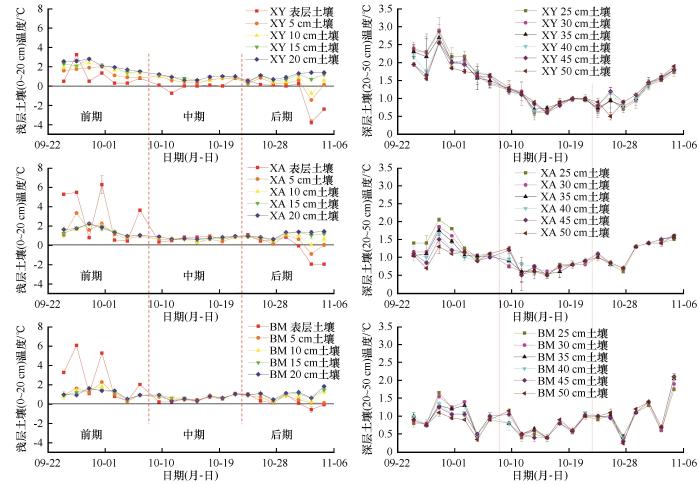
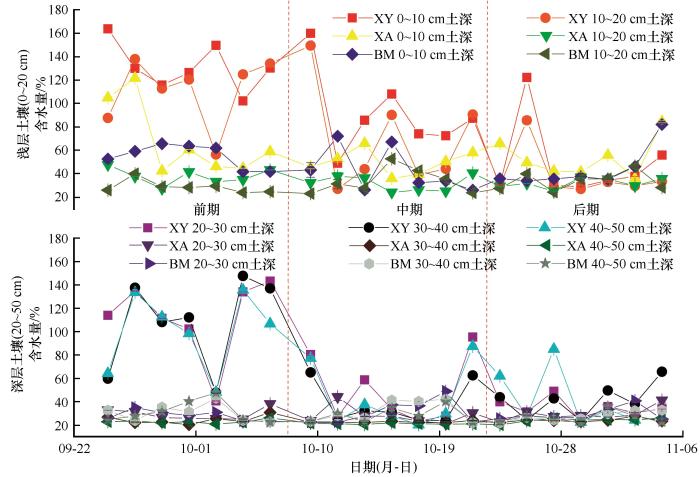
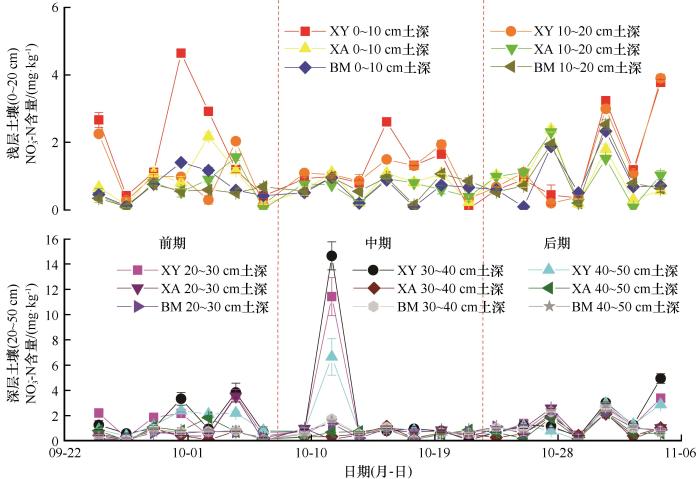

 甘公网安备 62010202000676号
甘公网安备 62010202000676号
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}